
Discovery of Polyvalent Myovirus (vB_STM-2) Phage as a Natural Antimicrobial System to Lysis and Biofilm Removal of Salmonella Typhimurium Isolates from Various Food Sources


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Salmonella Typhimurium Isolates Sources in the Study
2.2. Inoculums Preparation of Isolates
2.3. Confirm the Identification of the Salmonella Typhimurium Used
2.4. Statuses of Antibiotic-Resistant Profiles of Salmonella Typhimurium Used
2.5. Biofilm Forming Capacity of Salmonella Typhimurium Isolates
2.6. Isolation and Purification of Salmonella Typhimurium Phages
2.6.1. Isolation Source
2.6.2. Isolation and Enrichment
2.7. Phage Detection
- Spot-Assay Technique
- Plaque Assay Technique
- Purification, Propagation, and Titration
- Host Range Study
- Transmission Electron Microscopy (TEM)
- Thermo and pH Stability of vB_STM-2 Phage
- Biofilm Removal Activity of the vB_STM-2 Phage
2.8. Bacterial Challenge Measurements
In Broth Medium (Culture Clearing)
2.9. Preparation of the Food Sample Used in the Study
2.10. Effectiveness and Stability of the vB_STM-2 Phage in Food Preservation
2.11. Recovery of Inoculated Salmonella from Food
2.12. Statistical Analysis
3. Results
3.1. Bacterial Isolates, Identification, Antibiotic-Resistant and Biofilm Behavior
3.2. Most Potent Lytic Phages Isolation and Characterization
3.3. Selection of a Broad Host Spectrum and Polyvalent Phages
3.4. Thermo and pH Stability of the vB_STM-2 Phage
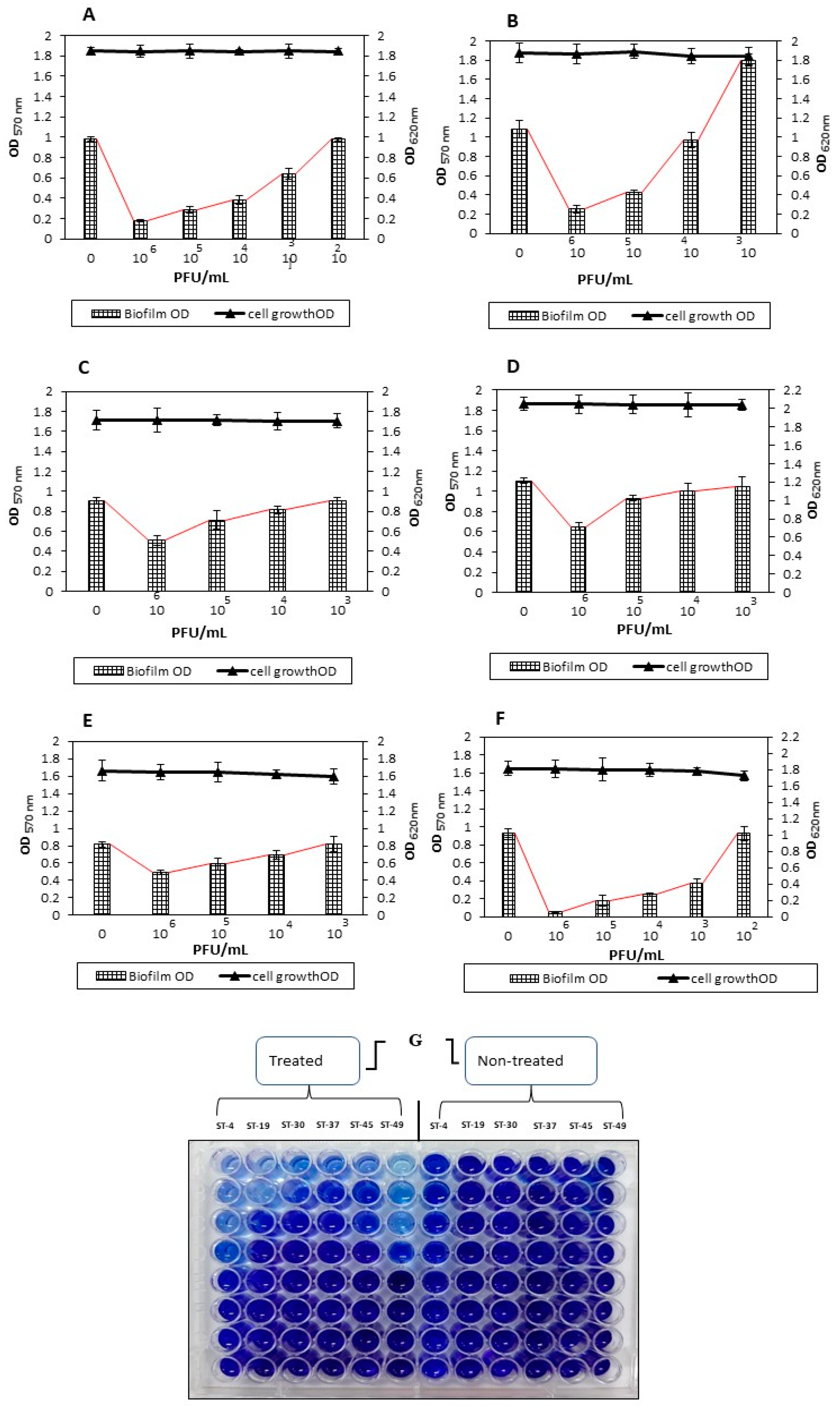
3.5. Biofilm Removal Effect of the vB_STM-2 Phage
3.6. The Ability of the vB_STM-2 Phage as a Natural Antibacterial
Lytic Potency in TSB Medium (Culture Clearance)
3.7. Control of Salmonella in Food and Phage Stability
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Musyoka, J.N.; Abong’, G.O.; Mbogo, D.M.; Fuchs, R.; Low, J.; Heck, S.; Muzhingi, T. Effects of Acidification and Preservatives on Microbial Growth during Storage of Orange Fleshed Sweet Potato Puree. Int. J. Food Sci. 2018, 2018, 8410747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamas, A.; Regal, P.; Vazquez, B.; Miranda, J.; Cepeda, A.; Franco, C.M. Salmonella and Campylobacter biofilm formation: A comparative assessment from farm to fork. J. Sci. Food Agric. 2018, 98, 4014–4032. [Google Scholar] [CrossRef]
- Abu-Shahba, M.S.; Mansour, M.M.; Mohamed, H.I.; Sofy, M.R. Comparative Cultivation and Biochemical Analysis of Iceberg Lettuce Grown in Sand Soil and Hydroponics with or without Microbubbles and Macrobubbles. J. Soil Sci. Plant Nutr. 2021, 21, 389–403. [Google Scholar] [CrossRef]
- Galletti, J.; Tobaldini-Valerio, F.K.; Silva, S.; Kioshima, É.S.; Pereira, L.T.; Bruschi, M.; Negri, M.; Svidzinski, T.I.E. Antibiofilm activity of propolis extract on Fusarium species from onychomycosis. Futur. Microbiol. 2017, 12, 1311–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pobiega, K.; Przybył, J.L.; Żubernik, J.; Gniewosz, M. Prolonging the Shelf Life of Cherry Tomatoes by Pullulan Coating with Ethanol Extract of Propolis During Refrigerated Storage. Food Bioprocess Technol. 2020, 13, 1447–1461. [Google Scholar] [CrossRef]
- CDC. Salmonella Homepage. 2016. Available online: https://www.cdc.gov/salmonella/index.html (accessed on 15 January 2021).
- Demirbilek, S.K. Salmonellosis in Animals. In Salmonella—A Re-Emerging Pathogen; IntechOpen Limited: London, UK, 2018. [Google Scholar]
- Ferrari, R.; Rosario, D.K.A.; Neto, A.C.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Junior, C.A. Worldwide Epidemiology of Salmonella Serovars in Animal-Based Foods: A Meta-analysis. Appl. Environ. Microbiol. 2019, 85, e00591-19. [Google Scholar] [CrossRef] [Green Version]
- Mueller-Doblies, D.; Speed, K.C.R.; Kidd, S.; Davies, R.H. Salmonella Typhimurium in livestock in Great Britain—Trends observed over a 32-year period. Epidemiol. Infect. 2018, 146, 409–422. [Google Scholar] [CrossRef] [Green Version]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The Global Burden of Nontyphoidal Salmonella Gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, S.M.; Keel, K.; Sanchez, S.; Trees, E.; Gerner-Smidt, P.; Adams, J.K.; Cheng, Y.; Ray, A.; Martin, G.; Presotto, A.; et al. Epidemiology of a Salmonella enterica subsp. enterica Serovar Typhimurium Strain Associated with a Songbird Outbreak. Appl. Environ. Microbiol. 2012, 78, 7290–7298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.; Hoekstra, R.M.; Schroeder, C.M.; Pires, S.M.; Ong, K.L.; Hartnett, E.; Naugle, A.; Harman, J.; Bennett, P.; Cieslak, P.; et al. Application of Bayesian Techniques to Model the Burden of Human Salmonellosis Attributable to U.S. Food Commodities at the Point of Processing: Adaptation of a Danish Model. Foodborne Pathog. Dis. 2011, 8, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Marchello, C.S.; Dale, A.P.; Pisharody, S.; Rubach, M.P.; Crump, J.A. A Systematic Review and Meta-analysis of the Prevalence of Community-Onset Bloodstream Infections among Hospitalized Patients in Africa and Asia. Antimicrob. Agents Chemother. 2019, 64, 01974-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Carvalho, C.C.C.R. Marine Biofilms: A Successful Microbial Strategy with Economic Implications. Front. Mar. Sci. 2018, 5, 126. [Google Scholar] [CrossRef] [Green Version]
- Mah, T.-F. Biofilm-specific antibiotic resistance. Futur. Microbiol. 2012, 7, 1061–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Geng, M.; Bai, L. Targeting Biofilms Therapy: Current Research Strategies and Development Hurdles. Microorganisms 2020, 8, 1222. [Google Scholar] [CrossRef]
- Agyare, C.; Boamah, V.E.; Zumbi, C.N.; Osei, F.B. Antibiotic Use in Poultry Production and Its Effects on Bacterial Resistance. In Antimicrobial Resistance—A Global Threat; Kumar, Y., Ed.; IntechOpen: London, UK, 2018; pp. 33–51. ISBN 978-1-78985-783-2. [Google Scholar]
- Moye, Z.D.; Woolston, J.; Sulakvelidze, A. Bacteriophage Applications for Food Production and Processing. Viruses 2018, 10, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeLièvre, V.; Besnard, A.; Schlusselhuber, M.; Desmasures, N.; Dalmasso, M. Phages for biocontrol in foods: What opportunities for Salmonella sp. control along the dairy food chain? Food Microbiol. 2019, 78, 89–98. [Google Scholar] [CrossRef]
- García-Anaya, M.C.; Sepulveda, D.R.; Sáenz-Mendoza, A.I.; Rios-Velasco, C.; Zamudio-Flores, P.B.; Acosta-Muñiz, C.H. Phages as biocontrol agents in dairy products. Trends Food Sci. Technol. 2020, 95, 10–20. [Google Scholar] [CrossRef]
- Tang, F.; Zhang, P.; Zhang, Q.; Xue, F.; Ren, J.; Sun, J.; Qu, Z.; Zhuge, X.; Li, D.; Wang, J.; et al. Isolation and characterization of a broad-spectrum phage of multiple drug resistant Salmonella and its therapeutic utility in mice. Microb. Pathog. 2019, 126, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Abhisingha, M.; Dumnil, J.; Pitaksutheepong, C. Efficiency of phage cocktail to reduce Salmonella Typhimurium on chicken meat during low temperature storage. LWT 2020, 129, 109580. [Google Scholar] [CrossRef]
- Li, Z.; Ma, W.; Li, W.; Ding, Y.; Zhang, Y.; Yang, Q.; Wang, J.; Wang, X. A broad-spectrum phage controls multidrug-resistant Salmonella in liquid eggs. Food Res. Int. 2020, 132, 109011. [Google Scholar] [CrossRef]
- Petsong, K.; Benjakul, S.; Vongkamjan, K. Evaluation of storage conditions and efficiency of a novel microencapsulated Salmonella phage cocktail for controlling S. enteritidis and S. typhimurium in-vitro and in fresh foods. Food Microbiol. 2019, 83, 167–174. [Google Scholar] [CrossRef]
- Ackermann, H.-W.; Audurier, A.; Berthiaume, L.; Jones, L.A.; Mayo, J.A.; Vidaver, A.K. Guidelines for Bacteriophage Characterization. Adv. Appl. Microbiol. 1978, 23, 1–24. [Google Scholar] [CrossRef]
- Park, M.; Lee, J.-H.; Shin, H.; Kim, M.; Choi, J.; Kang, D.-H.; Heu, S.; Ryu, S. Characterization and Comparative Genomic Analysis of a Novel Bacteriophage, SFP10, Simultaneously Inhibiting both Salmonella enterica and Escherichia coli O157:H. Appl. Environ. Microbiol. 2011, 78, 58–69. [Google Scholar] [CrossRef] [Green Version]
- Parra, B.; Robeson, J. Selection of polyvalent bacteriophages infecting Salmonella enterica serovar Choleraesuis. Electron. J. Biotechnol. 2016, 21, 72–76. [Google Scholar] [CrossRef] [Green Version]
- Malki, K.; Kula, A.; Bruder, K.; Sible, E.; Hatzopoulos, T.; Steidel, S.; Watkins, S.C.; Putonti, C. Bacteriophages isolated from Lake Michigan demonstrate broad host-range across several bacterial phyla. Virol. J. 2015, 12, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hungaro, H.M.; Mendonça, R.C.S.; Gouvêa, D.M.; Vanetti, M.C.D.; de Oliveira Pinto, C.L. Use of bacteriophages to reduce Salmonella in chicken skin in comparison with chemical agents. Food Res. Int. 2013, 52, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Augustine, J.; Bhat, S.G. Biocontrol of salmonella enteritidis in spiked chicken cuts by lytic bacteriophages φsp-1 and φsp-3. J. Basic Microbiol. 2015, 55, 500–503. [Google Scholar] [CrossRef] [PubMed]
- Bigwood, T.; Hudson, J.A.; Billington, C.; Carey-Smith, G.V.; Heinemann, J.A. Phage inactivation of foodborne pathogens on cooked and raw meat. Food Microbiol. 2008, 25, 400–406. [Google Scholar] [CrossRef]
- Hooton, S.P.; Atterbury, R.; Connerton, I.F. Application of a bacteriophage cocktail to reduce Salmonella Typhimurium U288 contamination on pig skin. Int. J. Food Microbiol. 2011, 151, 157–163. [Google Scholar] [CrossRef]
- Kocharunchitt, C.; Ross, T.; McNeil, D. Use of bacteriophages as biocontrol agents to control Salmonella associated with seed sprouts. Int. J. Food Microbiol. 2009, 128, 453–459. [Google Scholar] [CrossRef]
- Modi, R.; Hirvi, Y.; Hill, A.; Griffiths, M.W. Effect of Phage on Survival of Salmonella Enteritidis during Manufacture and Storage of Cheddar Cheese Made from Raw and Pasteurized Milk. J. Food Prot. 2001, 64, 927–933. [Google Scholar] [CrossRef]
- Silliker, J.; Taylor, W. Isolation of salmonellae from food samples: Ii. The effect of added food samples upon the performance of enrichment broths. Appl. Microbiol. 1958, 6, 228–232. [Google Scholar] [CrossRef]
- ISO. Microbiology of food and animal feeding stuffs-horizontal method for the detection of Salmonella spp. Amendment 1: Anex d: Detection of Salmonella spp. In Animal Faeces and in Environmental Samples from the Primary Production Stage; ISO: Geneva, Switzerland, 2007. [Google Scholar]
- Sieuwerts, S.; De Bok, F.; Mols, E.; De Vos, W.; Vlieg, J.V.H. A simple and fast method for determining colony forming units. Lett. Appl. Microbiol. 2008, 47, 275–278. [Google Scholar] [CrossRef]
- Søgaard, M.; Nørgaard, M.; Schønheyder, H.C. First Notification of Positive Blood Cultures and the High Accuracy of the Gram Stain Report. J. Clin. Microbiol. 2007, 45, 1113–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, W.; Waltman, R.; Gast, K.; Mallinson, E. Salmonellosis. A Laboratory Manual for the Isolation and Identification of Avian Pathogens, 4th ed.; American Association of Avian Pathologists: Kenett Square, PA, USA, 1998. [Google Scholar]
- Bossé, J. Manual of standards for diagnostic tests and vaccines. Can. Vet. J. 2000, 39, 183. [Google Scholar]
- Kauffman, G. Kauffmann white scheme. J. Acta. Path. Microbiol. Sci. 1974, 61, 385. [Google Scholar]
- Santiviago, C.; Toro, C.S.; Bucarey, S.A.; Mora, G.C. A chromosomal region surrounding the ompD porin gene marks a genetic difference between Salmonella typhi and the majority of Salmonella serovars. Microbiology 2001, 147, 1897–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, A. Antibiotic susceptibility testing by a standardized single disc method. Am. J. Clin. Pathol. 1966, 45, 149–158. [Google Scholar] [CrossRef]
- NCCLS/CLSI. Performance Standards for Antimicrobial Susceptibility Testing; National Committee for Clinical Laboratory Standards/Clinical and Laboratory Standards Institute: Wayne, MI, USA, 2007. [Google Scholar]
- Stepanović, S.; Vuković, D.; Hola, V.; DI Bonaventura, G.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Bibi, Z.; Abbas, Z.; Rehman, S.U. The phage P.E1isolated from hospital sewage reduces the growth of Escherichia coli. Biocontrol Sci. Technol. 2015, 26, 181–188. [Google Scholar] [CrossRef]
- Capra, M.; Quiberoni, A.; Reinheimer, J. Thermal and chemical resistance of Lactobacillus casei and Lactobacillus paracasei bacteriophages. Lett. Appl. Microbiol. 2004, 38, 499–504. [Google Scholar] [CrossRef]
- Sangha, K.K.; Kumar, B.V.S.; Agrawal, R.K.; Deka, D.; Verma, R. Proteomic Characterization of Lytic Bacteriophages of Staphylococcus aureus Isolated from Sewage Affluent of India. Int. Sch. Res. Not. 2014, 2014, 265298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirzaei, M.K.; Nilsson, A.S. Correction: Isolation of Phages for Phage Therapy: A Comparison of Spot Tests and Efficiency of Plating Analyses for Determination of Host Range and Efficacy. PLoS ONE 2015, 10, e0127606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackermann, H.-W. Bacteriophage Electron Microscopy. Adv. Appl. Microbiol. 2012, 82, 1–32. [Google Scholar] [CrossRef]
- Accolas, J.-P.; Spillmann, H. The Morphology of Six Bacteriophages of Streptococcus thermophilus. J. Appl. Bacteriol. 1979, 47, 135–144. [Google Scholar] [CrossRef]
- Philipson, L.; Albertsson, P.; Frick, G. The purification and concentration of viruses by aqueous polymer phase systems. Virology 1960, 11, 553–571. [Google Scholar] [CrossRef]
- Jamalludeen, N.; Johnson, R.P.; Friendship, R.; Kropinski, A.; Lingohr, E.J.; Gyles, C.L. Isolation and characterization of nine bacteriophages that lyse O149 enterotoxigenic Escherichia coli. Veter Microbiol. 2007, 124, 47–57. [Google Scholar] [CrossRef]
- Bekir, K.; Ben Abdallah, F.; Ellafi, A.; Bakhrouf, A. Adherence assays and slime production of Staphylococcus aureus strains after their incubation in seawater microcosms. Ann. Microbiol. 2011, 61, 819–823. [Google Scholar] [CrossRef]
- Else, T.A.; Pantle, C.R.; Amy, P.S. Boundaries for Biofilm Formation: Humidity and Temperature. Appl. Environ. Microbiol. 2003, 69, 5006–5010. [Google Scholar] [CrossRef] [Green Version]
- Kostaki, M.; Chorianopoulos, N.; Braxou, E.; Nychas, G.-J.; Giaouris, E. Differential Biofilm Formation and Chemical Disinfection Resistance of Sessile Cells of Listeria monocytogenes Strains under Monospecies and Dual-Species (with Salmonella enterica) Conditions. Appl. Environ. Microbiol. 2012, 78, 2586–2595. [Google Scholar] [CrossRef] [Green Version]
- Kusumaningrum, H.D.; Van Asselt, E.D.; Beumer, R.R.; Zwietering, M.H. A Quantitative Analysis of Cross-Contamination of Salmonella and Campylobacter spp. via Domestic Kitchen Surfaces. J. Food Prot. 2004, 67, 1892–1903. [Google Scholar] [CrossRef]
- FDA. Bacteriological Analytical Manual; Food and Drug Administration: Arlington, VA, USA, 2002.
- Spricigo, D.A.; Bardina, C.; Cortés, P.; Llagostera, M. Use of a bacteriophage cocktail to control Salmonella in food and the food industry. Int. J. Food Microbiol. 2013, 165, 169–174. [Google Scholar] [CrossRef]
- Tomat, D.; Casabonne, C.; Aquili, V.; Balagué, C.; Quiberoni, A. Evaluation of a novel cocktail of six lytic bacteriophages against Shiga toxin-producing Escherichia coli in broth, milk and meat. Food Microbiol. 2018, 76, 434–442. [Google Scholar] [CrossRef]
- Sofy, A.; Sofy, M.; Hmed, A.; Dawoud, R.; Alnaggar, A.; Soliman, A.; El-Dougdoug, N. Ameliorating the Adverse Effects of Tomato mosaic tobamovirus Infecting Tomato Plants in Egypt by Boosting Immunity in Tomato Plants Using Zinc Oxide Nanoparticles. Molecules 2021, 26, 1337. [Google Scholar] [CrossRef]
- Sofy, A.; Sofy, M.; Hmed, A.; Dawoud, R.; Refaey, E.; Mohamed, H.; El-Dougdoug, N. Molecular Characterization of the Alfalfa mosaic virus Infecting Solanum melongena in Egypt and the Control of Its Deleterious Effects with Melatonin and Salicylic Acid. Plants 2021, 10, 459. [Google Scholar] [CrossRef]
- Fong, K.; Mu, K.; Rheault, J.-G.; Levesque, R.C.; Kitts, D.D.; Delaquis, P.; Goodridge, L.; Wang, S. Bacteriophage-Insensitive Mutants of Antimicrobial-Resistant Salmonella enterica are Altered in their Tetracycline Resistance and Virulence in Caco-2 Intestinal Cells. Int. J. Mol. Sci. 2020, 21, 1883. [Google Scholar] [CrossRef] [Green Version]
- Heredia, N.; García, S. Animals as sources of food-borne pathogens: A review. Anim. Nutr. 2018, 4, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Browne, A.J.; Hamadani, B.H.K.; Kumaran, E.; Rao, P.; Longbottom, J.; Harriss, E.; Moore, C.; Dunachie, S.; Basnyat, B.; Baker, S.; et al. Drug-resistant enteric fever worldwide, 1990 to 2018: A systematic review and meta-analysis. BMC Med. 2020, 18, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Biswas, S.; Gu, G.; Elbediwi, M.; Li, Y.; Yue, M. Characterization of Multidrug Resistance Patterns of Emerging Salmonella enterica Serovar Rissen along the Food Chain in China. Antibiotics 2020, 9, 660. [Google Scholar] [CrossRef]
- Elshaarawy, R.F.; Mustafa, F.H.; Sofy, A.R.; Hmed, A.A.; Janiak, C. A new synthetic antifouling coatings integrated novel aminothiazole-functionalized ionic liquids motifs with enhanced antibacterial performance. J. Environ. Chem. Eng. 2019, 7, 102800. [Google Scholar] [CrossRef]
- Michael, G.; Schwarz, S. Antimicrobial resistance in zoonotic nontyphoidal Salmonella: An alarming trend? Clin. Microbiol. Infect. 2016, 22, 968–974. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2016. EFSA J. 2018, 16, e05182. [Google Scholar] [CrossRef]
- Eng, S.-K.; Pusparajah, P.; Ab Mutalib, N.-S.; Ser, H.-L.; Chan, K.-G.; Lee, L.-H. Salmonella: A review on pathogenesis, epidemiology and antibiotic resistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar] [CrossRef] [Green Version]
- Megahed, A.; Dougdoug, K.A.E.; Othman, B.; Lashin, S.; Ibrahim, M.; Sofy, A. A New Egyptian Satellite Strain of Cucumber Mosaic Cucumovirus. Int. J. Virol. 2012, 8, 240–257. [Google Scholar] [CrossRef] [Green Version]
- Megahed, A.; El-Dougdou, K.A.; Othman, B.; Lashin, S.; Ibrahim, M.; Sofy, A. Induction of Resistance in Tomato Plants Against Tomato mosaic tobamovirus Using Beneficial Microbial Isolates. Pak. J. Biol. Sci. 2013, 16, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Stefanucci, A.; Amato, J.; Brancaccio, D.; Pagano, B.; Randazzo, A.; Santoro, F.; Mayol, L.; Learte-Aymamí, S.; Rodriguez, J.; Mascareñas, J.L.; et al. A novel β-hairpin peptide derived from the ARC repressor selectively interacts with the major groove of B-DNA. Bioorganic Chem. 2021, 112, 104836. [Google Scholar] [CrossRef]
- Zare, N.; Makvandi, P.; Ashtari, B.; Rossi, F.; Motahari, A.; Perale, G. Progress in Conductive Polyaniline-Based Nanocomposites for Biomedical Applications: A Review. J. Med. Chem. 2020, 63, 1–22. [Google Scholar] [CrossRef]
- Zare, E.N.; Jamaledin, R.; Naserzadeh, P.; Afjeh-Dana, E.; Ashtari, B.; Hosseinzadeh, M.; Vecchione, R.; Wu, A.; Tay, F.R.; Borzacchiello, A.; et al. Metal-Based Nanostructures/PLGA Nanocomposites: Antimicrobial Activity, Cytotoxicity, and Their Biomedical Applications. ACS Appl. Mater. Interfaces 2019, 12, 3279–3300. [Google Scholar] [CrossRef]
- Li, X.-H.; Lee, J.-H. Antibiofilm agents: A new perspective for antimicrobial strategy. J. Microbiol. 2017, 55, 753–766. [Google Scholar] [CrossRef]
- Tasneem, U.; Yasin, N.; Nisa, I.; Shah, F.; Rasheed, U.; Momin, F.; Zaman, S.; Qasim, M. Biofilm producing bacteria: A serious threat to public health in developing countries. J. Food Sci. Nutr. 2018, 01, 25–31. [Google Scholar] [CrossRef]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control. 2019, 8, 76. [Google Scholar] [CrossRef]
- Jung, L.-S.; Ding, T.; Ahn, J. Evaluation of lytic bacteriophages for control of multidrug-resistant Salmonella Typhimurium. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 66. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.; Kavanagh, K.A. Evaluation of metal-based antimicrobial compounds for the treatment of bacterial pathogens. J. Med Microbiol. 2021, 70, 001363. [Google Scholar] [CrossRef]
- Guenther, S.; Herzig, O.; Fieseler, L.; Klumpp, J.; Loessner, M.J. Biocontrol of Salmonella Typhimurium in RTE foods with the virulent bacteriophage FO1-E2. Int. J. Food Microbiol. 2012, 154, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.; Zhou, Y.; Liang, L.; Nime, I.; Liu, K.; Yan, T.; Wang, X.; Li, J. Application of a Phage Cocktail for Control of Salmonella in Foods and Reducing Biofilms. Viruses 2019, 11, 841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, T.K.; Koeris, M.S. The next generation of bacteriophage therapy. Curr. Opin. Microbiol. 2011, 14, 524–531. [Google Scholar] [CrossRef]
- Chan, B.K.; Abedon, S.T. Phage Therapy Pharmacology. Adv. Appl. Microbiol. 2012, 78, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Sofy, A.R.; El Haliem, N.F.A.; Refaey, E.E.; Hmed, A.A. Polyvalent Phage CoNShP-3 as a Natural Antimicrobial Agent Showing Lytic and Antibiofilm Activities against Antibiotic-Resistant Coagulase-Negative Staphylococci Strains. Foods 2020, 9, 673. [Google Scholar] [CrossRef]
- Sofy, A.R.; Aboseidah, A.A.; El-Morsi, E.-S.; Azmy, H.A.; Hmed, A.A. Evaluation of Antibacterial and Antibiofilm Activity of New Antimicrobials as an Urgent Need to Counteract Stubborn Multidrug-resistant Bacteria. J. Pure Appl. Microbiol. 2020, 14, 595–608. [Google Scholar] [CrossRef] [Green Version]
- El-Sheshtawy, H.S.; Sofy, M.R.; Ghareeb, D.A.; Yacout, G.A.; Eldemellawy, M.A.; Ibrahim, B.M. Eco-friendly polyurethane acrylate (PUA)/natural filler-based composite as an antifouling product for marine coating. Appl. Microbiol. Biotechnol. 2021, 105, 7023–7034. [Google Scholar] [CrossRef]
- Bielke, L.; Higgins, S.; Donoghue, A.; Hargis, B.M. Salmonella Host Range of Bacteriophages That Infect Multiple Genera. Poult. Sci. 2007, 86, 2536–2540. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.P.; Oliveira, H.; Melo, L.; Sillankorva, S.; Azeredo, J. Bacteriophage-encoded depolymerases: Their diversity and biotechnological applications. Appl. Microbiol. Biotechnol. 2016, 100, 2141–2151. [Google Scholar] [CrossRef] [Green Version]
- O’Flynn, G.; Coffey, A.; Fitzgerald, G.; Ross, R. The newly isolated lytic bacteriophages st104a and st104b are highly virulent against Salmonella enterica. J. Appl. Microbiol. 2006, 101, 251–259. [Google Scholar] [CrossRef]
- Sofy, A.R.; Mahfouze, S.A.; El-Enany, M.A. Isozyme markers for response of wild potato species to potato spindle tuber viroid egyptian isolate. World Appl. Sci. J. 2013, 27, 1010–1022. [Google Scholar]
- Ahmadi, H.; Radford, D.; Kropinski, A.M.; Lim, L.-T.; Balamurugan, S. Thermal-Stability and Reconstitution Ability of Listeria Phages P100 and A511. Front. Microbiol. 2017, 8, 2375. [Google Scholar] [CrossRef] [PubMed]
- Sommer, J.; Trautner, C.; Witte, A.K.; Fister, S.; Schoder, D.; Rossmanith, P.; Mester, P.-J. Don’t Shut the Stable Door after the Phage Has Bolted—The Importance of Bacteriophage Inactivation in Food Environments. Viruses 2019, 11, 468. [Google Scholar] [CrossRef] [Green Version]
- Jepson, C.D.; March, J.B. Bacteriophage lambda is a highly stable DNA vaccine delivery vehicle. Vaccine 2004, 22, 2413–2419. [Google Scholar] [CrossRef] [PubMed]
- Magin, V.; Garrec, N.; Andrés, Y. Selection of Bacteriophages to Control In Vitro 24 h Old Biofilm of Pseudomonas aeruginosa Isolated from Drinking and Thermal Water. Viruses 2019, 11, 749. [Google Scholar] [CrossRef] [Green Version]
- Sofy, A.; El-Dougdoug, N.; Refaey, E.; Dawoud, R.; Hmed, A. Characterization and Full Genome Sequence of Novel KPP-5 Lytic Phage against Klebsiella pneumoniae Responsible for Recalcitrant Infection. Biomedicines 2021, 9, 342. [Google Scholar] [CrossRef] [PubMed]
- Sofy, M.R.; Aboseidah, A.A.; Heneidak, S.A.; Ahmed, H.R. ACC deaminase containing endophytic bacteria ameliorate salt stress in Pisum sativum through reduced oxidative damage and induction of antioxidative defense systems. Environ. Sci. Pollut. Res. 2021, 28, 40971–40991. [Google Scholar] [CrossRef] [PubMed]
- Dorman, H.J.D.; Deans, S.G. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef]
- Landers, T.F.; Cohen, B.; Wittum, T.E.; Larson, E.L. A Review of Antibiotic Use in Food Animals: Perspective, Policy, and Potential. Public Health Rep. 2012, 127, 4–22. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.-K.; Paik, H.-D. Status, Antimicrobial Mechanism, and Regulation of Natural Preservatives in Livestock Food Systems. Food Sci. Anim. Resour. 2016, 36, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Juhaimi, F.A.L.; Ghafoor, K.; Özcan, M.M.; Jahurul, M.H.A.; Babiker, E.E.; Jinap, S.; Sahena, F.; Sharifudin, M.S.; Zaidul, I.S.M. Effect of various food processing and handling methods on preservation of natural antioxidants in fruits and vegetables. J. Food Sci. Technol. 2018, 55, 3872–3880. [Google Scholar] [CrossRef] [PubMed]
- Harper, D.R.; Parracho, H.M.R.T.; Walker, J.; Sharp, R.J.; Hughes, G.; Werthén, M.; Lehman, S.M.; Morales, S. Bacteriophages and Biofilms. Antibiotics 2014, 3, 270–284. [Google Scholar] [CrossRef]
- Pennone, V.; Sanz-Gaitero, M.; O’Connor, P.; Coffey, A.; Jordan, K.; Van Raaij, M.J.; McAuliffe, O.; Gaitero, S.-; Connor, O.; Raaij, V. Inhibition of L. monocytogenes Biofilm Formation by the Amidase Domain of the Phage vB_LmoS_293 Endolysin. Viruses 2019, 11, 722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Virk, S.M.; Shi, J.; Zhou, Y.; Willias, S.P.; Morsy, M.K.; Abdelnabby, H.E.; Liu, J.; Wang, X.; Li, J. Isolation, Characterization, and Application of Bacteriophage LPSE1 against Salmonella enterica in Ready to Eat (RTE) Foods. Front. Microbiol. 2018, 9, 1046. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, L.D.; Bisha, B. Phage-based biocontrol strategies to reduce foodborne pathogens in foods. Bacteriophage 2011, 1, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Grant, A.; Hashem, F.; Parveen, S. Salmonella and Campylobacter: Antimicrobial resistance and bacteriophage control in poultry. Food Microbiol. 2016, 53, 104–109. [Google Scholar] [CrossRef]
- Osvaldo, L.O.P.C.; Nohelia, C.D.C.; Karina, R.; Bal, C.C.O.; López-Cuevas, O.; Campo, N.C.-D.; Ramírez, K.; Chaidez, C. Biocontrol of Salmonella Typhimurium growth in tomato surface by bacteriophage P22. Afr. J. Microbiol. Res. 2016, 10, 528–534. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serovars | Isolate ID | Source | Antibiotic-Resistance Profile | Optical Density of Cell Growth (OD620 nm) | Optical Density of Biofilm (OD570 nm) | Interpretation |
|---|---|---|---|---|---|---|
| Salmonella Typhimurium | ST-4 | Chicken (Egypt) | Resistant | 1.847 | 0.947 | Strong |
| Salmonella Typhimurium | ST-7 | Chicken (Egypt) | Resistant | 0.998 | 0.411 | Moderate |
| Salmonella Typhimurium | ST-9 | Chicken (Egypt) | Resistant | 0.876 | 0.563 | Moderate |
| Salmonella Typhimurium | ST-14 | Bovine (Egypt) | Resistant | 1.022 | 0.744 | Strong |
| Salmonella Typhimurium | ST-16 | Bovine (Egypt) | Resistant | 0.773 | 0.117 | Weak |
| Salmonella Typhimurium | ST-19 | Bovine (Egypt) | Resistant | 1.977 | 1.081 | Strong |
| Salmonella Typhimurium | ST-21 | Dairy products (Egypt) | Resistant | 0.712 | 0.108 | Weak |
| Salmonella Typhimurium | ST-22 | Dairy products (Egypt) | Resistant | 1.446 | 0.742 | Strong |
| Salmonella Typhimurium | ST-27 | Dairy products (Egypt) | Resistant | 1.784 | 0.748 | Strong |
| Salmonella Typhimurium | ST-30 | Chicken (Egypt) | Resistant | 1.709 | 0.904 | Strong |
| Salmonella Typhimurium | ST-33 | Chicken (Egypt) | Resistant | 0.993 | 0.321 | Moderate |
| Salmonella Typhimurium | ST-35 | Chicken (Egypt | Resistant | 1.003 | 0.411 | Moderate |
| Salmonella Typhimurium | ST-36 | Chicken (Egypt) | Resistant | 0.878 | 0.103 | Weak |
| Salmonella Typhimurium | ST-37 | Chicken (Egypt) | Resistant | 2.047 | 1.105 | Strong |
| Salmonella Typhimurium | ST-38 | Chicken (Egypt) | Resistant | 0.788 | 0.119 | Weak |
| Salmonella Typhimurium | ST-41 | Bovine (Egypt) | Resistant | 0.936 | 0.366 | Moderate |
| Salmonella Typhimurium | ST-43 | Bovine (Egypt) | Resistant | 1.171 | 0.773 | Strong |
| Salmonella Typhimurium | ST-45 | Bovine (Egypt) | Resistant | 1.664 | 0.817 | Strong |
| Salmonella Typhimurium | ST-47 | Bovine (Egypt) | Resistant | 0.899 | 0.541 | Moderate |
| Salmonella Typhimurium | ST-49 | Bovine (Egypt) | Resistant | 1.811 | 0.927 | Strong |
| Salmonella Typhimurium | ST-53 | Dairy products (Egypt) | Resistant | 0.659 | 0.173 | Weak |
| Salmonella Typhimurium | ST-55 | Dairy products (Egypt) | Resistant | 0.744 | 0.196 | Weak |
| Salmonella Typhimurium | ST-56 | Dairy products (Egypt) | Resistant | 1.022 | 0.511 | Moderate |
| Bacteria | Phages | Bacteria | Phages | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| vB_STS-1 | vB_STM-2 | vB_STS-3 | vB_STS-1 | vB_STM-2 | vB_STS-3 | ||||||||
| ST | EOP | ST | EOP | ST | EOP | ST | EOP | ST | EOP | ST | EOP | ||
| Salmonella Typhimurium (ST-4) Host | + | M | + | H | + | H | Salmonella Typhimurium ST-47 | + | L | + | H | + | L |
| Salmonella Typhimurium (ST-7) | + | H | + | H | + | M | Salmonella Typhimurium ST-49 | − | N | + | M | − | N |
| Salmonella Typhimurium (ST-9) | − | N | + | M | − | N | Salmonella Typhimurium (ST-53) | + | H | + | H | + | M |
| Salmonella Typhimurium (ST-14) | − | N | + | H | − | N | Salmonella Typhimurium (ST-55) | − | N | + | M | − | N |
| Salmonella Typhimurium (ST-16) | + | H | + | H | + | M | Salmonella Typhimurium (ST-56) | + | H | + | H | + | M |
| Salmonella Typhimurium (ST-19) | + | M | + | H | + | M | P. aeruginosa (PsaCI-1) | − | N | + | M | − | N |
| Salmonella Typhimurium (ST-21) | − | N | + | M | + | M | P. aeruginosa (PsaCI-2) | − | N | + | M | − | N |
| Salmonella Typhimurium (ST-22) | − | N | + | H | − | N | P. aeruginosa (PsaFI-1) | − | N | + | H | − | N |
| Salmonella Typhimurium (ST-27) | + | M | + | H | + | M | P. aeruginosa (PsaFI-2) | − | N | + | H | − | N |
| Salmonella Typhimurium (ST-30) | + | H | + | H | − | N | P. aeruginosa (PsaFI-1) | − | N | + | H | − | N |
| Salmonella Typhimurium (ST-33) | − | N | + | M | − | N | S. aureus (SaFI-1) | − | N | + | L | − | N |
| Salmonella Typhimurium (ST-35) | + | H | + | H | + | H | S. aureus (SaFI-2) | − | N | + | M | − | N |
| Salmonella Typhimurium (ST-36) | − | N | + | M | − | N | S. aureus (SaFI-3) | − | N | + | L | − | N |
| Salmonella Typhimurium (ST-37) | + | M | + | H | − | N | E. coli (EcCI-1) | − | N | + | M | − | N |
| Salmonella Typhimurium (ST-38) | + | M | + | H | + | M | E. coli (EcCI-2) | − | N | + | M | − | N |
| Salmonella Typhimurium (ST-41) | + | L | + | H | − | N | E. coli (EcCI-3) | − | N | + | M | − | N |
| Salmonella Typhimurium (ST-43) | + | L | + | H | − | N | E. coli (EcCI-4) | − | N | + | M | − | N |
| Salmonella Typhimurium (ST-45) | + | M | + | H | + | L | K. pneumoniae (KpCI-1) | − | N | − | N | − | N |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelhadi, I.M.A.; Sofy, A.R.; Hmed, A.A.; Refaey, E.E.; Soweha, H.E.; Abbas, M.A. Discovery of Polyvalent Myovirus (vB_STM-2) Phage as a Natural Antimicrobial System to Lysis and Biofilm Removal of Salmonella Typhimurium Isolates from Various Food Sources. Sustainability 2021, 13, 11602. https://doi.org/10.3390/su132111602
Abdelhadi IMA, Sofy AR, Hmed AA, Refaey EE, Soweha HE, Abbas MA. Discovery of Polyvalent Myovirus (vB_STM-2) Phage as a Natural Antimicrobial System to Lysis and Biofilm Removal of Salmonella Typhimurium Isolates from Various Food Sources. Sustainability. 2021; 13(21):11602. https://doi.org/10.3390/su132111602
Chicago/Turabian StyleAbdelhadi, Iman M. A., Ahmed R. Sofy, Ahmed A. Hmed, Ehab E. Refaey, Hany E. Soweha, and Mohamed A. Abbas. 2021. "Discovery of Polyvalent Myovirus (vB_STM-2) Phage as a Natural Antimicrobial System to Lysis and Biofilm Removal of Salmonella Typhimurium Isolates from Various Food Sources" Sustainability 13, no. 21: 11602. https://doi.org/10.3390/su132111602
APA StyleAbdelhadi, I. M. A., Sofy, A. R., Hmed, A. A., Refaey, E. E., Soweha, H. E., & Abbas, M. A. (2021). Discovery of Polyvalent Myovirus (vB_STM-2) Phage as a Natural Antimicrobial System to Lysis and Biofilm Removal of Salmonella Typhimurium Isolates from Various Food Sources. Sustainability, 13(21), 11602. https://doi.org/10.3390/su132111602