
Sharing the Agrarian Knowledge with Archaeology: First Evidence of the Dimorphism of Vitis Pollen from the Middle Bronze Age of N Italy (Terramara Santa Rosa di Poviglio)

 ,
, 
 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
1.1. Pollen Dimorphism
1.2. The Botanical and Agrarian Knowledge
1.3. A Key Plant for Archaeology
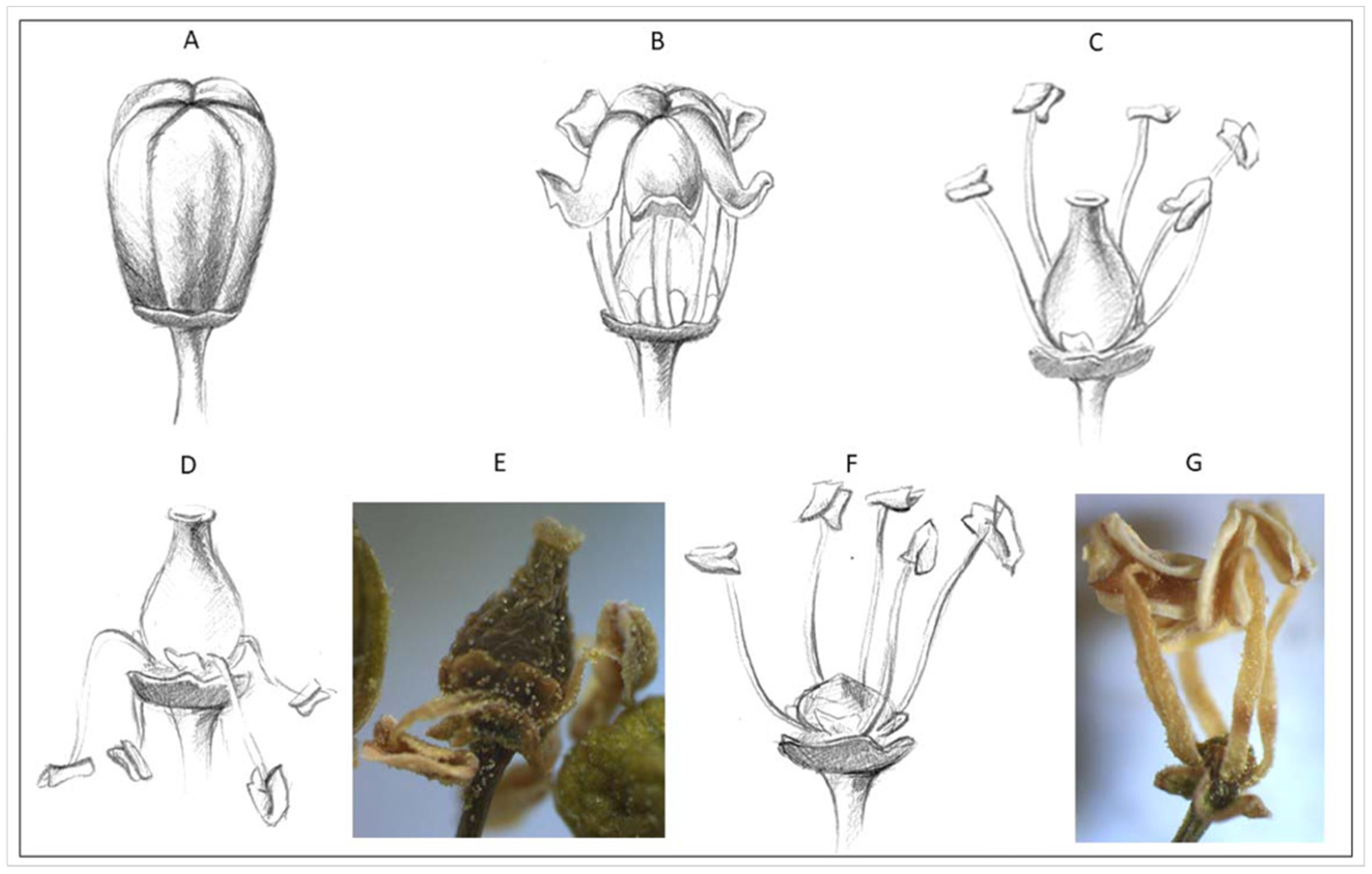
2. Materials and Methods
2.1. Reference Pollen Samples
2.1.1. V. vinifera Subsp. vinifera
2.1.2. V. vinifera Subsp. sylvestris
2.1.3. Acetolysis and Morphometry
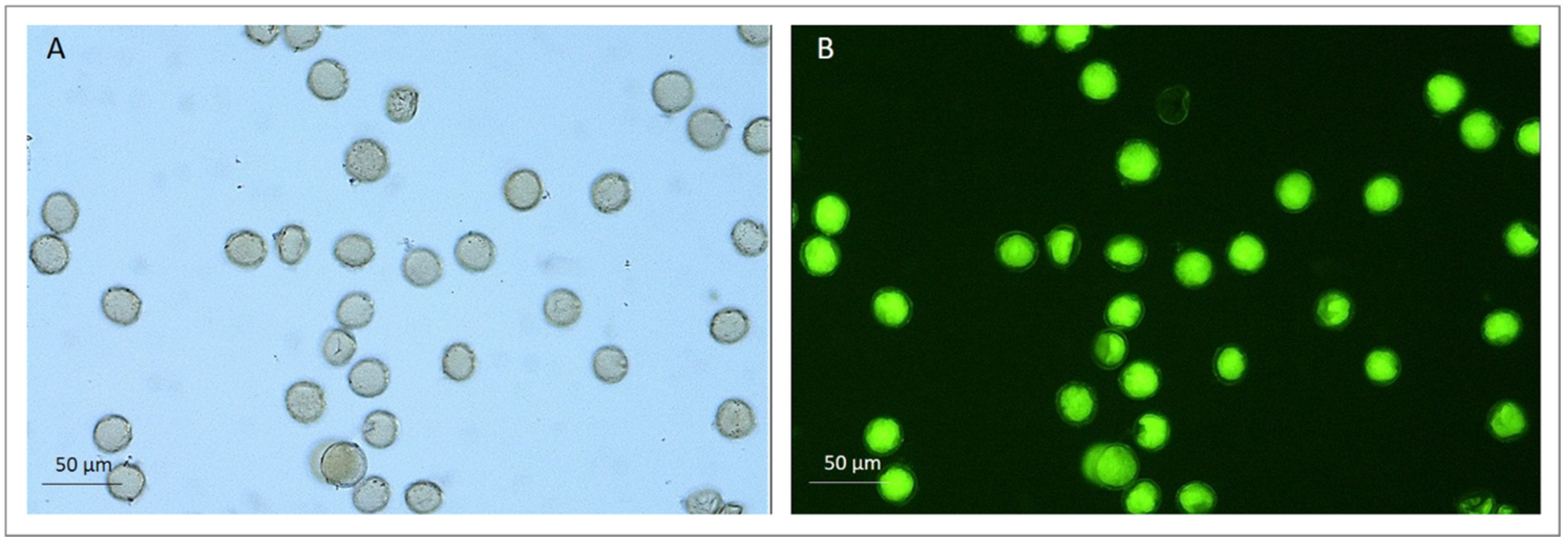
2.2. Pollen Viability and Germination
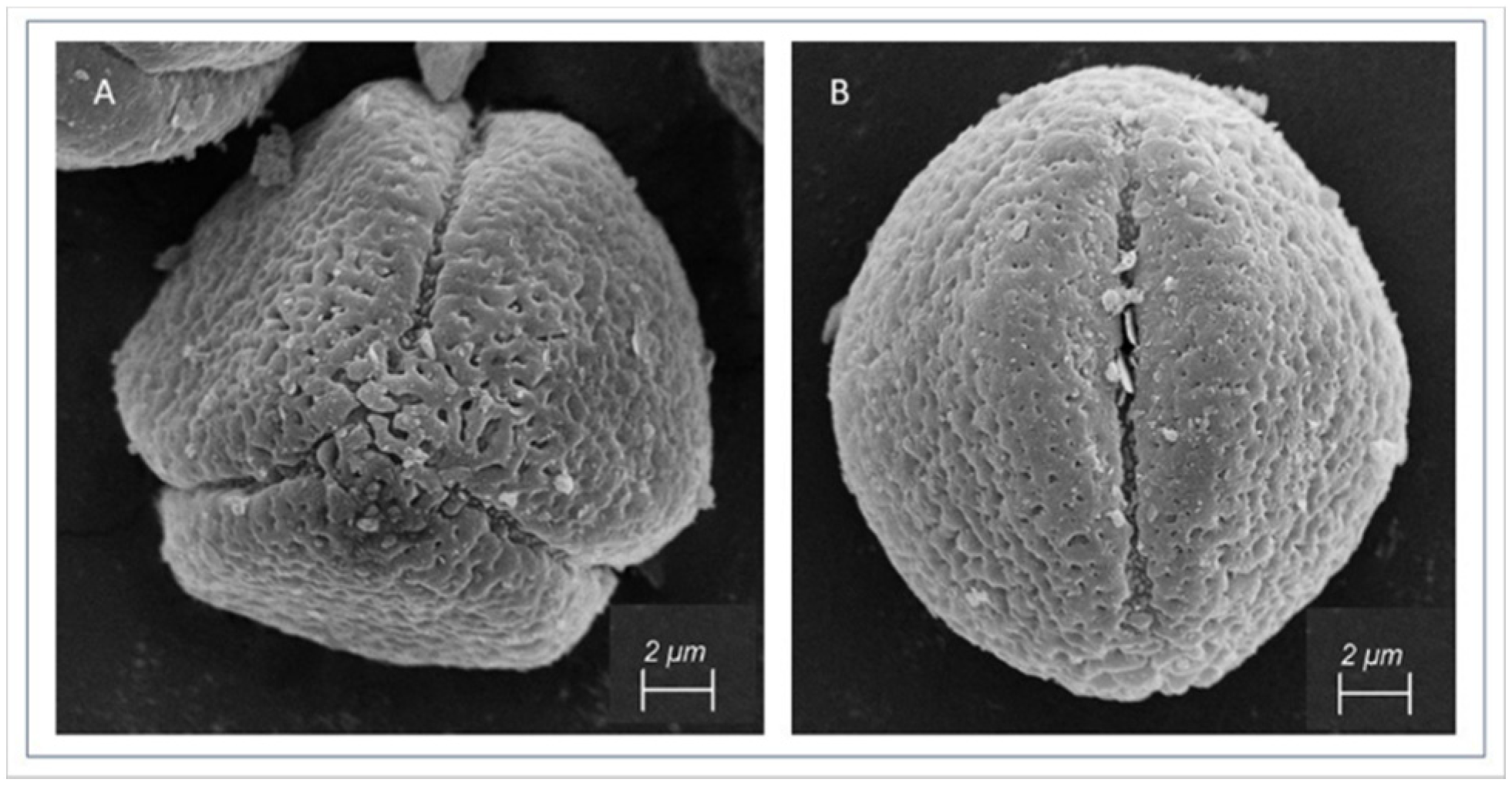
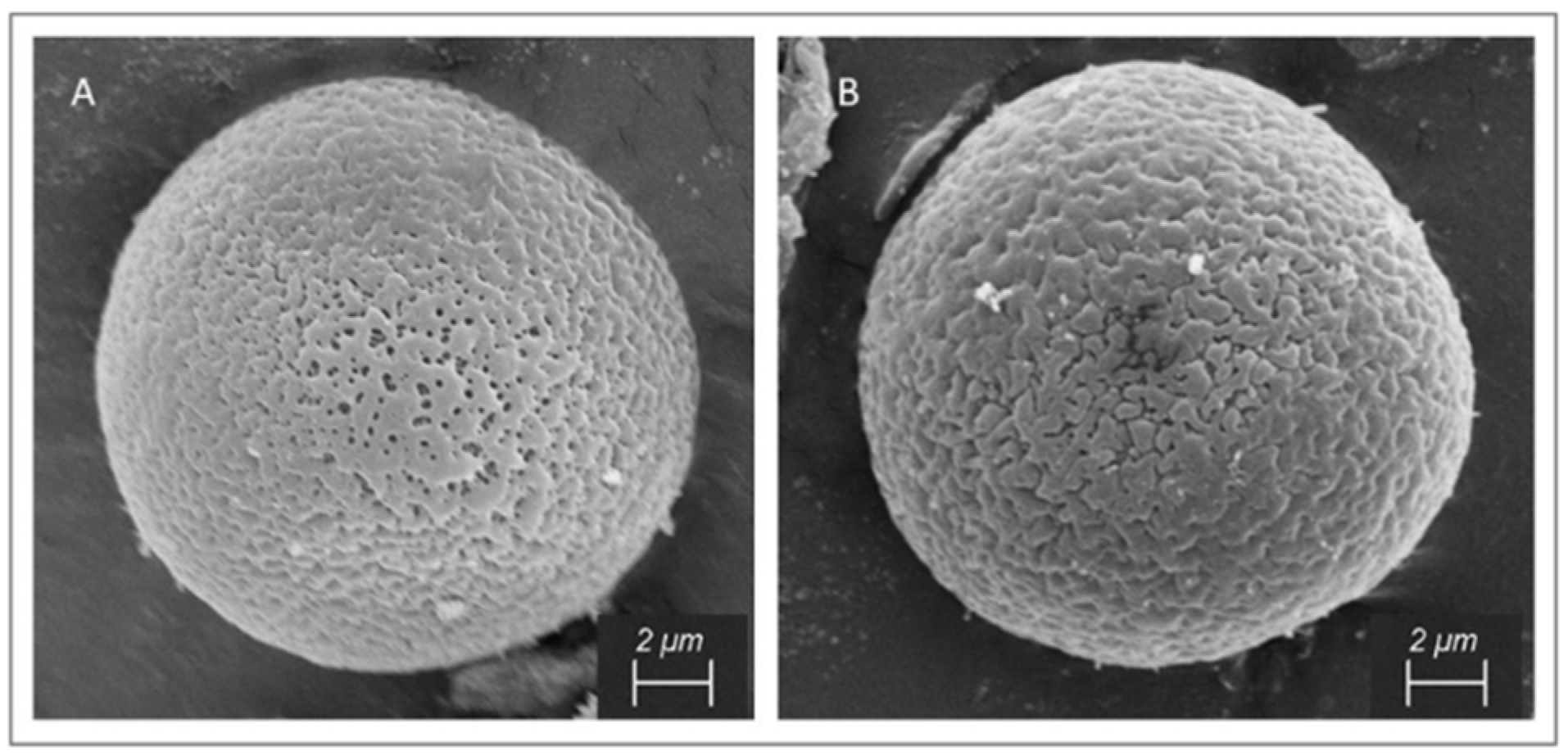
2.3. Scanning Electron Microscopy
2.4. The Archaeological Record for Pollen Analyses
3. Results
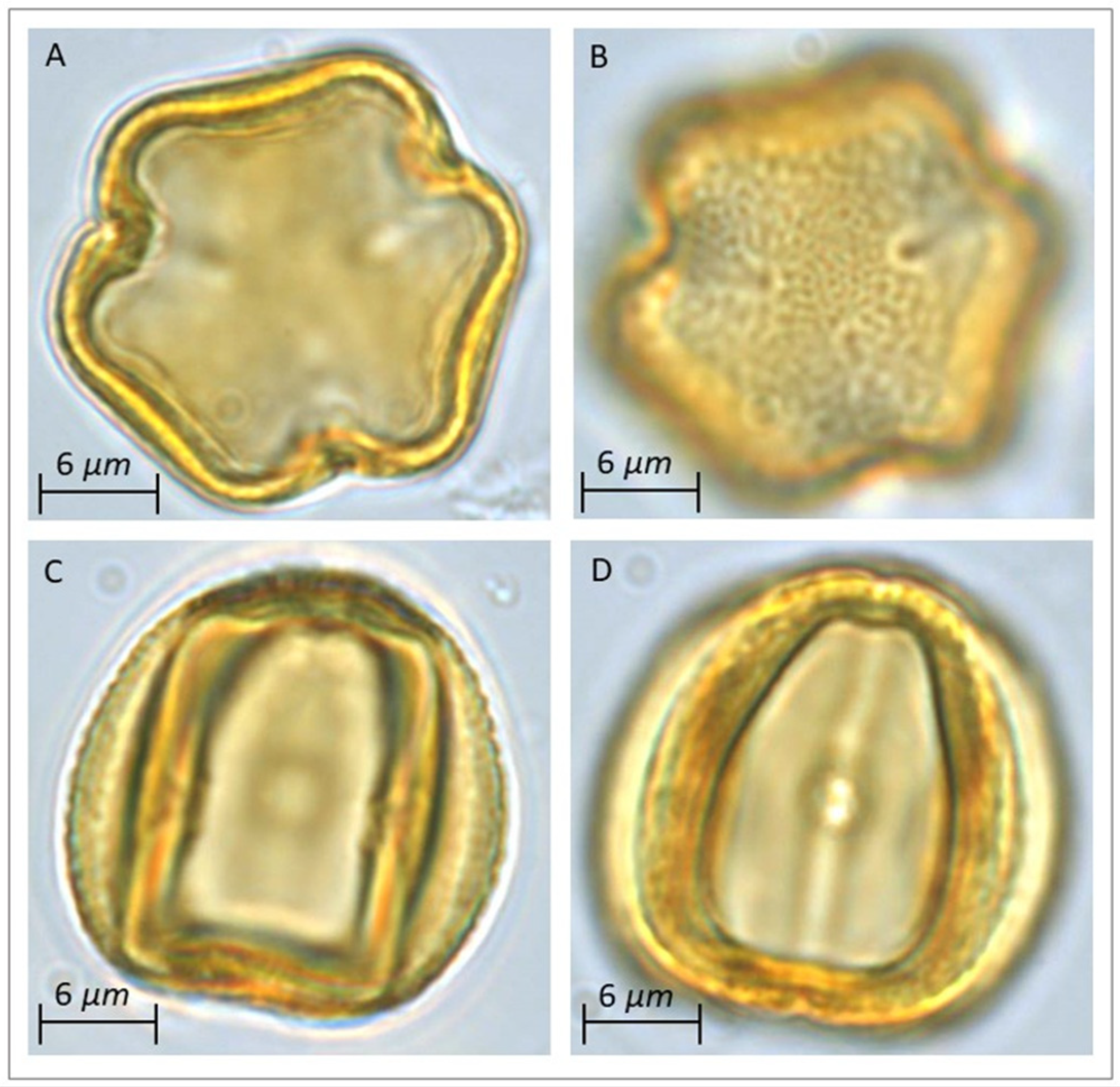
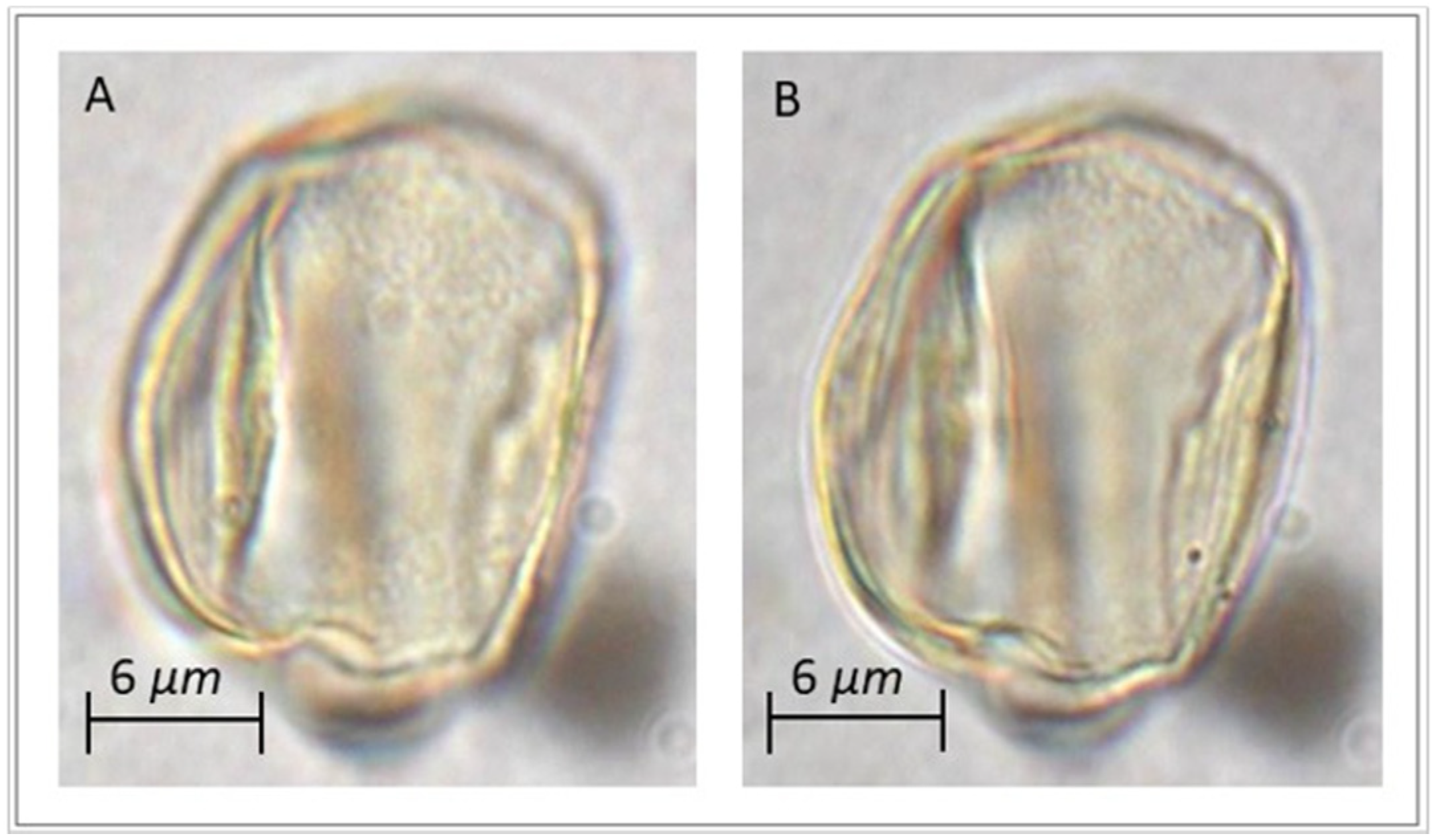
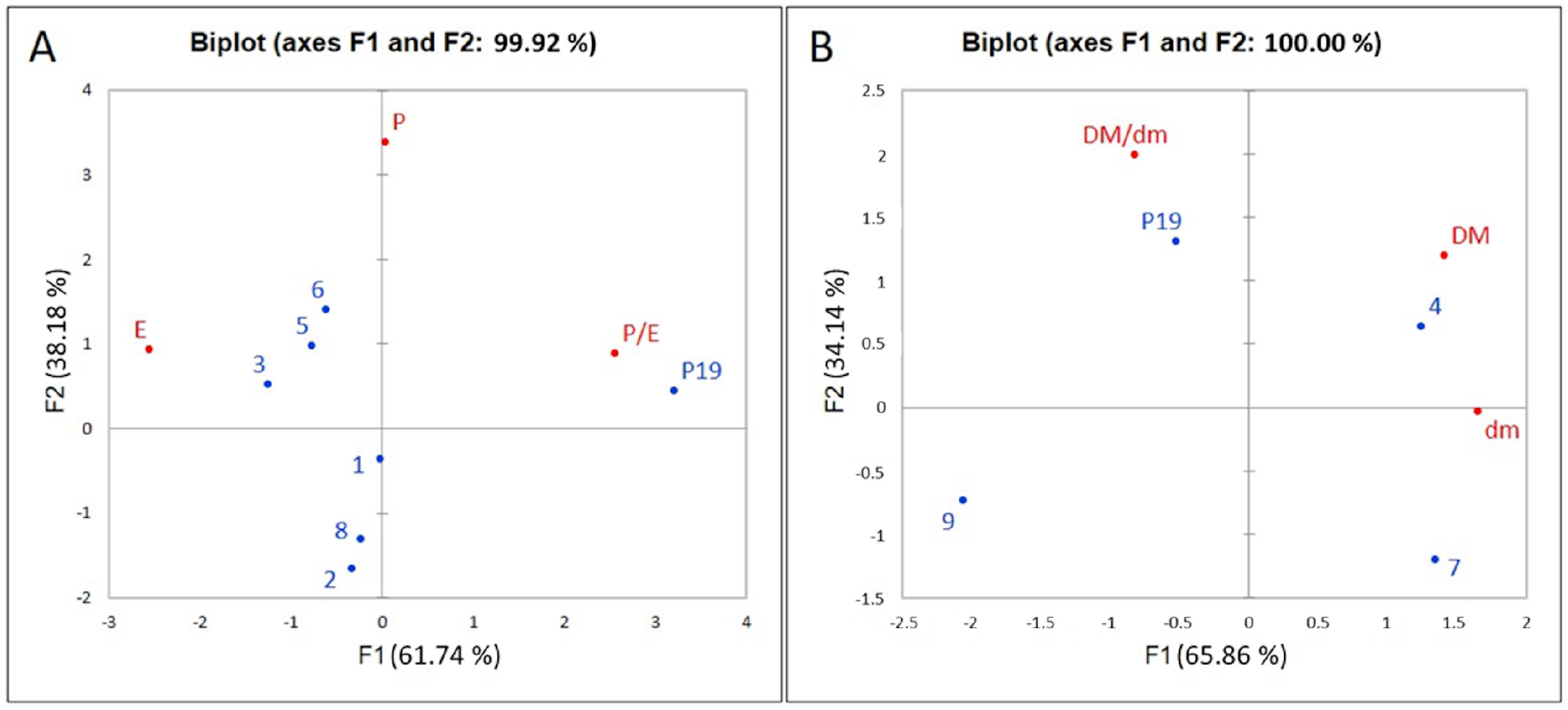
3.1. Pollen Morphology of Reference Pollen Samples
3.2. Pollen Viability
3.3. Pollen from Archaeological Samples
4. Discussion
4.1. Pollen Dimorphism of Current Vitis vinifera
4.1.1. Trizonocolporate Pollen
4.1.2. Inaperturate Pollen
4.2. The Current “Ancient Cultivars”
4.3. Viticulture during the Bronze Age in the Po Plain
4.4. Inaperturate Pollen as Evidence of Wild Plants in Past Records
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Horner, M.; Street, H.E. Pollen dimorphism—Origin and significance in pollen plant formation by another culture. Ann. Bot. 1978, 42, 763–777. [Google Scholar] [CrossRef]
- Kaltchuk-Santos, E.; Bodanese Zanettini, M.E.; Mundstock, E. Pollen dimorphism in soybean. Protoplasma 1993, 174, 74–78. [Google Scholar] [CrossRef]
- Kim, S.C.; Graham, S.A.; Graham, A. Palynology and pollen dimorphism in the genus Lagerstroemia (Lythraceae). Grana 1994, 33, 1–20. [Google Scholar] [CrossRef]
- Baker, H.G. The evolution, functioning and breakdown of heteromorphic incompatibility systems. I. Plumbaginaceae. Evolution 1966, 20, 349–368. [Google Scholar] [CrossRef]
- Barrett, S.C.H. Heterostyly in a tropical weed: The reproductive biology of the Turnera ulmifolia complex (Turneraceae). Can. J. Bot. 1978, 56, 1713–1725. [Google Scholar] [CrossRef]
- Haber, W.A.; Bawa, K.S. Evolution of dioecy in Saurauia (Dilleniaceae). Ann. Mo. Bot. Gard. 1984, 71, 289–293. [Google Scholar] [CrossRef]
- Anderson, G.J.; Symon, D.E. Functional dioecy and andromonoecy in Solanum. Evolution 1989, 43, 204–219. [Google Scholar] [CrossRef]
- Furness, C.A. Why does some pollen lack apertures? A review of inaperturate pollen in eudicots. Bot. J. Lin. Soc. 2007, 155, 29–48. [Google Scholar] [CrossRef] [Green Version]
- Penny, R.H.; Steven, J.C. Sexual dimorphism in pollen grain size in cryptically dioecious Thalictrum macrostylum. Plant Syst. Evol. 2009, 279, 11–19. [Google Scholar] [CrossRef]
- McNeill, J.; Crompton, C.W. Pollen dimorphism in Silene alba (Caryophyllaceae). Can. J. Bot. 1978, 56, 1280–1286. [Google Scholar] [CrossRef]
- Kevan, P.G.; Longair, R.W.; Gadawski, R.M. Dioecy and pollen dimorphism in Vitis riparia (Vitaceae). Can. J. Bot. 1985, 63, 2263–2267. [Google Scholar] [CrossRef]
- Kevan, P.G.; Blades, D.C.A.; Posluszny, U.; Ambrose, J.D. Pollen dimorphism and dioecy in Vitis aestivalis. Vitis 1988, 27, 143–146. [Google Scholar]
- Kimura, P.H.; Okamoto, G.; Hirano, K. The mode of pollination and stigma receptivity in Vitis coignetiae Pulliat. Am. J. Enol. Vitic. 1998, 49, 1–5. [Google Scholar]
- Oberle, G.D. A genetic study of variations in floral morphology and function in cultivated forms of Vitis. N. Y. State Agric. Exp. Stn. Bull. 1938, 250, 3–32. [Google Scholar]
- Pratt, C. Reproductive anatomy in cultivated grapes—A review. Am. J. Enol. Vitic. 1971, 22, 92–109. [Google Scholar]
- Caporali, E.; Spada, A.; Marziani, G.; Failla, O.; Scienza, A. The arrest of development of abortive reproductive organs in the unisexual flower of Vitis vinifera ssp. silvestris. Sex. Plant Reprod. 2003, 15, 291–300. [Google Scholar] [CrossRef]
- Vasconcelos, M.C.; Greven, M.; Winefield, C.S.; Trought, M.C.T.; Raw, V. The Flowering Process of Vitis vinifera: A review. Am. J. Enol. Vitic. 2009, 60, 411–434. [Google Scholar]
- Meneghetti, S.; Calò, A.; Costacurta, A.; Frare, E.; Crespan, M. Valutazione della variabilità intra-varietale in vite ai fini dell’identificazione clonale. Riv. Vitic. Enol. 2010, 1–4, 93–103. [Google Scholar]
- Coito, J.L.; Silva, H.G.; Ramos, M.; Cunha, J.; Eiras-Dias, J.; Amâncio, S.; Costa, M.; Rocheta, M. Vitis flower types: From the wild to crop plants. PeerJ 2019, 7, e7879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lorenzis, G.; Chipashvili, R.; Failla, O.; Maghradze, D. Study of genetic variability in Vitis vinifera L. germplasm by high-throughput Vitis18kSNP array: The case of Georgian genetic resources. BMC Plant Biol. 2015, 15, 154. [Google Scholar] [CrossRef] [Green Version]
- Dellaporta, S.L.; Calderon-Urrea, A. Sex determination in flowering plants. Plant Cell 1993, 5, 1241–1251. [Google Scholar] [PubMed] [Green Version]
- Olmo, H.P. The origin and domestication of the vinifera grape. In The Origins and Ancient History of Wine, 1st ed.; McGovern, P.E., Fleming, S.J., Katz, S.H., Eds.; Gordon and Breach: New York, NY, USA, 1996; pp. 31–43. ISBN 9780203392836. [Google Scholar]
- Zohary, D.; Hopf, M. Domestication of Plants in the Old World, 3rd ed.; Oxford University Press: Oxford, UK, 2000; ISBN 0198503569. [Google Scholar]
- This, P.; Lacombe, T.; Thomas, M.R. Historical origins and genetic diversity of wine grapes. Trends Genet. 2006, 22, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Henry, I.M.; Akagi, T.; Tao, R.; Comai, L. One hundred ways to invent the sexes: Theoretical and observed paths to dioecy in plants. Annu. Rev. Plant Biol. 2018, 69, 553–575. [Google Scholar] [CrossRef] [Green Version]
- Massonnet, M.; Cochetel, N.; Minio, A.; Vondras, A.M.; Lin, J.; Muyle, A.; Garcia, J.F.; Zhou, Y.; Delledonne, M.; Riaz, S.; et al. The genetic basis of sex determination in grapes. Nat. Commun. 2020, 11, 2902. [Google Scholar] [CrossRef]
- Sgarbi, E.; Barbieri, C.; Olmi, L.; Imazio, S.; Bignami, C. Morfologia, anatomia e funzionalità dei fiori in vitigni emiliani. Quad. Vitic. Enol. Univ. Torino 2013, 32, 403–407. [Google Scholar]
- Meneghetti, S.; Gardiman, M.; Calò, A. Flower biology of grapevine. A review. Adv. Hortic. Sci. 2006, 20, 317–325. [Google Scholar]
- Barbieri, C.; Baroni, A.; Sgarbi, E.; Bignami, C.; Meglioraldi, S.; Storchi, M.; Sivestroni, O. Pollination and fruit set: Critical factors for the production of the grapevine cultivar ‘Malbo Gentile’. Acta Hortic. 2012, 932, 155–162. [Google Scholar] [CrossRef]
- Dorsey, M.J. Pollen Development in the Grape with Special Reference to Sterility; Bulletin 144; University of Minnesota, Agricultural Experiment Station, University Farm: St. Paul, MN, USA, 1914; pp. 1–69. [Google Scholar]
- Cargnello, G.; Carraro, L.; Lombardo, G.; Gerola, F.M. Pollen morphology of Picolit grown in different Italian regions. Vitis 1980, 19, 201–206. [Google Scholar]
- Abreu, I.; Costa, I.; Oliveira, M.; Cunha, M.; De Castro, R. Ultrastructure and germination of Vitis vinifera cv. Loureiro pollen. Protoplasma 2006, 228, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Martínez, S.; Díaz, T.E.; Fernández-González, F.; Izco, J.; Lousã, M.; Penas, Á. Vascular plant communities of Spain and Portugal Addenda to the syntaxonomical checklist of 2001. Itinera Geobot. 2002, 15, 5–922. [Google Scholar]
- Italian Interpretation Manual of the Habitats (92/43/EEC Directive). Società Botanica Italiana, Ministero dell’Ambiente e della Tutela del Territorio e del Mare, D.P.N., 2009. Available online: http://vnr.unipg.it/habitat (accessed on 5 May 2020).
- Castiglioni, E.; Rottoli, M. Capua (Caserta). Località Strepparo e Cento Moggie. Scavi nell’area del Cira. Resti botanici da un pozzo dell’età del Bronzo. Boll. Archeol. 2001, 37–38, 62–67. [Google Scholar]
- Aranguren, B.; Bellini, C.; Mariotti Lippi, M.; Mori Secci, M.; Perazzi, P. L’avvio della coltura della vite in Toscana: L’esempio di San Lorenzo a Greve (FI). In Archeologia della Vite e del Vino in Etruria; Ciacci, A., Rendini, P., Zifferero, A., Eds.; Ci.Vin: Siena, Italy, 2007; pp. 88–97. [Google Scholar]
- Mariotti Lippi, M.; Bellini, C.; Mori Secci, M.; Gonnelli, T. Comparing seeds/fruits and pollen from a Middle Bronze Age pit in Florence (Italy). J. Archaeol. Sci. 2009, 36, 1135–1141. [Google Scholar] [CrossRef]
- Martinelli, M.C.; Fiorentino, G.; Prosdocimi, B.; D’Oronzo, C.; Levi, S.T.; Mangano, G.; Stellati, A.; Wolff, N. Nuove ricerche nell’insediamento sull’istmo di Filo Braccio a Filicudi. Nota preliminare sugli scavi 2009. Origini 2010, 32, 285–314. [Google Scholar]
- Buonincontri, M.P.; Pecci, A.; Di Pasquale, G.; Ricci, P.; Lubritto, C. Multiproxy approach to the study of Medieval food habits in Tuscany (central Italy). Archaeol. Anthropol. Sci. 2017, 9, 53–671. [Google Scholar] [CrossRef]
- Valamoti, S.M.; Pagnoux, C.; Ntinou, M.; Bouby, L.; Bonhomme, V.; Terral, J.F. More than meets the eye: New archaeobotanical evidence on Bronze Age viticulture and wine making in the Peloponnese, Greece. Veg. Hist. Archaeobot. 2020, 29, 35–50. [Google Scholar] [CrossRef]
- Pagnoux, C.; Bouby, L.; Ivorra, S.; Petit, C.; Valamoti, S.M.; Pastor, T.; Picq, S.; Terral, J.F. Inferring the agrobiodiversity of Vitis vinifera L. (grapevine) in ancient Greece by comparative shape analysis of archaeological and modern seeds. Veg. Hist. Archaeobot. 2015, 24, 75–84. [Google Scholar] [CrossRef]
- McGovern, P.; Jalabadze, M.; Batiuk, S.; Callahan, M.P.; Smith, K.E.; Hall, G.R.; Kvavadze, E.; Maghradze, D.; Rusishvili, N.; Bouby, L.; et al. Early Neolithic wine of Georgia in the South Caucasus. Proc. Natl. Acad. Sci. USA 2017, 114, E10309–E10318. [Google Scholar] [CrossRef] [Green Version]
- Zohary, D. The domestication of the grapevine Vitis vinifera L. in the near east. In The Origins and Ancient History of Wine; McGovern, P.E., Fleming, S.J., Katz, S.H., Eds.; Gordon and Breach: Amsterdam, The Netherlands, 1995; pp. 23–30. ISBN 9782881245770. [Google Scholar]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World; Oxford University Press: Oxford, UK, 2012; ISBN 9780199549061. [Google Scholar]
- Bakels, C. Plant remains from Sardinia, Italy, with notes on barley and grape. Veg. Hist. Archaeobot. 2002, 11, 3–8. [Google Scholar] [CrossRef]
- Terral, J.F.; Tabard, E.; Bouby, L.; Ivorra, S.; Pastor, T.; Figueiral, I.; Picq, S.; Chevance, J.B.; Jung, C.; Fabre, L.; et al. Evolution and history of grapevine (Vitis vinifera) under domestication: New morphometric perspectives to understand seed domestication syndrome and reveal origins of ancient European cultivars. Ann. Bot. 2010, 105, 443–455. [Google Scholar] [CrossRef]
- Ucchesu, M.; Orrù, M.; Grillo, O.; Venora, G.; Usai, A.; Serreli, P.F.; Bacchetta, G. Earliest evidence of a primitive cultivar of Vitis vinifera L. during the Bronze Age in Sardinia (Italy). Veg. Hist. Archaeobot. 2015, 24, 587–600. [Google Scholar] [CrossRef]
- Wales, N.; Ramos Madrigal, J.; Cappellini, E.; Carmona Baez, A.; Samaniego Castruita, J.A.; Romero Navarro, J.A.; Carøe, C.; Avila Arcos, M.C.; Penaloza, F.; Moreno Mayar, J.V.; et al. The limits and potential of paleogenomic techniques for reconstructing grapevine domestication. J. Archaeol. Sci. 2016, 72, 57–70. [Google Scholar] [CrossRef]
- Aversano, R.; Basile, B.; Buonincontri, M.P.; Carucci, F.; Carputo, D.; Frusciante, L.; Di Pasquale, G. Dating the beginning of the Roman viticultural model in the Western Mediterranean: The case study of Chianti (Central Italy). PLoS ONE 2017, 12, e0186298. [Google Scholar] [CrossRef] [Green Version]
- Grasso, A.M.; Fiorentino, G. Waterlogged grape remains (Vitis vinifera ssp. vinifera L.) from a Byzantine well at Supersano, Southeast Italy: Remains of wine making? In Proceedings of the 15th Conference of the International Work Group for Palaeoethnobotany (IWGP 2010), Wilhelmshaven, Germany, 31 May—5 June 2010; Bittmann, F., Ed.; Terra Nostra-Schriften der GeoUnion Alfred-Wegener-Stiftung: Berlin, Germany, 2010; p. 132. ISSN 0946-8978. [Google Scholar]
- Mariotti Lippi, M.; Mori Secci, M.; Giachi, G.; Bouby, L.; Terral, J.F.; Castiglioni, E.; Cottini, M.; Rottoli, M.; de Grummond, N.T. Plant remains in an Etruscan-Roman well at Cetamura del Chianti, Italy. Archaeol. Anthropol. Sci. 2020, 12, 1–18. [Google Scholar] [CrossRef]
- Pecci, A.; Borgna, E.; Mileto, S.; Dalla Longa, E.; Bosi, G.; Florenzano, A.; Mercuri, A.M.; Corazza, S.; Marchesini, M.; Vidale, M. Wine consumption in Bronze Age Italy: Combining organic residue analysis, botanical data and ceramic variability. J. Archaeol. Sci. 2020, 123, 105256. [Google Scholar] [CrossRef]
- Bouby, L.; Figueiral, I.; Bouchette, A.; Rovira, N.; Ivorra, S.; Lacombe, T.; Pastor, T.; Picq, S.; Marinval, P.; Terral, J.F. Bioarchaeological Insights into the Process of Domestication of Grapevine (Vitis vinifera L.) during Roman Times in Southern France. PLoS ONE 2013, 8, e63195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ucchesu, M.; Orrù, M.; Grillo, O.; Venora, G.; Paglietti, G.; Ardu, A.; Bacchetta, G. Predictive method for correct identification of archaeological charred grape seeds: Support for advances in knowledge of grape domestication process. PLoS ONE 2016, 11, e0149814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montecchi, M.C.; Mercuri, A.M. When palynology meets classical archaeology: The Roman and medieval landscapes at the Villa del Casale di Piazza Armerina, UNESCO site in Sicily. Archaeol. Anthropol. Sci. 2018, 10, 743–757. [Google Scholar] [CrossRef]
- Turner, S.D.; Brown, A.G. Vitis pollen dispersal in and from organic vineyards I. Pollen trap and soil pollen data. Rev. Palaeobot. Palynol. 2004, 129, 117–132. [Google Scholar] [CrossRef]
- Bottema, S.; Woldring, H. Anthropogenic indicators in the pollen record of the Eastern Mediterranean. In Man’s Role in the Shaping of the Eastern Mediterranean Landscape; Bottema, S., Entjes-Nieborg, G., van Zeist, W., Eds.; A.A. Balkema: Rotterdam, The Netherlands, 1990; pp. 231–264. [Google Scholar]
- Cremaschi, M.; Mercuri, A.M.; Benatti, A.; Bosi, G.; Brandolini, F.; Clò, E.; Florenzano, A.; Furia, E.; Mariani, G.S.; Mazzanti, M.; et al. The SUCCESSO-TERRA Project: A Lesson of Sustainability from the Terramare Culture, Middle Bronze Age of the Po Plain (Northern Italy). Interdiscip. Archaeol. 2018, 2, 221–229. [Google Scholar] [CrossRef]
- Bernabò Brea, M.; Cardarelli, A. Le Terramare nel tempo. In Le Terramare. La Più Antica Civiltà Padana; Bernabò Brea, M., Cardarelli, A., Cremaschi, M., Eds.; Electa: Milano, Italy, 1997; pp. 295–301. ISBN 9788843560622. [Google Scholar]
- Cremaschi, M.; Mercuri, A.M.; Torri, P.; Florenzano, A.; Pizzi, C.; Marchesini, M.; Zerboni, A. Climate change versus land management in the Po Plain (Northern Italy) during the Bronze Age: New insights from the VP/VG sequence of the Terramara Santa Rosa di Poviglio. Quat. Sci. Rev. 2016, 136, 153–172. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Montecchi, M.C.; Pellacani, G.; Florenzano, A.; Rattighieri, E.; Cardarelli, A. Environment, human impact and the role of trees on the Po plain during the Middle and Recent Bronze Age: Pollen evidence from the local influence of the terramare of Baggiovara and Casinalbo. Rev. Pal. Pal. 2015, 218, 231–249. [Google Scholar] [CrossRef]
- Database Viticolo Italiano (VitisDB). Available online: https://vitisdb.it (accessed on 1 July 2020).
- The European Vitis Database. Available online: http://www.eu-vitis.de (accessed on 1 July 2020).
- Erdtmann, G. Handbook of Palynolgy: Morphology, Taxonomy, Ecology. An Introduction to the Study of Pollen Grains and Spores; Munksgaard: Copenhagen, Denmark, 1969. [Google Scholar]
- Wrońska-Pilarek, D.; Jagodziński, A.M.; Bocianowski, J.; Janyszek, M. The optimal sample size in pollen morphological studies using the example of Rosa canina L. (Rosaceae). Palynology 2015, 39, 56–75. [Google Scholar] [CrossRef]
- Messora, R.; Florenzano, A.; Torri, P.; Mercuri, A.M.; Muzzalupo, I.; Arru, L. Morphology and Discrimination Features of Pollen from Italian Olive Cultivars (Olea europaea L.). Grana 2017, 56, 204–214. [Google Scholar] [CrossRef]
- Heslop-Harrison, J.; Heslop-Harrison, Y. Evaluation of pollen viability by enzymatically induced fluorescence; intracellular hydrolysis of fluorescein diacetate. Stain Technol. 1970, 45, 115–120. [Google Scholar] [CrossRef]
- Shivanna, K.R.; Rangaswamy, N.S. Pollen Biology. A Laboratory Manual; Springer: Berlin, Germany, 1992; ISBN 9780387551708. [Google Scholar]
- Florenzano, A.; Mercuri, A.M.; Pederzoli, A.; Torri, P.; Bosi, G.; Olmi, L.; Rinaldi, R.; Bandini Mazzanti, M. The significance of intestinal parasite remains in pollen samples from Medieval pits in the Piazza Garibaldi of Parma, Emilia-Romagna, Northern Italy. Geoarchaeology 2012, 27, 34–47. [Google Scholar] [CrossRef]
- Erdtman, G. Pollen Morphology and Plant Taxonomy. Angiosperms I; Almqvist & Wiksell: Stockholm, Sweden, 1952. [Google Scholar]
- Punt, W.; Hoen, P.P.; Blackmore, S.; Nilsson, S.; Le Thomas, A. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol. 2007, 143, 1–81. [Google Scholar] [CrossRef]
- Lombardo, G.; Carraro, L.; Cargnello, G.; Bassi, M. Ultrastructure of pollen of Vitis vinifera L. cv. ‘Picolit giallo’ and its behaviour in experiments of self-and cross-pollination. Vitis 1976, 15, 73–81. [Google Scholar]
- Barbieri, C. Analisi degli Effetti di Biologia Fiorale e Cambiamenti Climatici su Qualità e Quantità della Produzione di Vitis vinifera L. nella Viti-Vinicoltura Reggiana. Ph.D. Thesis, University of Modena and Reggio Emilia, Reggio Emilia, Italy, 2012. [Google Scholar]
- Zohary, D. Unconscious selection and the evolution of domesticated plants. Econ. Bot. 2004, 58, 5–10. [Google Scholar] [CrossRef]
- Punt, W.; Marks, A.; Hoen, P.P. Vitaceae. In The Northwest European Pollen Flora VIII; Punt, W., Blackmore, S., Hoen, P.P., Stafford, P.J., Eds.; Elsevier: Amsterdam, The Netherland, 2003; pp. 67–70. ISBN 978-0444827579. [Google Scholar]
- Gallardo, A.; Ocete, R.; López, M.A.; Lara, M.; Rivera, D. Assessment of pollen dimorphism of Vitis vinifera L. subspecies sylvestris (Gmelin) Hegi in Spain. Vitis 2009, 48, 59–62. [Google Scholar]
- Beug, H.J. Leitfaden der Pollenbestimmung für Mitteleuropa und Angrenzende Gebiete, 2nd ed.; Verlag Dr. Friedrich Pfeil: München, Germany, 2015; ISBN 978-3899370430. [Google Scholar]
- Dobritsa, A.A.; Coerper, D. The novel plant protein Inaperturate Pollen 1 marks distinct cellular domains and controls formation of apertures in the Arabidopsis pollen exine. Plant Cell 2012, 24, 4452–4464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büyükkartal, H.N.; Çölgeçen, H.; Marasa, B. Development of anther wall throughout microsporogenesis in Vitis vinifera L. cv. Çavuş. Int. J. Agric. Biol. 2005, 7, 616–620. [Google Scholar]
- Ucchesu, M.; Penã-Chocarro, L.; Sabato, D.; Tanda, G. Bronze Age subsistence in Sardinia (Italy): Cultivated plants and wild resources. Veg. Hist. Archaeobot. 2014, 24, 343–355. [Google Scholar] [CrossRef]
- Maini, L. Catalogo Alfabetico di Quasi Tutte le Uve o Viti Conosciute e Coltivate Nelle Provincie di Modena e Reggio Secondo i Loro Nomi Volgari; Tipografia Moneti e Pelloni: Modena, Italy, 1851. [Google Scholar]
- Re, F. Manoscritto; Biblioteca Panizzi: Reggio Emilia, Italy, 1805. [Google Scholar]
- Labra, M.; Winfield, M.; Ghiani, A.; Grassi, G.; Sala, F.; Scienza, A.; Failla, O. Genetic studies on Trebbiano and morphologically related varieties by SSR and AFLP markers. Vitis 2001, 40, 187–190. [Google Scholar]
- Bignami, C.; Imazio, S.; Antonelli, A.; Masino, F.; Matrella, V.; Montevecchi, G.; Vasile Simone, G.; Olmi, L.; Sgarbi, E. Malbo gentile. Italian Vitis Database, 2015. Available online: www.vitisdb.it (accessed on 5 July 2020).
- Emanuelli, F.; Lorenzi, S.; Grzeskowiak, L.; Catalano, V.; Stefanini, M.; Troggio, M.; Myles, S.; Martinez-Zapater, J.M.; Zyprian, E.; Moreira, F.M.; et al. Genetic diversity and population structure assessed by SSR and SNP markers in a large germplasm collection of grape. BMC Plant Biol. 2013, 13, 39. [Google Scholar] [CrossRef] [Green Version]
- Popescu, C.F.; Dejeu, L.C.; Ocete, R.R. Preliminary characterization of wild grapevine populations (Vitis vinifera ssp. sylvestris) grown along the Danube river. Not. Bot. Horti Agrobot. Cluj Napoca 2013, 41, 472–477. [Google Scholar] [CrossRef] [Green Version]
- Maghradze, D.; Aslanishvili, A.; Mdinaradze, I.; Tkemaladze, D.; Mekhuzla, L.; Lordkipanidze, D.; Jalabadze, M.; Kvavadze, E.; Rusishvili, N.; McGovern, P.; et al. Progress for research of grape and wine culture in Georgia, the South Caucasus. BIO Web Conf. 2019, 12, 3003. [Google Scholar] [CrossRef]
- Arroyo-García, R.A.; Ruiz-García, L.; Bolling, L.; Ocete, R.; López, M.A.; Arnold, C.; Ergul, A.; Söylemezoğlu, G.; Uzun, H.I.; Cabello, F.; et al. Multiple origins of cultivated grapevine (Vitis vinifera L. ssp. sativa) based on chloroplast DNA polymorphisms. Mol. Ecol. 2006, 15, 3707–3714. [Google Scholar]
- Garnier, N.; Valamoti, S.M. Prehistoric wine-making at Dikili Tash (Northern Greece): Integrating residue analysis and Archaeobotany. J. Archaeol. Sci. 2016, 74, 195–206. [Google Scholar] [CrossRef]
- McGovern, P.E.; Zhang, J.; Tang, J.; Zhang, Z.; Hall, G.R.; Moreau, R.A.; Nuñez, A.; Butrym, E.D.; Richards, M.P.; Wang, C.-S.; et al. Fermented beverages of pre- and proto-historic China. Proc. Natl. Acad. Sci. USA 2004, 101, 17593–17598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercuri, A.M.; Fornaciari, R.; Gallinaro, M.; Vanin, S.; di Lernia, S. Plant behaviour from human imprints and the cultivation of wild cereals in Holocene Sahara. Nat. Plants 2018, 4, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Castelletti, L.; Castiglioni, E.; Rottoli, M. L’agricoltura dell’Italia settentrionale dal Neolitico al Medioevo. In Le Piante Coltivate e la Loro Storia. Dalle Origini al Transgenico in Lombardia nel Centenario della Riscoperta Genetica di Mendel; Failla, O., Forni, G., Eds.; Franco Angeli: Milano, Italy, 2001; pp. 33–84. ISBN 978-8846427212. [Google Scholar]
- Cardarelli, A.; Bosi, G.; Rinaldi, R.; Ucchesu, M.; Bacchetta, G. Vino o non vino. Nuovi dati sui vinaccioli della Terramara di Montale (Modena) tra la fine della media età del Bronzo e il Bronzo Recente. In Proceedings of the 50th Riunione Preistoria del Cibo. L’Alimentazione nella Preistoria e nella Protostoria, Rome, Italy, 5–8 October 2015; Available online: http://preistoriadelcibo.iipp.it/contributi/3_31.pdf (accessed on 7 July 2020).
- Menozzi, B.I.; Fichera, A.; Guido, M.A.; Mariotti Lippi, M.; Montanari, C.; Zanchetta, G.; Bonadonna, F.P.; Garbari, F. Lineamenti paleoambientali del bacino del Lago di Massaciuccoli (Toscana nord-occidentale). Atti Soc. Toscana Sci. Nat. Ser. B 2002, 109, 177–187. [Google Scholar]
- Cardarelli, A. La Necropoli della Terramara di Casinalbo; All’Insegna del Giglio: Firenze, Italy, 2014; ISBN 978-88-7814-537-5. [Google Scholar]
- Mercuri, A.M.; Accorsi, C.A.; Bandini Mazzanti, M.; Bosi, G.; Cardarelli, A.; Labate, D.; Trevisan Grandi, G. Economy and environment of Bronze Age settlements -Terramaras- in the Po Plain (Northern Italy): First results of the archaeobotanical research at the Terramara di Montale. Veg. Hist. Archaeobot. 2006, 16, 43–60. [Google Scholar] [CrossRef]
- Mariotti Lippi, M.; Di Tommaso, P.L.; Giachi, G.; Mori Secci, M.; Paci, S. Archaeobotanical investigations into an etruscan farmhouse at Pian d’Alma (Grosseto, Italy). Atti Soc. Toscana Sci. Nat. Ser. B 2002, 109, 159–165. [Google Scholar]
- Mercuri, A.M.; Bandini Mazzanti, M.; Florenzano, A.; Montecchi, M.C.; Rattighieri, E. Olea, Juglans and Castanea: The OJC group as pollen evidence of the development of human induced environments in the Italian peninsula. Quat. Int. 2013, 303, 24–42. [Google Scholar] [CrossRef]
- Berger, J.F.; Shennan, S.; Woodbridge, J.; Palmisano, A.; Mazier, F.; Nuninger, L.; Guillon, S.; Doyen, E.; Begeot, C.; ANdrieu-Ponel, V.; et al. Holocene land cover and population dynamics in Southern France. Holocene 2019, 29, 776–798. [Google Scholar] [CrossRef]
- Deza-Araujo, M.; Morales-Molino, C.; Tinner, W.; Henne, P.D.; Heitz, C.; Pezzatti, G.B.; Hafner, A.; Conedera, M. A critical assessment of human-impact indices based on anthropogenic pollen indicators. Quat. Sci. Rev. 2020, 236, 106291. [Google Scholar] [CrossRef]
- Rivera Nuñez, D.; Walker, M.J. A review of palaeobotanical findings of early Vitis in the Mediterranean and of the origins of cultivated grapevines, with special reference to new pointers to prehistoric exploitation in the western Mediterranean. Rev. Palaeobot. Palynol. 1989, 61, 205–237. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Species | Subsp. | Cultivar/ Label | Flower Morphology | Flower Functionality | Location | Date |
|---|---|---|---|---|---|---|---|
| 1 | Vitis vinifera | vinifera | Covra | Hermaphroditic | Male/female | Corlo di Formigine (MO) | May 2014 |
| 2 | Vitis vinifera | vinifera | Lambrusco Grasparossa | Hermaphroditic | Male/female | Corlo di Formigine (MO) | May 2014 |
| 3 | Vitis vinifera | vinifera | Bianca di Poviglio | Hermaphroditic | Male/female | Reggio Emilia (RE) | May 2016 |
| 4 | Vitis vinifera | vinifera | Malbo Gentile | Hermaphroditic | Female | Albinea (RE) | May/ June 2009 |
| 5 | Vitis vinifera | sylvestris | Accessione Canalone Sotto | Abortive pistil—fully developed stamens | Male | Torrazza Coste (PV) | May 2016 |
| 6 | Vitis vinifera | sylvestris | Accessione Ponte 5 | Abortive pistil—fully developed stamens | Male | Torrazza Coste (PV) | May 2016 |
| 7 | Vitis vinifera | sylvestris | Accessione Pioppetto 3 | Hermaphroditic | Female | Torrazza Coste (PV) | May 2016 |
| 8 | Vitis vinifera | sylvestris | Wild dioecious plants | Abortive pistil—fully developed stamens | Male | Sammontana (FI) | June 2019, 2020 |
| 9 | Vitis vinifera | sylvestris | Wild dioecious plants | Hermaphroditic | Female | Sammontana (FI) | June 2019 |
| Sample | Cultivar/ Label | No of Pollen Grains | Ex (µm) | DM (µm) | dm (µm) | DM/ dm | Pollen Shape (%) | Exine | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 5 | ||||||||
| 4 | Malbo Gentile | 30 | 1.2 ± 0.10 | 27.1 ± 1.70 | 25.1 ± 2.20 | 1.08 | - | 10 | 17 | 47 | 23 | 3 | 1 |
| 7 | Accessione Pioppetto 3 | 30 | 1.3 ± 0.13 | 25.4 ± 1.35 | 25.3 ± 1.12 | 1.00 | - | 37 | 10 | - | 53 | - | 0 |
| 9 | Wild dioecious plants | 45 | 1.2 ± 0.35 | 21.9 ± 0.9 | 20.3 ± 1.5 | 1.08 | - | - | 13 | 67 | 20 | - | 2 |
| Poviglio P19 | - | 15 | 1.1 ± 0.15 | 25.7 ± 4.6 | 22.5 ± 3.5 | 1.14 | - | - | 4 | 38 | 42 | 16 | 0 |
| Sample | Cultivar/ Label | No of Pollen Grains | Equatorial View | Polar View | Pollen Shape (%) | Exine | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P (µm) | E (µm) | P/E | MES (µm) | P- Porus (µm) | E- Porus (µm) | Ex (µm) | E1 (µm) | E2 (µm) | 0 | 1 | 2 | 3 | 4 | 5 | ||||
| 1 | Covra | 30 | 22.5 ± 1.34 | 20.7 ± 1.38 | 1.09 | 9.8 ± 1.06 | 1.7 ± 0.26 | 1.5 ± 0.19 | 0.9 ± 0.10 | 17.8 ± 1.14 | 20.4 ± 0.70 | - | 7 | 3 | 70 | 20 | - | 0 |
| 2 | Lambrusco Grasparossa | 30 | 20.7 ± 1.65 | 20.5 ± 1.62 | 1.01 | 10.3 ± 1.33 | 2.1 ± 0.33 | 1.7 ± 0.33 | 0.8 ± 0.10 | 20.9 ± 1.03 | 22.7 ± 0.93 | 3 | 47 | 6.5 | 37 | 6.5 | - | 0 |
| 3 | Bianca di Poviglio | 30 | 23.6 ± 2.10 | 23.6 ± 2.19 | 1.00 | 9.8 ± 1.80 | 1.8 ± 0.31 | 1.6 ± 0.25 | 0.9 ± 0.16 | 21.4 ± 2.06 | 24.7 ± 2.09 | 6 | 53 | - | 37 | 4 | - | 0 |
| 5 | Accessione Canalone Sotto | 30 | 24.3 ± 1.43 | 22.9 ± 1.10 | 1.06 | 12.5 ± 0.89 | 1.7 ± 0.29 | 1.5 ± 0.28 | 0.9 ± 0.10 | 18.3 ± 1.02 | 21.1 ± 1.05 | 6 | 14 | - | 74 | 6 | - | 0 |
| 6 | Accessione Ponte 5 | 30 | 24.9 ± 1.90 | 22.8 ± 1.22 | 1.09 | 10.4 ± 1.20 | 1.5 ± 0.23 | 1.4 ± 0.20 | 0.8 ± 0.08 | 20.4 ± 1.28 | 22.8 ± 1.31 | - | 3 | - | 83 | 14 | - | 0 |
| 8 | Wild dioecious plants | 82 | 21.2 ± 3.66 | 20.5 ± 4.20 | 1.03 | 13.2 ± 3.80 | 2.6 ± 1.60 | 2.1 ± 1.30 | 1.6 ± 0.70 | 18.8 ± 2.70 | 20.5 ± 3.40 | 1 | 11 | 4 | 62 | 21 | 1 | 0/1 |
| Poviglio P19 | - | 30 | 23.6 ± 3.90 | 15.8 ± 2.50 | 1.49 | - | - | - | 0.9 ± 0.12 | - | - | - | - | - | - | 6 | 94 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mercuri, A.M.; Torri, P.; Florenzano, A.; Clò, E.; Mariotti Lippi, M.; Sgarbi, E.; Bignami, C. Sharing the Agrarian Knowledge with Archaeology: First Evidence of the Dimorphism of Vitis Pollen from the Middle Bronze Age of N Italy (Terramara Santa Rosa di Poviglio). Sustainability 2021, 13, 2287. https://doi.org/10.3390/su13042287
Mercuri AM, Torri P, Florenzano A, Clò E, Mariotti Lippi M, Sgarbi E, Bignami C. Sharing the Agrarian Knowledge with Archaeology: First Evidence of the Dimorphism of Vitis Pollen from the Middle Bronze Age of N Italy (Terramara Santa Rosa di Poviglio). Sustainability. 2021; 13(4):2287. https://doi.org/10.3390/su13042287
Chicago/Turabian StyleMercuri, Anna Maria, Paola Torri, Assunta Florenzano, Eleonora Clò, Marta Mariotti Lippi, Elisabetta Sgarbi, and Cristina Bignami. 2021. "Sharing the Agrarian Knowledge with Archaeology: First Evidence of the Dimorphism of Vitis Pollen from the Middle Bronze Age of N Italy (Terramara Santa Rosa di Poviglio)" Sustainability 13, no. 4: 2287. https://doi.org/10.3390/su13042287
APA StyleMercuri, A. M., Torri, P., Florenzano, A., Clò, E., Mariotti Lippi, M., Sgarbi, E., & Bignami, C. (2021). Sharing the Agrarian Knowledge with Archaeology: First Evidence of the Dimorphism of Vitis Pollen from the Middle Bronze Age of N Italy (Terramara Santa Rosa di Poviglio). Sustainability, 13(4), 2287. https://doi.org/10.3390/su13042287