Evaluation of Biological Reference Points for Conservation and Management of the Bigeye Thresher Shark, Alopias superciliosus, in the Northwest Pacific


Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Data
2.2. Mortality Estimation
2.3. Model Fitting and Convergence
2.4. Biological Reference Points
2.5. Demographic Model Development
- (1)
- Age at maturity = 12 years
- (2)
- Fecundity = 2 pups
- (3)
- Sex ratio = 0.5 for embryos
- (4)
- Selectivity (assumed constant dome-shaped distribution).
- (5)
- A knife-edge maturity was assumed in this model and age-at-first-reproduction calculated as the mean age at maturity + the gestation period (set as 1 year in this study).
2.6. Design of the Simulation Study
2.6.1. Biological Reference Points
2.6.2. Estimates of Population Growth Rates
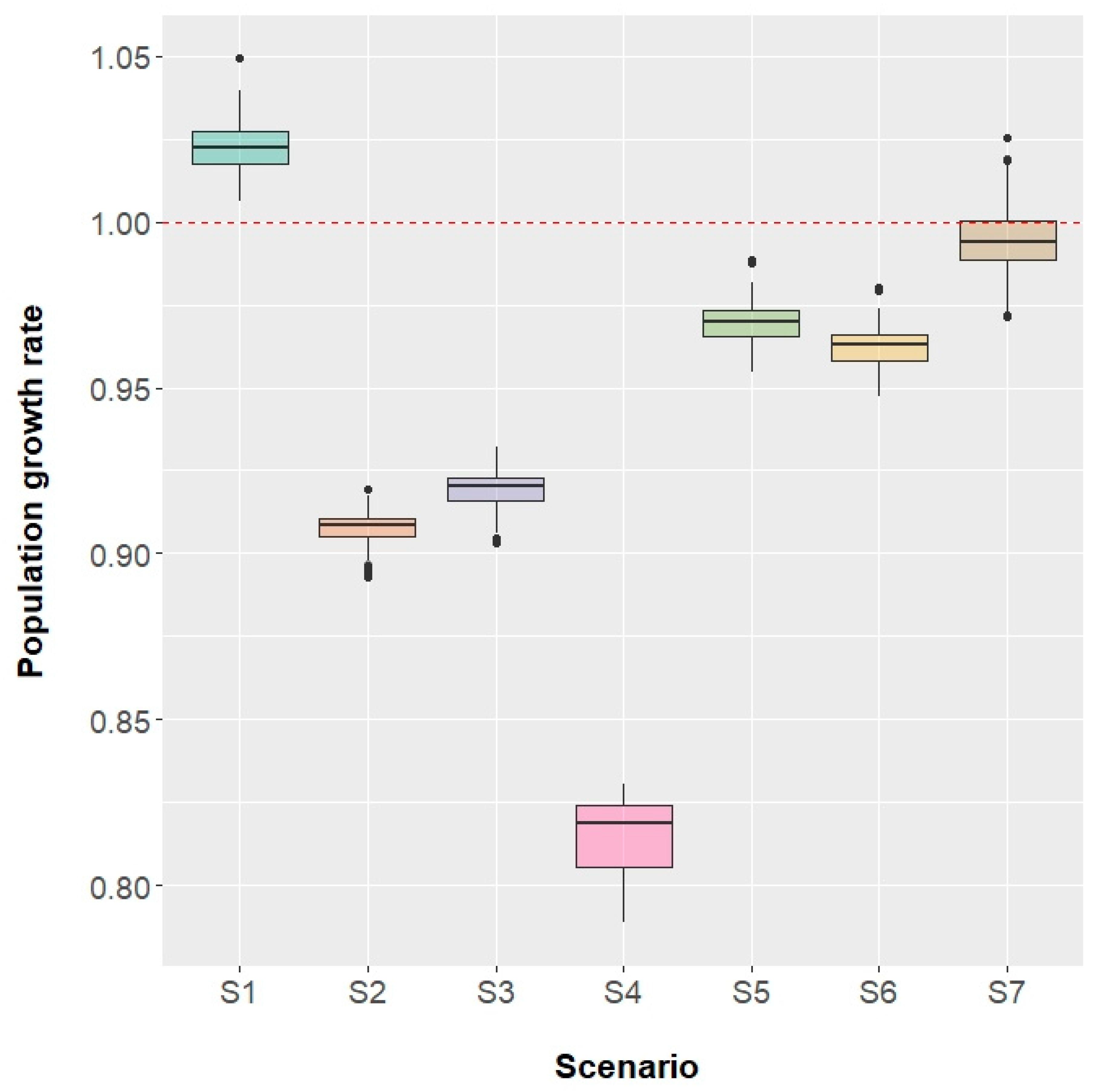
- Scenario 1: fishing mortality for all ages set to 0.
- Scenario 2: fishing mortality equal to its current level by age.
- Scenario 3: fishing mortality set to the F0.1 level.
- Scenario 4: fishing mortality set to the Fmax level.
- Scenario 5: fishing mortality set to the FSPR35% level.
- Scenario 6: fishing mortality set to the FSPR30% level.
- Scenario 7: fishing mortality set to the Fcrit level.
3. Results
3.1. Deterministic Estimates
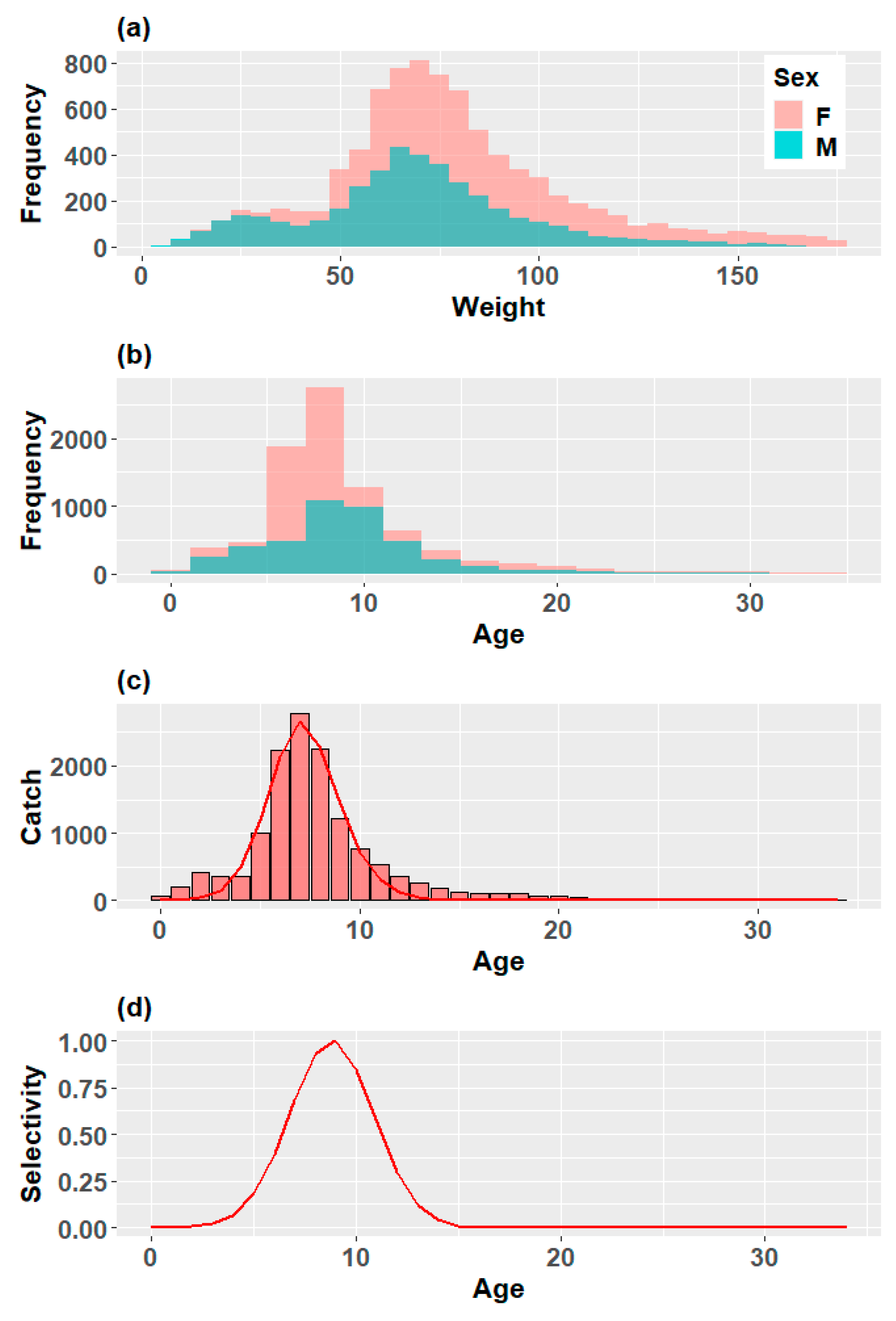
3.1.1. Sex-Specific Catch and Weight Compositions
3.1.2. Mortality and Selectivity
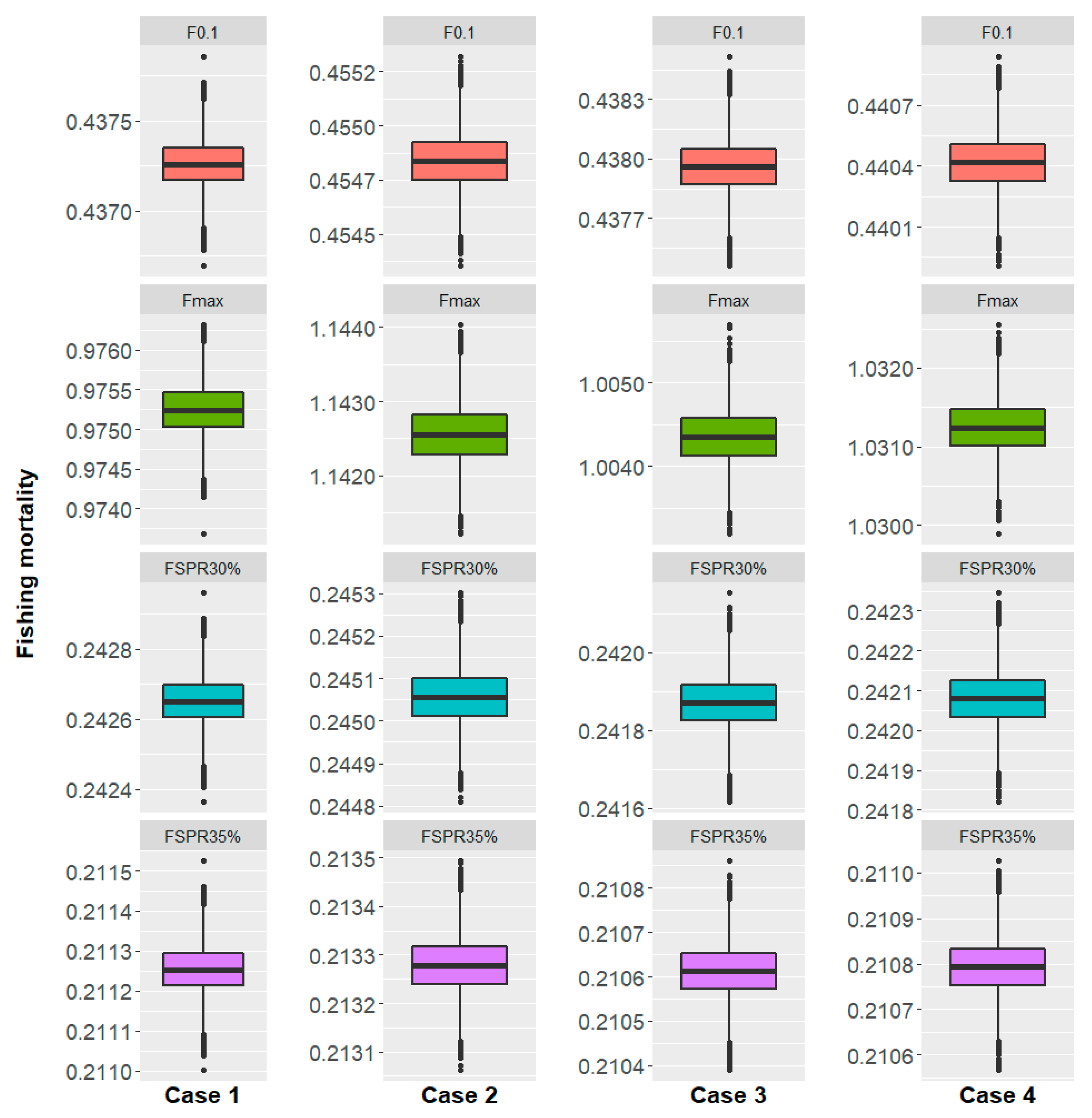
3.1.3. Biological Reference Points
3.1.4. Population Increase Rate
3.2. Estimates with Uncertainty
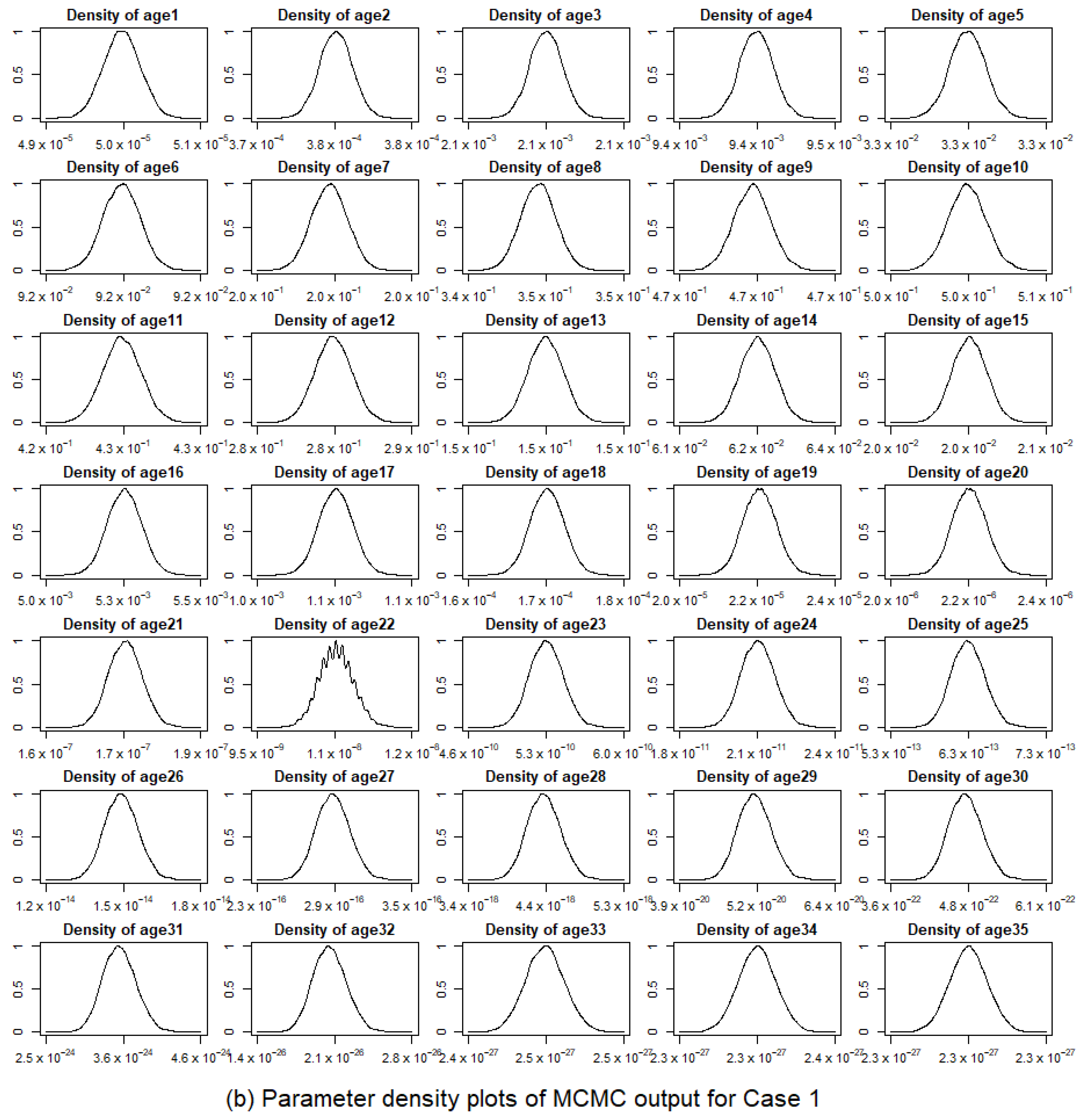
3.2.1. Model Convergence
3.2.2. Biological Reference Points
3.2.3. Population Increase Rate
4. Discussion
4.1. Biological Reference Points
4.2. Demographic Model
4.3. Uncertainty
4.4. Stock Status
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fishing Mortality | ||||||||
|---|---|---|---|---|---|---|---|---|
| Age/Case | Case 1 | Case 2 | Case 3 | Case 4 | ||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| 1 | 0.0001 | 0.0000 | 0.0000 | 0.0000 | 0.0001 | 0.0000 | 0.0000 | 0.0000 |
| 2 | 0.0004 | 0.0000 | 0.0004 | 0.0000 | 0.0004 | 0.0000 | 0.0004 | 0.0000 |
| 3 | 0.0021 | 0.0000 | 0.0020 | 0.0000 | 0.0021 | 0.0000 | 0.0021 | 0.0000 |
| 4 | 0.0094 | 0.0000 | 0.0091 | 0.0000 | 0.0093 | 0.0000 | 0.0092 | 0.0000 |
| 5 | 0.0331 | 0.0000 | 0.0322 | 0.0000 | 0.0329 | 0.0000 | 0.0327 | 0.0000 |
| 6 | 0.0918 | 0.0000 | 0.0896 | 0.0000 | 0.0915 | 0.0000 | 0.0911 | 0.0000 |
| 7 | 0.2005 | 0.0001 | 0.1967 | 0.0001 | 0.2006 | 0.0001 | 0.2001 | 0.0001 |
| 8 | 0.3457 | 0.0002 | 0.3399 | 0.0002 | 0.3468 | 0.0002 | 0.3461 | 0.0002 |
| 9 | 0.4700 | 0.0004 | 0.4628 | 0.0004 | 0.4723 | 0.0004 | 0.4716 | 0.0004 |
| 10 | 0.5042 | 0.0007 | 0.4965 | 0.0007 | 0.5071 | 0.0007 | 0.5063 | 0.0007 |
| 11 | 0.4266 | 0.0009 | 0.4197 | 0.0009 | 0.4291 | 0.0009 | 0.4282 | 0.0009 |
| 12 | 0.2848 | 0.0009 | 0.2795 | 0.0009 | 0.2861 | 0.0009 | 0.2853 | 0.0009 |
| 13 | 0.1499 | 0.0007 | 0.1467 | 0.0006 | 0.1504 | 0.0006 | 0.1497 | 0.0006 |
| 14 | 0.0623 | 0.0004 | 0.0606 | 0.0003 | 0.0623 | 0.0004 | 0.0619 | 0.0004 |
| 15 | 0.0204 | 0.0002 | 0.0198 | 0.0001 | 0.0203 | 0.0001 | 0.0202 | 0.0001 |
| 16 | 0.0053 | 0.0000 | 0.0051 | 0.0000 | 0.0052 | 0.0000 | 0.0052 | 0.0000 |
| 17 | 0.0011 | 0.0000 | 0.0010 | 0.0000 | 0.0011 | 0.0000 | 0.0010 | 0.0000 |
| 18 | 0.0002 | 0.0000 | 0.0002 | 0.0000 | 0.0002 | 0.0000 | 0.0002 | 0.0000 |
| 19 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 20 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 21 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 22 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 23 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 24 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 25 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 26 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 27 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 28 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 29 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 30 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 31 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 32 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 33 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 34 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 35 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| Selectivity | ||||||||
|---|---|---|---|---|---|---|---|---|
| Age/Case | Case 1 | Case 2 | Case 3 | Case 4 | ||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| 1 | 0.0001 | 0.0000 | 0.0001 | 0.0000 | 0.0001 | 0.0000 | 0.0001 | 0.0000 |
| 2 | 0.0007 | 0.0000 | 0.0007 | 0.0000 | 0.0007 | 0.0000 | 0.0007 | 0.0000 |
| 3 | 0.0042 | 0.0000 | 0.0041 | 0.0000 | 0.0041 | 0.0000 | 0.0041 | 0.0000 |
| 4 | 0.0187 | 0.0000 | 0.0184 | 0.0000 | 0.0184 | 0.0000 | 0.0183 | 0.0000 |
| 5 | 0.0657 | 0.0001 | 0.0649 | 0.0001 | 0.0648 | 0.0001 | 0.0646 | 0.0001 |
| 6 | 0.1820 | 0.0002 | 0.1806 | 0.0002 | 0.1804 | 0.0002 | 0.1800 | 0.0002 |
| 7 | 0.3977 | 0.0005 | 0.3961 | 0.0005 | 0.3957 | 0.0005 | 0.3952 | 0.0005 |
| 8 | 0.6856 | 0.0007 | 0.6845 | 0.0007 | 0.6838 | 0.0007 | 0.6836 | 0.0007 |
| 9 | 0.9323 | 0.0006 | 0.9321 | 0.0006 | 0.9315 | 0.0005 | 0.9315 | 0.0005 |
| 10 | 1.0000 | 0.0000 | 1.0000 | 0.0000 | 1.0000 | 0.0000 | 1.0000 | 0.0000 |
| 11 | 0.8462 | 0.0007 | 0.8453 | 0.0007 | 0.8461 | 0.0007 | 0.8457 | 0.0007 |
| 12 | 0.5648 | 0.0010 | 0.5630 | 0.0010 | 0.5642 | 0.0010 | 0.5635 | 0.0010 |
| 13 | 0.2974 | 0.0009 | 0.2954 | 0.0009 | 0.2965 | 0.0009 | 0.2958 | 0.0009 |
| 14 | 0.1235 | 0.0005 | 0.1221 | 0.0005 | 0.1228 | 0.0005 | 0.1223 | 0.0005 |
| 15 | 0.0405 | 0.0002 | 0.0398 | 0.0002 | 0.0401 | 0.0002 | 0.0398 | 0.0002 |
| 16 | 0.0105 | 0.0001 | 0.0102 | 0.0001 | 0.0103 | 0.0001 | 0.0102 | 0.0001 |
| 17 | 0.0021 | 0.0000 | 0.0021 | 0.0000 | 0.0021 | 0.0000 | 0.0021 | 0.0000 |
| 18 | 0.0003 | 0.0000 | 0.0003 | 0.0000 | 0.0003 | 0.0000 | 0.0003 | 0.0000 |
| 19 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 20 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 21 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 22 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 23 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 24 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 25 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 26 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 27 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 28 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 29 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 30 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 31 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 32 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 33 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 34 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| 35 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
References
- Holden, M.J. Are long-term sustainable fisheries for elasmobranchs possible? Rapp. Proces. Verbaux Reun. Counseil Int. Pour l’Exploracion Mer. 1973, 164, 330–367. [Google Scholar]
- Hoenig, J.M.; Gruber, S.H. Life-history patterns in the elasmobranchs: Implications or fisheries management. NOAA Tech. Rep. 1990, 90, 1–16. [Google Scholar]
- Cortés, E. Comparative Life History and Demography of Pelagic Sharks. In Sharks of the Open Ocean: Biology, Fisheries and Conservation; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2008; pp. 309–320. [Google Scholar]
- Dulvy, N.K.; Baum, J.K.; Clarke, S.; Compagno, L.J.V.; Cortés, E.; Domingo, A.; Fordham, S.; Fowler, S.; Francis, M.P.; Gibson, C.; et al. You can swim but you can’t hide: The global status and conservation of oceanic pelagic sharks and rays. Aquat. Conserv. Mar. Freshw. Ecol. 2008, 18, 459–482. [Google Scholar] [CrossRef]
- Musick, J.A.; Burgess, G.; Cailliet, G.; Camhi, M.; Fordham, S. Management of sharks and their relatives (Elasmobranchii). Fish 2000, 25, 9–13. [Google Scholar] [CrossRef]
- Stevens, J.D.; Bonfil, R.; Dulvy, N.K.; Walker, P.A. The effects of fishing on sharks, rays, and chimaeras (chondrichthyans), and implications for marine ecosystems. Ices J. Mar. Sci. 2000, 57, 476–494. [Google Scholar] [CrossRef]
- Bonfil, R. Overview of World Elasmobranch Fisheries; FAO Fishery Technology Paper 341; FAO: Rome, Italy, 1994. [Google Scholar]
- Clarke, S.C.; McAllister, M.K.; Milner-Gulland, E.J.; Kirkwood, G.P.; Michielsens, C.G.J.; Agnew, D.J.; Pikitch, E.K.; Nakano, H.; Shivji, M.S. Global estimates of shark catches using trade records from commercial markets. Ecol. Lett. 2006, 9, 1115–1126. [Google Scholar] [CrossRef]
- Fernandez-Carvalho, J.; Coelho, R.; Mejuto, J.; Cortés, E.; Domingo, A.; Yokawa, K.; Liu, K.M.; García-Cortés, B.; Forsellado, R.; Ohshimo, S.; et al. Pan-Atlantic distribution patterns and reproductive biology of the bigeye thresher, (Alopias superciliosus). Rev. Fish Biol. Fish. 2015, 25, 511–568. [Google Scholar] [CrossRef]
- Pikitch, E.K.; Camhi, M.D.; Babcock, E.A. Introduction to Sharks of the Open Ocean. In Sharks of the Open Ocean: Biology, Fisheries and Conservation; Camhi, M.D., Pikitch, E.K., Babcock, E.A., Eds.; Blackwell Scientific Publications: Oxford, UK, 2008; pp. 3–13. [Google Scholar]
- Simpfendorfer, C.A. Demographic models: Life tables, matrix models and rebound potential. FAO Fish. Technol. Pap. 2005, 474, 143–153. [Google Scholar]
- Smart, J.J.; Chin, A.; Tobin, A.J.; White, W.T.; Kumasi, B.; Simpfendorfer, C.A. Stochastic demographic analyses of the silvertip shark (Carcharhinus albimarginatus) and the common blacktip shark (Carcharhinus limbatus) from the Indo-Pacific. Fish. Res. 2017, 191, 95–107. [Google Scholar] [CrossRef]
- Hayes, D.B. A biological reference point based on the Leslie matrix. Fish. Bull. 2000, 98, 75–85. [Google Scholar]
- Gallucci, V.F.; Taylor, I.G.; Erzini, K. Conservation and management of exploited shark populations based on reproductive value. Can. J. Fish. Aquat. Sci. 2006, 63, 931–942. [Google Scholar] [CrossRef]
- Aires-da-Silva, A.M.; Gallucci, V.F. Demographic and risk analyses applied to management and conservation of the blue shark (Prionace glauca) in the North Atlantic Ocean. Mar. Freshwater Res. 2007, 58, 570–580. [Google Scholar] [CrossRef]
- Tsai, W.P.; Liu, K.M.; Punt, A.E.; Sun, C.L. Assessing the potential biases of ignoring sexual dimorphism and mating mechanism in using a single-sex demographic model: The shortfin mako shark as a case study. Ices J. Mar. Sci. 2015, 72, 793–803. [Google Scholar] [CrossRef] [Green Version]
- Compagno, L.J.V. Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date; Food and Agricultural Organization: Italy, Rome, 2002; Volume 2, pp. 83–85. [Google Scholar]
- Fu, D.; Roux, M.J.; Clarke, S.; Francis, M.; Dunn, A.; Hoyle, S. Pacific-wide sustainability risk assessment of bigeye thresher shark (Alopias superciliosus). In Proceedings of the Western Central Pacific Fisheries Commission Scientific Committee Thirteenth Regular Session, Cook Islands, Rarotonga, 9–17 August 2017. [Google Scholar]
- Liu, K.M.; Chiang, P.J.; Chen, C.T. Age and growth estimates of the bigeye thresher shark (Alopias superciliosus) in northeastern Taiwan waters. Fish. Bull. 1998, 96, 482–491. [Google Scholar]
- Tsai, W.P.; Chang, Y.J.; Liu, K.M. Development and testing of a Bayesian population model for the bigeye thresher shark, Alopias superciliosus, in an area subset of the western North Pacific. Fish. Manag. Ecol. 2019, 26, 269–294. [Google Scholar] [CrossRef]
- Amorim, A.; Baum, J.; Cailliet, G.M.; Clò, S.; Clarke, S.C.; Fergusson, I.; Gonzalez, M.; Macias, D.; Mancini, P.; Mancusi, C.; et al. Alopias superciliosus. The IUCN Red List of Threatened Species; IUCN: Cambridge, UK, 2009. [Google Scholar]
- CITES Animals Committee. Conservation and Management of Sharks and Stingrays: Other Animals Committee Tasks Related to Conservation and Management of Sharks. In Proceedings of the Sixteenth meeting of the Conference of the Parties, Johannesburg, South Africa, 24 September–4 October 2016. [Google Scholar]
- ICCAT Compliance Committee. CCAT Newsletter No. 12. In Proceedings of the Inter-Sessional Meeting of the Compliance Committee, Madrid, Spain, 24–26 February 2010. [Google Scholar]
- IOTC WPEB. Report of the Sixth Session of the IOTC Working Party on Ecosystems and Bycatch. In Proceedings of the Working Party on Ecosystems and Bycatch (WPEB) Meeting, Victoria, Seychelles, 27–30 October 2010. [Google Scholar]
- Vaughan, D.S.; Shertzer, K.W.; Smith, J.W. Gulf menhaden (Brevoortia patronus) in the US Gulf of Mexico: Fishery characteristics and biological reference points for management. Fish. Res. 2007, 83, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Restrepo, V.R.; Powers, J.E. Precautionary control rules in US fisheries management: Specification and performance. ICES J. Mar. Sci. 1999, 56, 846–852. [Google Scholar] [CrossRef]
- Tsai, W.P.; Sun, C.L.; Wang, S.P.; Liu, K.M. Evaluating the impacts of uncertainty on the estimation of biological reference points for the shortfin mako shark (Isurus oxyrinchus) in the Northwest Pacific Ocean. Mar. Freshw. Res. 2011, 62, 1383–1394. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Y.; Zhu, J.; Tian, S.; Chen, X. Evaluating effectiveness of biological reference points for bigeye tuna (Thunnus obesus) and yellowfin tuna (Thunnus albacares) fisheries in the Indian Ocean. Aquac. Fish. 2017, 2, 84–93. [Google Scholar] [CrossRef]
- Hoenig, J.M. Empirical use of longevity data to estimate mortality rates. Fish. Bull. 1983, 82, 898–903. [Google Scholar]
- Campana, S.; Marks, L.; Joyce, W.; Harley, S. Analytical Assessment of the Porbeagle Shark (Lamna nasus) Population in the Northwest Atlantic, with Estimates of Long-Term Sustainable Yield. Available online: https://www.researchgate.net/publication/268389652_Analytical_Assessment_of_the_Porbeagle_Shark_Lamna_nasus_Population_in_the_Northwest_Atlantic_with_Estimates_of_Long-term_Sustainable_Yield (accessed on 19 October 2020).
- Peterson, I.; Wroblewski, J.S. Mortality rate of fishes in the pelagic ecosystem. Can. J. Fish. Aquat. Sci. 1984, 41, 1117–1120. [Google Scholar] [CrossRef]
- Lorenzen, K. The relationship between body weight and natural mortality in juvenile and adult fish: A comparison of natural ecosystems and aquaculture. J. Fish Biol. 1996, 49, 627–647. [Google Scholar] [CrossRef]
- Ricker, W.E. Computation and interpretation of biological statistics of fish population. J. Wild. Manag. 1975, 191, 353–366. [Google Scholar]
- Fournier, D.A.; Skaug, H.J.; Ancheta, J.; Ianelli, J.; Magnusson, A.; Maunder, M.N.; Nielsen, A.; Sibert, J. AD Model Builder: Using automatic differentiation for statistical inference of highly parameterized complex nonlinear models. Optim. Methods Softw. 2012, 27, 233–249. [Google Scholar] [CrossRef] [Green Version]
- Best, N.G.; Cowles, M.K.; Vines, K. CODA: Convergence Diagnosis and Output Analysis Software for Gibbs Sampling Output, Version 0.30; MRC Biostatistics Unit: Cambridge, UK, 1995; p. 52. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing. Available online: http://softlibre.unizar.es/manuales/aplicaciones/r/fullrefman.pdf (accessed on 19 October 2020).
- Thompson, W.F.; Bell, F.H. Biological statistics of the Pacific halibut fishery 2. Effect of changes in intensity upon total yield and yield per unit of gear. Rep. Int. Fish. Comm. 1934, 8, 1–49. [Google Scholar]
- Goodyear, C.P. Spawning Stock Biomass Per Recruit in Fisheries Management: Foundation and Current Use. Available online: https://www.researchgate.net/publication/251832024_Spawning_Stock_Biomass_per_Recruit_in_Fisheries_Management_Foundation_and_Current_Use (accessed on 19 October 2020).
- Chen, C.T.; Liu, K.M.; Chang, Y.C. Reproductive biology of the bigeye thresher shark, Alopias superciliosus (Lowe, 1839) (Chondrichthyes: Alopiidae), in the northwestern Pacific. Ichthy. Res. 1997, 44, 27–235. [Google Scholar] [CrossRef]
- Liu, K.M.; Chen, C.T.; Liao, T.H.; Joung, S.J. Age, growth, and reproduction of the pelagic thresher shark, Alopias pelagicus in the northwestern Pacific. Copeia 1999, 1, 68–74. [Google Scholar] [CrossRef]
- Caswell, H. Matrix Population Models. Construction, Analysis, and Interpretation, 2nd ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2001; p. 722. [Google Scholar]
- Mollet, H.F.; Cailliet, G.M. Comparative population demography of elasmobranches using life history tables, Leslie matrices and stage-based matrix models. Mar. Freshw. Res. 2002, 53, 503–516. [Google Scholar] [CrossRef]
- Tsai, W.P.; Liu, K.M.; Joung, S.J. Demographic analysis of the pelagic thresher shark (Alopias pelagicus) in the Northwestern Pacific using a stochastic stage-based model. Mar. Freshw. Res. 2010, 61, 1056–1066. [Google Scholar] [CrossRef]
- Cortés, E. Incorporating Uncertainty into Demographic Modeling: Application to Shark Populations and their conservation. Conser. Biol. 2002, 16, 1048–1062. [Google Scholar] [CrossRef]
- Hood, G. Poptools [Computer Software]; Pest Animal Control Co-operative Research Center(CSIRO): Canberra, Australia, 2004. [Google Scholar]
- Clarke., S.; Hoyle, S. Development of limit reference points for elasmobranchs. In Proceedings of the Scientific Committee Tenth Regular Session, Republic of the Marshall Islands, Majuro, Marshall Islands, 6–14 August 2014. [Google Scholar]
- Cortés, E. Demographic analysis as an aid in shark stock assessment and management. Fish. Res. 1998, 39, 199–208. [Google Scholar] [CrossRef]
- Tsai, W.P.; Sun, C.L.; Punt, A.E.; Liu, K.M. Demographic analysis of the shortfin mako shark (Isurus oxyrinchus) in the Northwest Pacific using a two-sex stage-based matrix model. ICES J. Mar. Sci. 2014, 71, 1604–1618. [Google Scholar] [CrossRef] [Green Version]
- Romine, J.G.; Musick, J.A.; Burgess, G.H. Demographic analyses of the dusky shark, Carcharhinus obscurus, in the Northwest Atlantic incorporating hooking mortality estimates and revised reproductive parameters. Environ. Biol. Fish. 2009, 84, 277–289. [Google Scholar] [CrossRef]
- Jensen, A.L. Beverton and Holt life history invariants result from optimal trade-off of reproduction and survival. Can. J. Fish Aquat. Sci. 1996, 53, 820–822. [Google Scholar] [CrossRef]
- Pauly, D. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. ICES J. Mar. Sci. 1980, 39, 175–192. [Google Scholar] [CrossRef]
- Chen, P.; Yuan, W. Demographic analysis based on the growth parameter of sharks. Fish. Res. 2006, 78, 374–379. [Google Scholar] [CrossRef]
- Liu, K.M.; Chin, C.P.; Chen, C.H.; Chang, J.H. Estimating finite rate of population increase for sharks based on vital parameters. PLoS ONE 2015, 10, e0143008. [Google Scholar] [CrossRef]
- Natanson, L.J.; Skomal, G.B.; Hoffmann, S.L.; Porter, M.E.; Goldman, K.J.; Serra, D. Age and growth of sharks: Do vertebral band pairs record age? Mar. Freshw. Res. 2018, 69, 1440–1452. [Google Scholar] [CrossRef]
- Musick, J.A. Life in the Slow Lane: Ecology and Conservation of Long-Lived Marine Animals; American Fisheries Society: Bethesda, MD, USA, 1999; pp. 1–10. [Google Scholar]
- Otway, N.M.; Bradshaw, C.J.A.; Harcourt, R.G. Estimating the rate of quasi-extinction of the Australian grey nurse shark (Carcharias taurus) population using deterministic age- and stage-classified models. Biol. Conser. 2004, 119, 341–350. [Google Scholar] [CrossRef]
- Hisano, M.; Connolly, S.R.; Robbins, W.D. Population growth rates of reef sharks with and without fishing on the Great Barrier Reef: Robust estimation with multiple models. PLoS ONE 2011, 6, e25028. [Google Scholar] [CrossRef] [Green Version]




| Parameter | Female |
|---|---|
| Sex ratio () 1 | |
| 0.5 | |
| 0.218 | |
| 0.262 | |
| 1 | |
| Length–weight relationship 2 | |
| 2.769 | |
| VBGE 3 | |
| 224.6 | |
| 0.092 | |
| −4.21 | |
| Maturity fraction 4 | |
| −0.747 | |
| 12 |
| Age | Weight | Case 1 | Case 2 | Case 3 | Case 4 |
|---|---|---|---|---|---|
| 1 | 12.336 | 0.115 | 0.132 | 0.182 | 0.199 |
| 2 | 18.698 | 0.115 | 0.132 | 0.164 | 0.177 |
| 3 | 26.049 | 0.115 | 0.132 | 0.151 | 0.160 |
| 4 | 34.166 | 0.115 | 0.132 | 0.141 | 0.148 |
| 5 | 42.839 | 0.115 | 0.132 | 0.133 | 0.139 |
| 6 | 51.872 | 0.115 | 0.132 | 0.127 | 0.132 |
| 7 | 61.099 | 0.115 | 0.132 | 0.122 | 0.126 |
| 8 | 70.373 | 0.115 | 0.132 | 0.118 | 0.121 |
| 9 | 79.575 | 0.115 | 0.132 | 0.114 | 0.116 |
| 10 | 88.608 | 0.115 | 0.132 | 0.111 | 0.113 |
| 11 | 97.396 | 0.115 | 0.132 | 0.109 | 0.110 |
| 12 | 105.880 | 0.115 | 0.132 | 0.106 | 0.107 |
| 13 | 114.017 | 0.115 | 0.132 | 0.104 | 0.105 |
| 14 | 121.778 | 0.115 | 0.132 | 0.103 | 0.103 |
| 15 | 129.144 | 0.115 | 0.132 | 0.101 | 0.101 |
| 16 | 136.106 | 0.115 | 0.132 | 0.100 | 0.100 |
| 17 | 142.660 | 0.115 | 0.132 | 0.099 | 0.098 |
| 18 | 148.812 | 0.115 | 0.132 | 0.098 | 0.097 |
| 19 | 154.567 | 0.115 | 0.132 | 0.097 | 0.096 |
| 20 | 159.940 | 0.115 | 0.132 | 0.096 | 0.095 |
| 21 | 164.943 | 0.115 | 0.132 | 0.095 | 0.094 |
| 22 | 169.592 | 0.115 | 0.132 | 0.095 | 0.094 |
| 23 | 173.905 | 0.115 | 0.132 | 0.094 | 0.093 |
| 24 | 177.899 | 0.115 | 0.132 | 0.093 | 0.092 |
| 25 | 181.593 | 0.115 | 0.132 | 0.093 | 0.092 |
| 26 | 185.005 | 0.115 | 0.132 | 0.093 | 0.091 |
| 27 | 188.152 | 0.115 | 0.132 | 0.092 | 0.091 |
| 28 | 191.053 | 0.115 | 0.132 | 0.092 | 0.090 |
| 29 | 193.723 | 0.115 | 0.132 | 0.092 | 0.090 |
| 30 | 196.179 | 0.115 | 0.132 | 0.091 | 0.090 |
| 31 | 198.437 | 0.115 | 0.132 | 0.091 | 0.089 |
| 32 | 200.510 | 0.115 | 0.132 | 0.091 | 0.089 |
| 33 | 202.413 | 0.115 | 0.132 | 0.091 | 0.089 |
| 34 | 204.159 | 0.115 | 0.132 | 0.090 | 0.089 |
| 35 | 205.760 | 0.115 | 0.132 | 0.090 | 0.088 |
| Mean | 132.264 | 0.115 | 0.132 | 0.107 | 0.109 |
| M | Case 1 | Case 2 | Case 3 | Case 4 |
| F | 0.504 | 0.497 | 0.507 | 0.506 |
| 8.796 | 8.795 | 8.798 | 8.797 | |
| 2.053 | 2.048 | 2.049 | 2.048 | |
| Age | Selectivity | |||
| 1 | 0.000 | 0.000 | 0.000 | 0.000 |
| 2 | 0.001 | 0.001 | 0.001 | 0.001 |
| 3 | 0.004 | 0.004 | 0.004 | 0.004 |
| 4 | 0.019 | 0.018 | 0.018 | 0.018 |
| 5 | 0.066 | 0.065 | 0.065 | 0.065 |
| 6 | 0.182 | 0.181 | 0.180 | 0.180 |
| 7 | 0.398 | 0.396 | 0.396 | 0.395 |
| 8 | 0.686 | 0.685 | 0.684 | 0.684 |
| 9 | 0.932 | 0.932 | 0.932 | 0.932 |
| 10 | 1.000 | 1.000 | 1.000 | 1.000 |
| 11 | 0.846 | 0.845 | 0.846 | 0.846 |
| 12 | 0.565 | 0.563 | 0.564 | 0.564 |
| 13 | 0.297 | 0.295 | 0.297 | 0.296 |
| 14 | 0.124 | 0.122 | 0.123 | 0.122 |
| 15 | 0.041 | 0.040 | 0.040 | 0.040 |
| 16 | 0.011 | 0.010 | 0.010 | 0.010 |
| 17 | 0.002 | 0.002 | 0.002 | 0.002 |
| 18 | 0.000 | 0.000 | 0.000 | 0.000 |
| 19 | 0.000 | 0.000 | 0.000 | 0.000 |
| 20 | 0.000 | 0.000 | 0.000 | 0.000 |
| 21 | 0.000 | 0.000 | 0.000 | 0.000 |
| 22 | 0.000 | 0.000 | 0.000 | 0.000 |
| 23 | 0.000 | 0.000 | 0.000 | 0.000 |
| 24 | 0.000 | 0.000 | 0.000 | 0.000 |
| 25 | 0.000 | 0.000 | 0.000 | 0.000 |
| 26 | 0.000 | 0.000 | 0.000 | 0.000 |
| 27 | 0.000 | 0.000 | 0.000 | 0.000 |
| 28 | 0.000 | 0.000 | 0.000 | 0.000 |
| 29 | 0.000 | 0.000 | 0.000 | 0.000 |
| 30 | 0.000 | 0.000 | 0.000 | 0.000 |
| 31 | 0.000 | 0.000 | 0.000 | 0.000 |
| 32 | 0.000 | 0.000 | 0.000 | 0.000 |
| 33 | 0.000 | 0.000 | 0.000 | 0.000 |
| 34 | 0.000 | 0.000 | 0.000 | 0.000 |
| 35 | 0.000 | 0.000 | 0.000 | 0.000 |
| Natural Mortality | Longevity | Reference Points | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Fcurr | YPR | F0.1 | Fmax | SPR(%) | FSPR35% | FSPR30% | Fcrit | ||
| Case 1 | amax = 35 | 0.504 | 28.327 | 0.437 | 0.975 | 8.578 | 0.211 | 0.243 | 0.139 |
| Case 2 | 0.497 | 25.160 | 0.455 | 1.143 | 9.169 | 0.213 | 0.245 | 0.079 | |
| Case 3 | 0.507 | 24.506 | 0.438 | 1.004 | 8.355 | 0.211 | 0.242 | 0.116 | |
| Case 4 | 0.506 | 23.450 | 0.440 | 1.031 | 8.405 | 0.211 | 0.242 | 0.102 | |
| Case 1 | amax = 30 | 0.497 | 25.162 | 0.455 | 1.142 | 9.260 | 0.214 | 0.246 | 0.070 |
| Case 2 | 0.487 | 21.514 | 0.480 | 1.620 | 10.078 | 0.217 | 0.249 | - | |
| Case 3 | 0.507 | 24.506 | 0.438 | 1.004 | 8.467 | 0.211 | 0.243 | 0.101 | |
| Case 4 | 0.506 | 23.450 | 0.440 | 1.031 | 8.520 | 0.212 | 0.243 | 0.087 | |
| Case 1 | amax = 25 | 0.487 | 21.402 | 0.481 | 1.648 | 10.284 | 0.218 | 0.251 | - |
| Case 2 | 0.474 | 17.293 | 0.519 | 4.199 | 11.493 | 0.223 | 0.256 | - | |
| Case 3 | 0.507 | 24.506 | 0.438 | 1.004 | 8.688 | 0.213 | 0.245 | 0.074 | |
| Case 4 | 0.506 | 23.450 | 0.440 | 1.031 | 8.744 | 0.213 | 0.245 | 0.060 | |
| Natural Mortality | Longevity | Population Increase Rate | |
|---|---|---|---|
| F = 0 | F = Fcurr | ||
| Case 1 | amax = 35 | 1.039 | 0.913 |
| Case 2 | 1.022 | 0.900 | |
| Case 3 | 1.031 | 0.911 | |
| Case 4 | 1.027 | 0.909 | |
| Case 1 | amax = 30 | 1.020 | 0.892 |
| Case 2 | 0.998 | 0.876 | |
| Case 3 | 1.029 | 0.901 | |
| Case 4 | 1.024 | 0.898 | |
| Case 1 | amax = 25 | 0.993 | 0.863 |
| Case 2 | 0.964 | 0.840 | |
| Case 3 | 1.022 | 0.885 | |
| Case 4 | 1.018 | 0.882 | |
| Scenario | Type of F | Lower CL | Upper CL | r | Lower CL | Upper CL | |
|---|---|---|---|---|---|---|---|
| 1 | F = 0 | 1.023 | 1.010 | 1.039 | 0.023 | 0.010 | 0.039 |
| 2 | Fcur | 0.906 | 0.894 | 0.915 | −0.098 | −0.112 | −0.089 |
| 3 | F0.1 | 0.919 | 0.905 | 0.928 | −0.085 | −0.100 | −0.075 |
| 4 | Fmax | 0.815 | 0.789 | 0.829 | −0.204 | −0.237 | −0.187 |
| 5 | FSPR35% | 0.969 | 0.957 | 0.981 | −0.031 | −0.044 | −0.019 |
| 6 | FSPR30% | 0.962 | 0.950 | 0.974 | −0.039 | −0.052 | −0.027 |
| 7 | Fcrit | 0.995 | 0.974 | 1.017 | −0.006 | −0.026 | 0.017 |
| Natural Mortality | Longevity | Fcrit | Corresponding SPR% |
|---|---|---|---|
| Case 1 | amax = 35 | 0.139 | 49.907 |
| Case 2 | 0.079 | 67.644 | |
| Case 3 | 0.116 | 55.989 | |
| Case 4 | 0.102 | 60.044 | |
| Case 1 | amax = 30 | 0.070 | 70.723 |
| Case 2 | - | - | |
| Case 3 | 0.101 | 60.447 | |
| Case 4 | 0.087 | 64.896 | |
| Case 1 | amax = 25 | - | - |
| Case 2 | - | - | |
| Case 3 | 0.074 | 69.099 | |
| Case 4 | 0.060 | 74.262 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, W.-P.; Liu, K.-M.; Chang, Y.-J. Evaluation of Biological Reference Points for Conservation and Management of the Bigeye Thresher Shark, Alopias superciliosus, in the Northwest Pacific. Sustainability 2020, 12, 8646. https://doi.org/10.3390/su12208646
Tsai W-P, Liu K-M, Chang Y-J. Evaluation of Biological Reference Points for Conservation and Management of the Bigeye Thresher Shark, Alopias superciliosus, in the Northwest Pacific. Sustainability. 2020; 12(20):8646. https://doi.org/10.3390/su12208646
Chicago/Turabian StyleTsai, Wen-Pei, Kwang-Ming Liu, and Yi-Jay Chang. 2020. "Evaluation of Biological Reference Points for Conservation and Management of the Bigeye Thresher Shark, Alopias superciliosus, in the Northwest Pacific" Sustainability 12, no. 20: 8646. https://doi.org/10.3390/su12208646
APA StyleTsai, W. -P., Liu, K. -M., & Chang, Y. -J. (2020). Evaluation of Biological Reference Points for Conservation and Management of the Bigeye Thresher Shark, Alopias superciliosus, in the Northwest Pacific. Sustainability, 12(20), 8646. https://doi.org/10.3390/su12208646