Use of Dry Dairy Manure Pellets as Nutrient Source for Tomato (Solanum lycopersicum var. cerasiforme) Growth in Soilless Media

Abstract
:1. Introduction
2. Materials and Methods
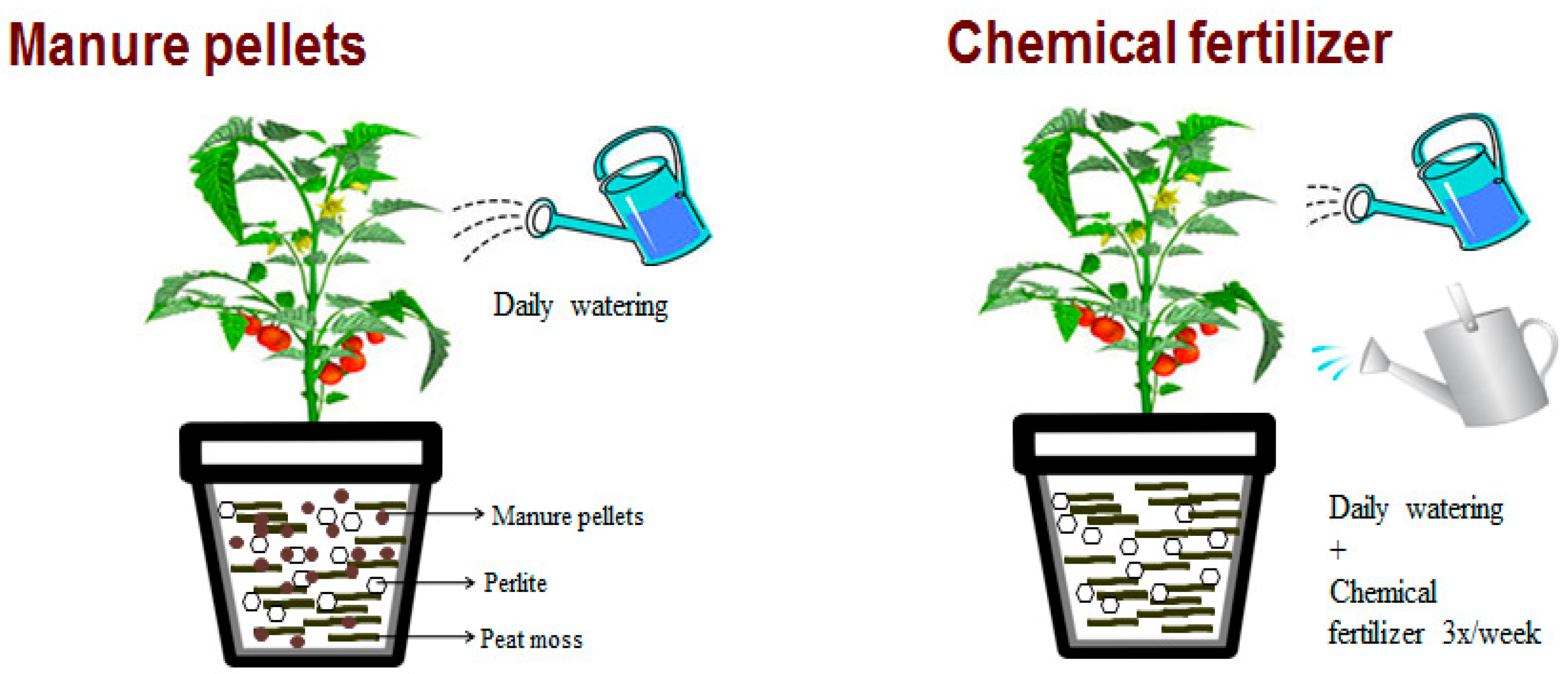
2.1. Manure Pellets
2.2. Soilless Media Treatments
2.3. Tomato Plant Culture
2.4. Statistical Analysis
3. Results and Discussion
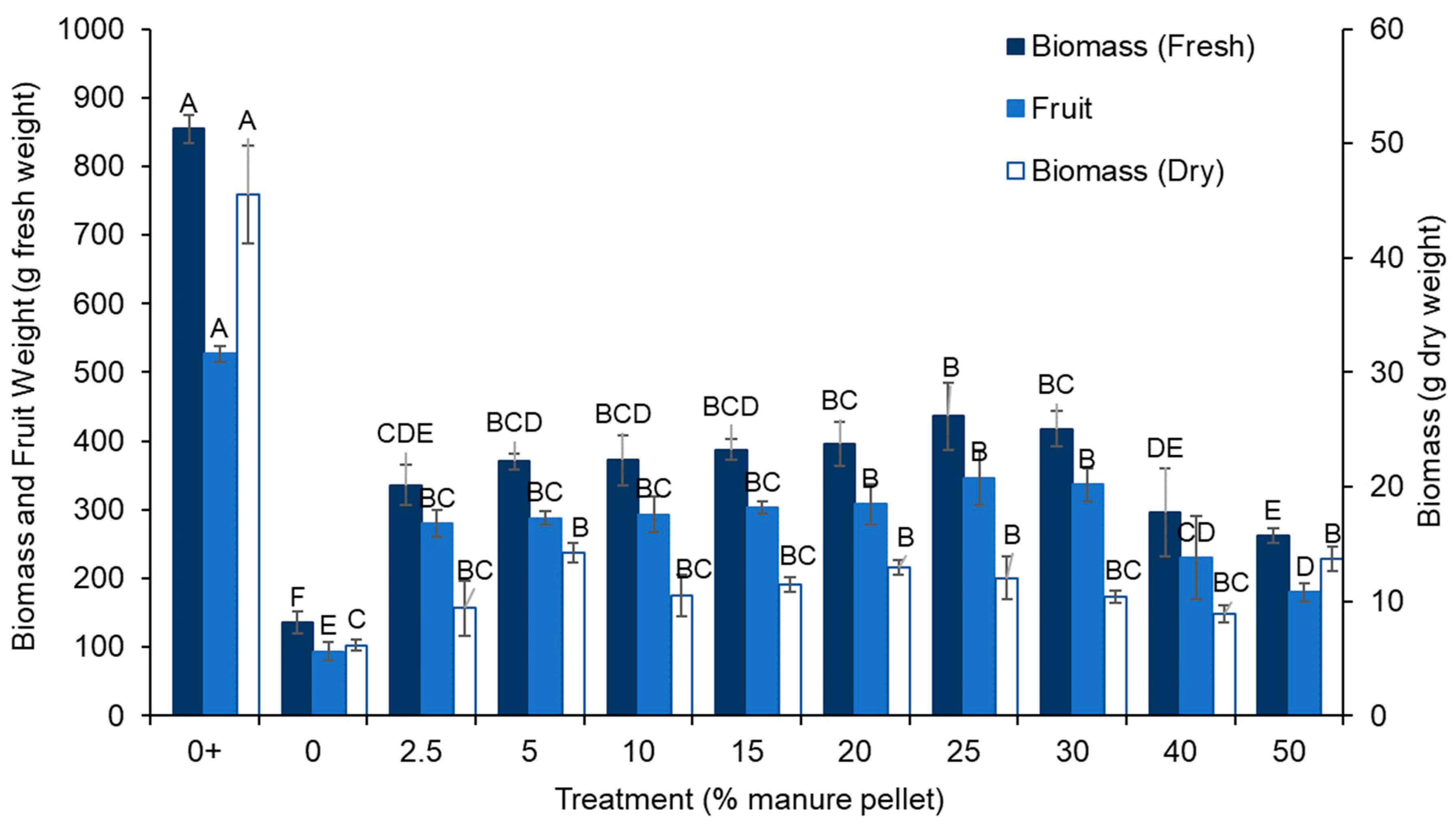
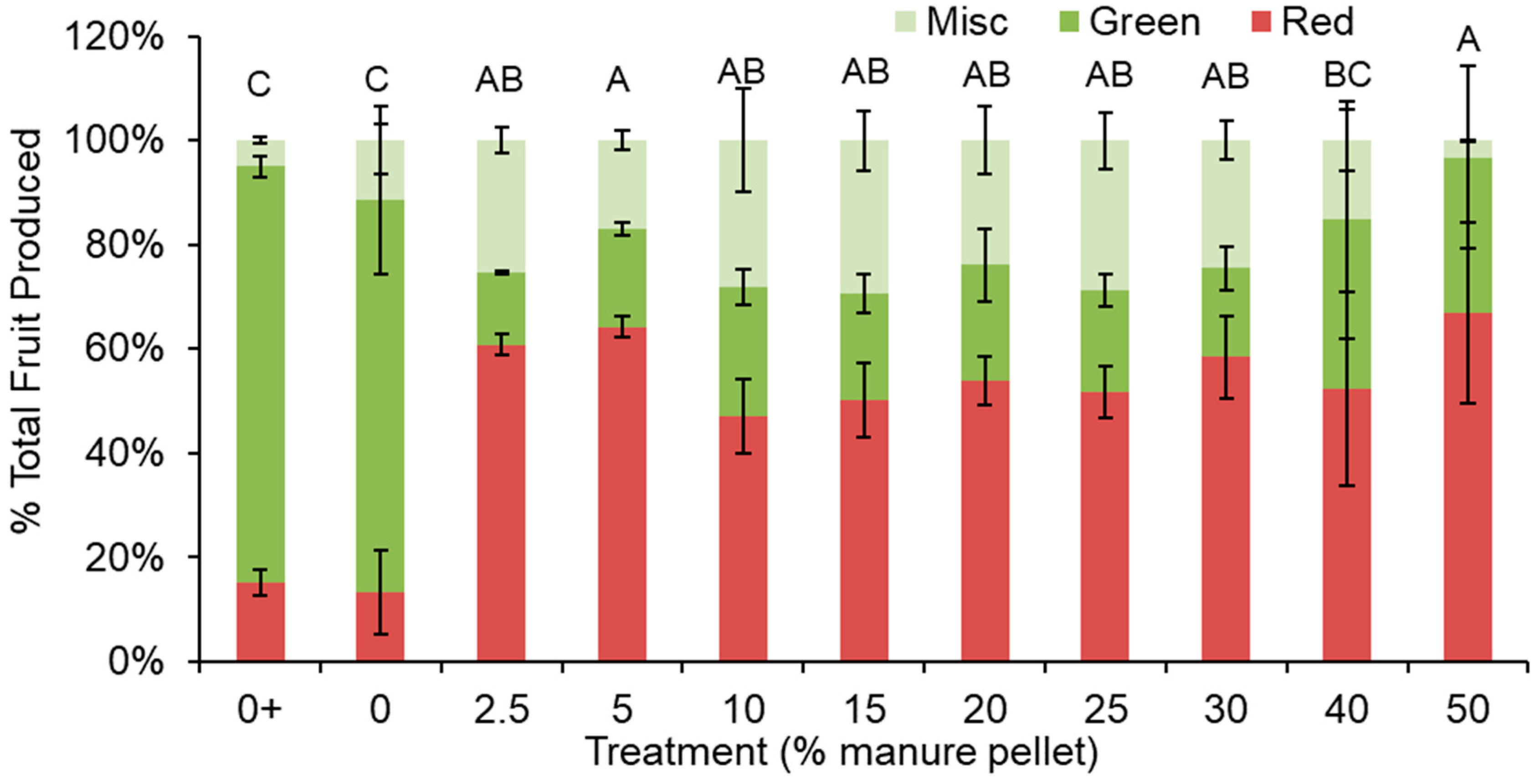
3.1. Fruit
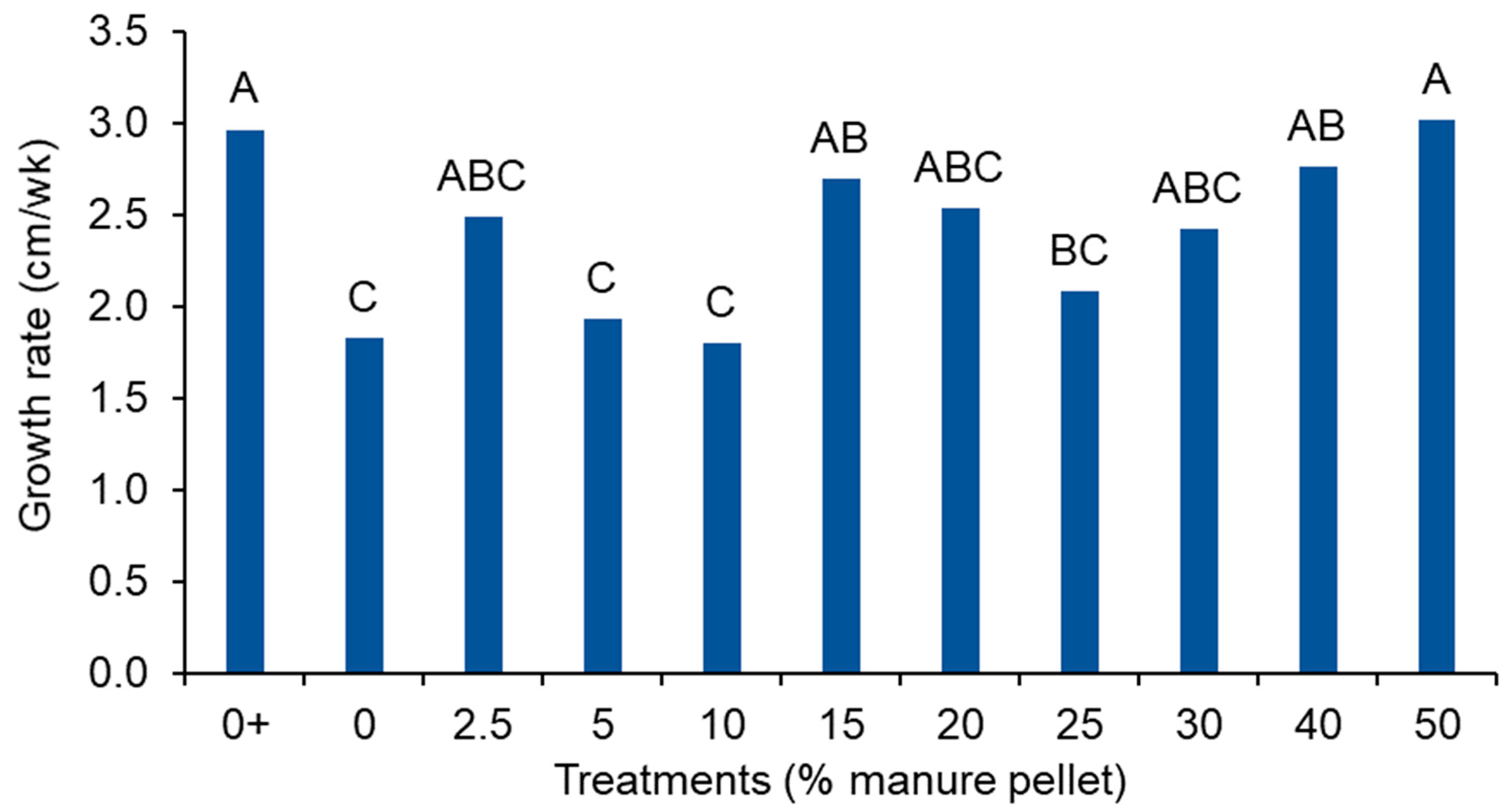
3.2. Plant Growth
3.3. Biomass
3.4. Soilless Media Mix Analysis Post-Harvest
4. Conclusions and Implications
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Pachepsky, Y.A.; Sadeghi, A.; Bradford, S.; Shelton, D.; Guber, A.; Dao, T. Transport and fate of manure-borne pathogens: Modeling perspective. Agric. Water Manag. 2006, 86, 81–92. [Google Scholar] [CrossRef]
- Sharpley, A.N.; McDowell, R.W.; Kleinman, P.J. Amounts, forms, and solubility of phosphorus in soils receiving manure. Soil Sci. Soc. Am. J. 2004, 68, 2048–2057. [Google Scholar] [CrossRef]
- Szögi, A.A.; Vanotti, M.B.; Hunt, P.G. Dewatering of phosphorus extracted from liquid swine waste. Bioresour. Technol. 2006, 97, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Møller, H.B.; Sommer, S.G.; Ahring, B.K. Separation efficiency and particle size distribution in relation to manure type and storage conditions. Bioresour. Technol. 2002, 85, 189–196. [Google Scholar] [CrossRef]
- Weiss, W.; Willett, L.; St-Pierre, N.; Borger, D.; McKelvey, T.; Wyatt, D. Varying forage type, metabolizable protein concentration, and carbohydrate source affects manure excretion, manure ammonia, and nitrogen metabolism of dairy cows. J. Dairy Sci. 2009, 92, 5607–5619. [Google Scholar] [CrossRef] [PubMed]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.B.; Combs, S.; Hoskins, B.; Jarman, J.; Kovar, J.; Watson, M.; Wolf, A.; Wolf, N. Recommended Methods of Manure Analysis; University of Wisconsin Cooperative Extension Publishing: Madison, WI, USA, 2003. [Google Scholar]
- Liu, Z.; Carroll, Z.S.; Long, S.C.; Gunasekaran, S.; Runge, T. Use of cationic polymers to reduce pathogen levels during dairy manure separation. J. Environ. Manag. 2016, 166, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Warncke, D. Greenhouse root media. In Recommended Chemical Soil Test Procedures for the North Central Region, 1st ed.; NCR Publication: Kansas City, MO, USA, 1998; Chapter 14; Volume 221, pp. 61–64. [Google Scholar]
- Butler, T.; Muir, J.P. Dairy manure compost improves soil and increases tall wheatgrass yield. Agron. J. 2006, 98, 1090–1096. [Google Scholar] [CrossRef]
- NCDA&CS. Tissue Sampling & Analysis for Greenhouse Tomatoes; North Carolina Department of Agriculture and Consumer Services Agronomic Division: Raleigh, NC, USA, 2011. Available online: http://www.ncagr.gov/agronomi/pdffiles/gtomato.pdf (accessed on 17 October 2018).
- Bot, J.L.; Jeannequin, B.; Fabre, R. Growth and nitrogen status of soilless tomato plants following nitrate withdrawal from the nutrient solution. Ann. Bot. 2001, 88, 361–370. [Google Scholar] [CrossRef]
- Juárez-Maldonado, A.; de-Alba-Romenus, K.; Morales-Díaz, A.B.; Benavides-Mendoza, A. Macro-nutrient uptake dynamics in greenhouse tomato crop. J. Plant Nutr. 2017, 40, 1908–1919. [Google Scholar] [CrossRef]
- Liu, C.; Muchhal, U.S.; Uthappa, M.; Kononowicz, A.K.; Raghothama, K.G. Tomato phosphate transporter genes are differentially regulated in plant tissues by phosphorus. Plant Physiol. 1998, 116, 91–99. [Google Scholar] [CrossRef] [PubMed]
- White, P.J. Ion uptake mechanisms of individual cells and roots: Short-distance transport. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012; Chapter 2; pp. 7–47. [Google Scholar]
- Sainju, U.M.; Dris, R.; Singh, B. Mineral nutrition of tomato. J. Food. Agric. Environ. 2003, 1, 176–183. [Google Scholar]
- Belda, R.M.; Mendoza-Hernández, D.; Fornes, F. Nutrient-rich compost versus nutrient-poor vermicompost as growth media for ornamental-plant production. J. Plant Nutr. Soil Sci. 2013, 176, 827–835. [Google Scholar] [CrossRef]
- Elia, A.; Conversa, G.; La Rotonda, P.; Montemurrow, F. Nitrogen level effect on yield and quality of fertigated processing tomato in Southern Italy. Acta Hortic. 2007, 235–240. [Google Scholar] [CrossRef]
- Ozores-Hampton, M.; Simonne, E.; Roka, F.; Morgan, K.; Sargent, S.; Snodgrass, C.; McAvoy, E. Nitrogen rates effects on the yield nutritional status, fruit quality, and the profitability of tomato grown in the spring with subsurface irrigation. Hortscience 2012, 57, 1129–1135. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | N | P | K | Ca | Mg | S | Zn | B | Mn | Fe | Cu | Al | Na |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | mg·kg−1 | ||||||||||||
| Pellets | 3.7 | 1.8 | 0.90 | 2.9 | 1.3 | 0.50 | 529 | 27 | 290 | 1000 | 170 | 634 | 3000 |
| Treatment | ||||||||
|---|---|---|---|---|---|---|---|---|
| % by Volume | g Pellets pot−1 | Mg Pellets ha−1 | g N pot−1 | kg N ha−1 | g P pot−1 | kg P ha−1 | g K pot−1 | kg K ha−1 |
| 2.5% | 21 | 0.21 | 0.8 | 777 | 0.4 | 378 | 0.2 | 189 |
| 5% | 42 | 0.42 | 1.6 | 1554 | 0.8 | 756 | 0.4 | 378 |
| 10% | 84 | 0.84 | 3.1 | 3108 | 1.5 | 1512 | 0.8 | 756 |
| 15% | 126 | 1.26 | 4.7 | 4662 | 2.3 | 2268 | 1.1 | 1134 |
| 20% | 168 | 1.68 | 6.2 | 6216 | 3.0 | 3024 | 1.5 | 1512 |
| 25% | 210 | 2.10 | 7.8 | 7770 | 3.8 | 3780 | 1.9 | 1890 |
| 30% | 252 | 2.52 | 9.3 | 9324 | 4.5 | 4536 | 2.3 | 2268 |
| 40% | 336 | 3.36 | 12.4 | 12432 | 6.0 | 6048 | 3.0 | 3024 |
| 50% | 420 | 4.20 | 15.5 | 15540 | 7.6 | 7560 | 3.8 | 3780 |
| Treatment | N | P | K | Ca | Mg | S | Zn | Mn | B | Fe | Cu |
|---|---|---|---|---|---|---|---|---|---|---|---|
| % Manure | % | mg·kg−1 | |||||||||
| Sufficiency range * | 3.5–5.0 | 0.3-0.65 | 2.8-4.0 | 1.0–3.0 | 0.35–1.0 | 0.2–1.0 | 18–80 | 25–200 | 30–75 | 50–300 | 5–35 |
| 0+ | 3.90 | 0.61 | 3.15 | 2.19 | 1.04 | 0.36 | 37 | 166 | 141 | 94 | 8.8 |
| 0 | 1.15 | 0.18 | 1.02 | 1.76 | 1.14 | 0.60 | 3.8 | 71 | 41 | 46 | 2.5 |
| 2.5 | 1.30 | 0.12 | 0.64 | 1.89 | 1.27 | 0.65 | 3.1 | 101 | 37 | 46 | 3.4 |
| 5 | 1.27 | 0.15 | 0.46 | 2.11 | 1.46 | 0.64 | 4.0 | 69 | 41 | 42 | 4.4 |
| 10 | 1.49 | 0.35 | 0.38 | 2.88 | 1.80 | 0.73 | 5.6 | 38 | 53 | 45 | 5.0 |
| 15 | 1.61 | 0.58 | 0.40 | 2.98 | 2.07 | 0.70 | 6.3 | 36 | 84 | 36 | 6.6 |
| 20 | 1.63 | 0.50 | 0.35 | 2.63 | 2.09 | 0.69 | 5.0 | 38 | 86 | 39 | 6.9 |
| 25 | 1.62 | 0.63 | 0.38 | 3.05 | 2.60 | 0.76 | 6.2 | 49 | 108 | 35 | 7.0 |
| 30 | 1.62 | 0.73 | 0.38 | 3.43 | 2.69 | 0.83 | 9.0 | 65 | 102 | 31 | 7.7 |
| 40 | 1.41 | 0.54 | 0.43 | 2.58 | 2.58 | 0.74 | 8.6 | 55 | 95 | 28 | 7.7 |
| 50 | 1.91 | 0.47 | 0.76 | 1.59 | 2.40 | 0.58 | 15 | 47 | 77 | 35 | 7.9 |
| Treatment | pH | EC | Nitrate | P | K | Ca | Mg | Fe | Na | Zn | S |
|---|---|---|---|---|---|---|---|---|---|---|---|
| dS m−1 | mg·L−1 DTPA Saturated Media Extract Method | ||||||||||
| 0%+ | 5.7 | 0.14 | 13 | 20 | 20 | 129 | 88 | 33 | 27 | 44 | 5 |
| 0% | 6.1 | 0.09 | 0 | 5 | 6 | 142 | 90 | 38 | 11 | 9 | 8 |
| 2.5% | 7.3 | 0.10 | 0 | 3 | 2 | 168 | 82 | 29 | 11 | 63 | 5 |
| 5% | 7.6 | 0.12 | 1 | 4 | 5 | 190 | 82 | 23 | 11 | 92 | 5 |
| 10% | 7.9 | 0.15 | 6 | 21 | 4 | 220 | 86 | 16 | 10 | 163 | 9 |
| 15% | 7.8 | 0.21 | 5 | 49 | 5 | 226 | 101 | 12 | 11 | 236 | 9 |
| 20% | 7.9 | 0.26 | 4 | 53 | 5 | 223 | 111 | 9 | 12 | 259 | 11 |
| 25% | 7.9 | 0.25 | 5 | 46 | 27 | 233 | 131 | 6 | 19 | 252 | 16 |
| 30% | 7.9 | 0.27 | 12 | 43 | 9 | 217 | 121 | 7 | 12 | 311 | 17 |
| 40% | 8.0 | 0.36 | 8 | 35 | 14 | 206 | 140 | 5 | 15 | 337 | 22 |
| 50% | 8.2 | 0.42 | 22 | 26 | 14 | 222 | 185 | 5 | 41 | 401 | 39 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Howe, J.; Wang, X.; Liang, X.; Runge, T. Use of Dry Dairy Manure Pellets as Nutrient Source for Tomato (Solanum lycopersicum var. cerasiforme) Growth in Soilless Media. Sustainability 2019, 11, 811. https://doi.org/10.3390/su11030811
Liu Z, Howe J, Wang X, Liang X, Runge T. Use of Dry Dairy Manure Pellets as Nutrient Source for Tomato (Solanum lycopersicum var. cerasiforme) Growth in Soilless Media. Sustainability. 2019; 11(3):811. https://doi.org/10.3390/su11030811
Chicago/Turabian StyleLiu, Zong, Julie Howe, Xiao Wang, Xiao Liang, and Troy Runge. 2019. "Use of Dry Dairy Manure Pellets as Nutrient Source for Tomato (Solanum lycopersicum var. cerasiforme) Growth in Soilless Media" Sustainability 11, no. 3: 811. https://doi.org/10.3390/su11030811
APA StyleLiu, Z., Howe, J., Wang, X., Liang, X., & Runge, T. (2019). Use of Dry Dairy Manure Pellets as Nutrient Source for Tomato (Solanum lycopersicum var. cerasiforme) Growth in Soilless Media. Sustainability, 11(3), 811. https://doi.org/10.3390/su11030811