
Morphological and Genetic Characterization of Local Maize Accessions from Emilia Romagna Region, Italy

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Germplasm
2.2. DNA Extraction and PCR Amplification
2.3. Molecular Marker Data Analysis
3. Results and Discussion
Historical and Morphological Characterization of Germplasm
4. Genetic Characterization of Accessions
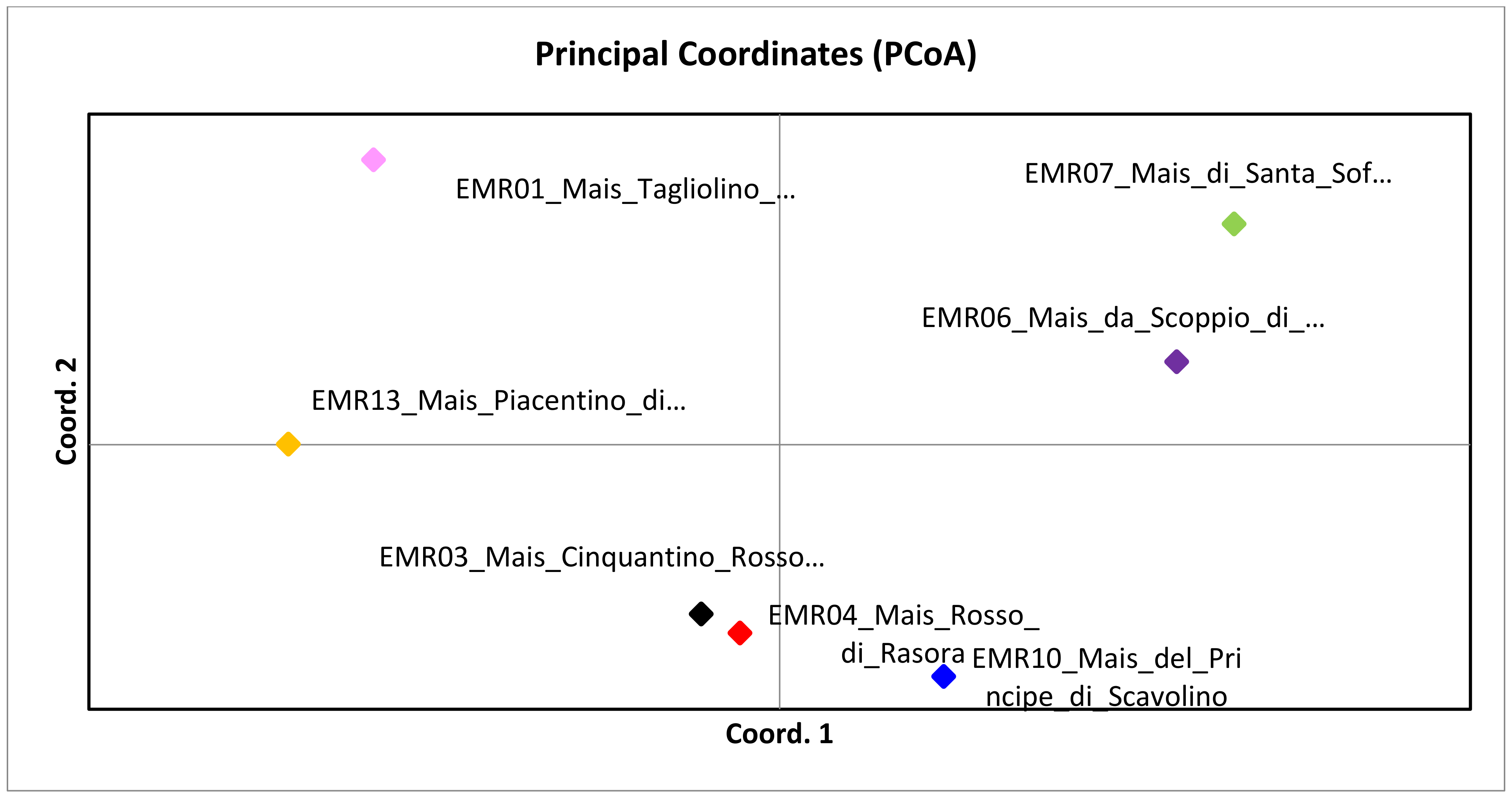
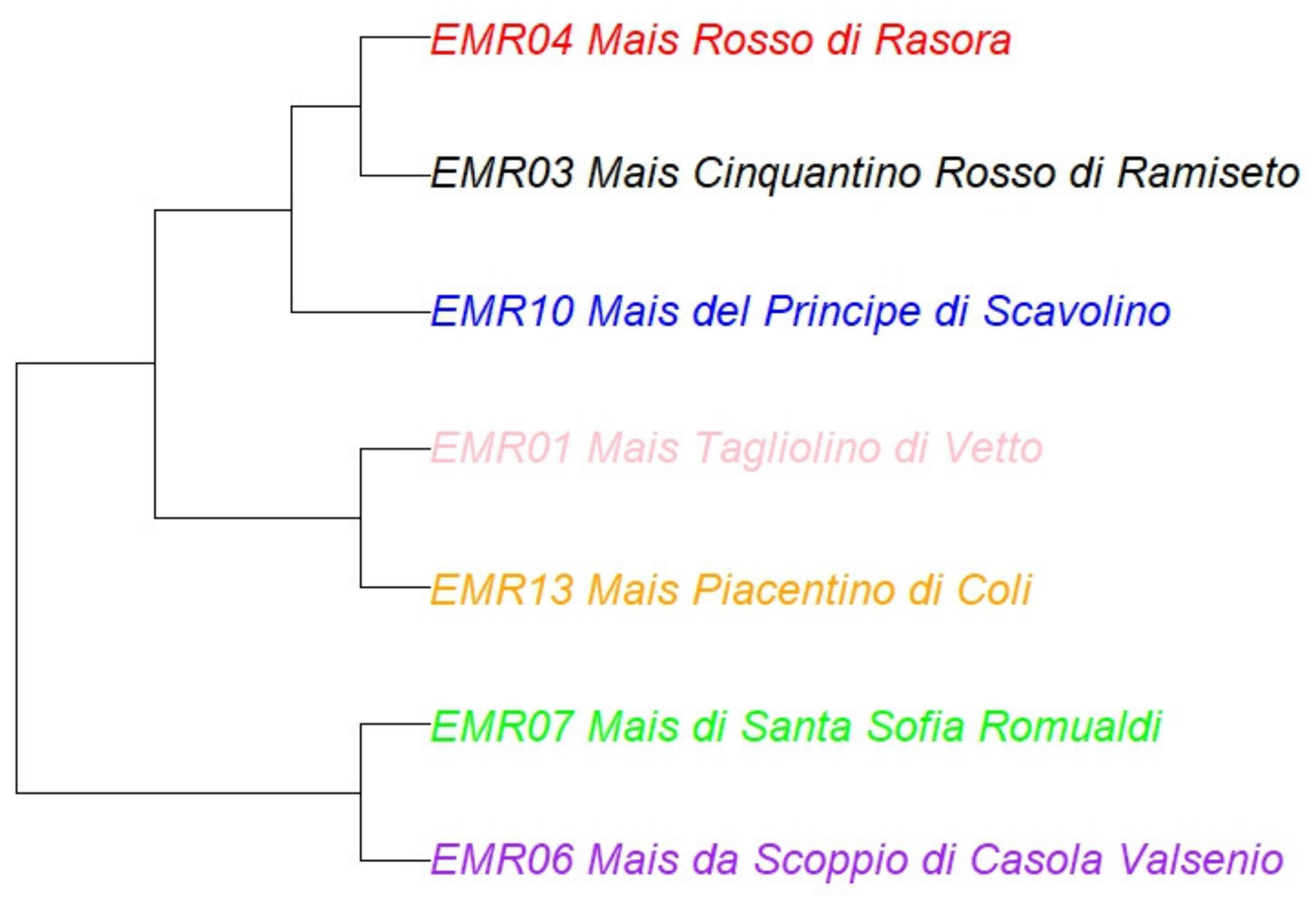
5. Cluster Analysis and Phylogenetic Tree
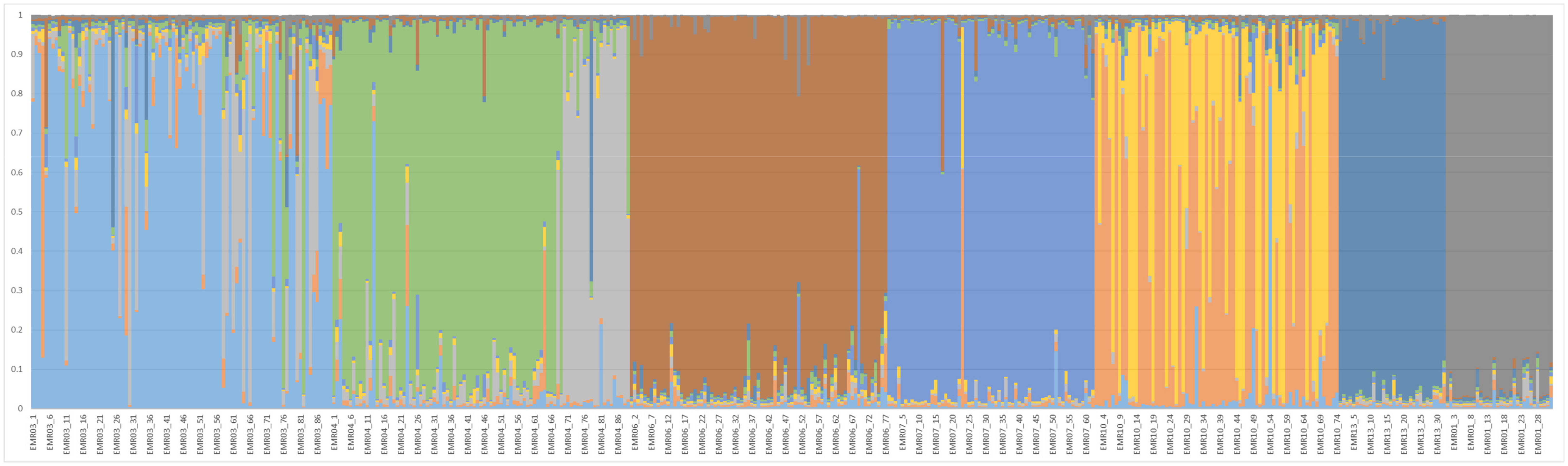
6. Structure Analysis
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brandolini, A.; Brandolini, A. Maize introduction, evolution and diffusion in Italy. Maydica 2009, 54, 233–242. [Google Scholar]
- Stagnati, L.; Martino, M.; Soffritti, G.; Lanubile, A.; Ravasio, A.; Marocco, A.; Rossi, G.; Busconi, M. Microsatellite and morphological characterization of three Rostrato di Val Chiavenna (Sondrio, Italy) maize (Zea mays L.) accessions. Genet. Resour. Crop Evol. 2021, 68, 3025–3038. [Google Scholar] [CrossRef]
- Brandolini, A.; Brandolini, A. Il Mais in Italia. Storia Naturale e Agricola, 2nd ed.; CRF Press: Bergamo, Italy, 2006. [Google Scholar]
- Eschholz, T.W.; Stamp, P.; Peter, R.; Leipner, J.; Hund, A. Genetic structure and history of Swiss maize (Zea mays L. ssp. mays) landraces. Genet. Resour. Crop Evol. 2010, 57, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Cömertpay, G.; Baloch, F.S.; Kilian, B.; Ülger, A.C.; Özkan, H. Diversity Assessment of Turkish Maize Landraces Based on Fluorescent Labelled SSR Markers. Plant Mol. Biol. Rep. 2012, 30, 261–274. [Google Scholar] [CrossRef]
- Ardenghi, N.M.G.; Rossi, G.; Guzzon, F. Back to beaked: Zea mays subsp. Mays Rostrata Group in northern Italy, refugia and revival of open-pollinated maize landraces in an intensive cropping system. PeerJ 2018, 6, e5123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedoya, C.A.; Dreisigacker, S.; Hearne, S.; Franco, J.; Mir, C.; Prasanna, B.M.; Taba, S.; Charcosset, A.; Warburton, M.L. Genetic diversity and population structure of native maize populations in Latin America and the Caribbean. PLoS ONE 2017, 12, e0173488. [Google Scholar] [CrossRef] [Green Version]
- Giupponi, L.; Pedrali, D.; Leoni, V.; Rodari, A.; Giorgi, A. The Analysis of Italian Plant Agrobiodiversity Databases Reveals That Hilly and Sub-Mountain Areas Are Hotspots of Herbaceous Landraces. Diversity 2021, 13, 70. [Google Scholar] [CrossRef]
- Canella, M.; Ardenghi, N.; Müller, J.; Rossi, G.; Guzzon, F. Plant agrobiodiversity conservation and use in intensive farming systems: N-Italy as a case study. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Cassani, E.; Puglisi, D.; Cantaluppi, E.; Landoni, M.; Giupponi, L.; Giorgi, A.; Pilu, R. Genetic studies regarding the control of seed pigmentation of an ancient European pointed maize (Zea mays L.) rich in phlobaphenes: The “Nero Spinoso” from the Camonica valley. Genet. Resour. Crop Evol. 2017, 64, 761–773. [Google Scholar] [CrossRef]
- Oppong, A.; Bedoya, C.A.; Ewool, M.B.; Asante, M.D.; Thompson, R.; Adu-Dapaah, H.; Lamptey, J.N.L.; Ofori, K.; Offei, S.; Warburton, M. Bulk genetic characterization of Ghanaian maize landraces using microsatellite markers. Maydica 2014, 59, 1–8. [Google Scholar]
- McLean-Rodríguez, F.D.; Costich, D.E.; Camacho-Villa, T.C.; Pè, M.E.; Dell’Acqua, M. Genetic diversity and selection signatures in maize landraces compared across 50 years of in situ and ex situ conservation. Heredity 2021, 126, 913–928. [Google Scholar] [CrossRef]
- Fenzi, M.; Couix, N. Growing maize landraces in industrialized countries: From the search for seeds to the emergence of new practices and values. Int. J. Sustain. Agric. Res. 2021, 1–19. [Google Scholar] [CrossRef]
- Guzzon, F.; Ardenghi, N.M.G.; Bodino, S.; Tazzari, E.; Rossi, G. Guida All’Agrobiodiverità Vegetale Della Provincia Di Pavia, 1st ed.; Pavia University Press: Pavia, Italy, 2019. [Google Scholar]
- Barcaccia, G.; Lucchin, M.; Parrini, P. Characterization of a flint maize (Zea mays var. indurata) Italian landrace, II. Genetic diversity and relatedness assessed by SSR and Inter-SSR molecular markers. Genet. Resour. Crop Evol. 2003, 50, 253–271. [Google Scholar] [CrossRef]
- Reif, J.C.; Hamrit, S.; Heckenberger, M.; Schipprack, W.; Maurer, H.P.; Bohn, M.; Melchinger, A.E. Genetic structure and diversity of European flint maize populations determined with SSR analyses of individuals and bulks. Theor. Appl. Genet. 2005, 111, 906–913. [Google Scholar] [CrossRef]
- Reif, J.C.; Warburton, M.L.; Xia, X.C.; Hoisington, D.A.; Crossa, J.; Taba, S.; Muminović, J.; Bohn, M.; Frisch, M.; Melchinger, A.E. Grouping of accessions of Mexican races of maize revisited with SSR markers. Theor. Appl. Genet. 2006, 113, 177–185. [Google Scholar] [CrossRef]
- Palumbo, F.; Galla, G.; Martínez-Bello, L.; Barcaccia, G. Venetian Local Corn (Zea mays L.) Germplasm: Disclosing the Genetic Anatomy of Old Landraces Suited for Typical Cornmeal Mush Production. Diversity 2017, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Troyer, F. Temperate Corn—Background, Behavior, and Breeding. In Specialty Corns, 2nd ed.; Hallauer, A.R., Ed.; CRC Press: Boca Raton, FL, USA, 2000; p. 74. ISBN 9780429127458. [Google Scholar]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. PowerMarker: Integrated analysis environment for genetic marker data. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [Green Version]
- Schliep, K.P. phangorn: Phylogenetic analysis in R. Bioinformatics 2011, 27, 592–593. [Google Scholar] [CrossRef] [Green Version]
- Clark, L.; Jasieniuk, M. polysat: An R package for polyploid microsatellite analysis. Mol. Ecol. Resour. 2011, 11, 562–566. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 8, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Giupponi, L.; Leoni, V.; Colombo, F.; Cassani, E.; Hejna, M.; Rossi, L.; Pilu, R. Characterization of “Mais delle Fiorine” (Zea mays L.) and nutritional, morphometric and genetic comparison with other maize landraces of Lombardy region (Northern Italy). Genet. Resour. Crop Evol. 2021, 68, 2075–2091. [Google Scholar] [CrossRef]
- Scarano, D.; Rao, R. DNA Markers for Food Products Authentication. Diversity 2014, 6, 579–596. [Google Scholar] [CrossRef]
- Ignjatović-Micić, D.; Drinić, S.M.; Nikolić, A.; Lazić-Jancić, V. SSR analysis for genetic structure and diversity determination of maize local populations from former Yugoslavia territories. Genetika 2008, 44, 1517–1524. [Google Scholar] [CrossRef]
- Qi-Lun, Y.; Ping, F.; Ke-Cheng, K.; Guang-Tang, P. Genetic diversity based on SSR markers in maize (Zea mays L.) landraces from Wuling mountain region in China. J. Genet. 2008, 87, 287–291. [Google Scholar] [CrossRef]
- Senior, M.L.; Murphy, J.P.; Goodman, M.M.; Stuber, C.W. Utility of SSRs for Determining Genetic Similarities an Relationships in Maize Using an Agarose Gel System. Crop Sci. 1998, 38, 1088–1098. [Google Scholar] [CrossRef]
- Sharma, L.; Prasanna, B.M.; Ramesh, B. Analysis of phenotypic and microsatellite-based diversity of maize landraces in India, especially from the North East Himalayan region. Genetica 2010, 138, 619–631. [Google Scholar] [CrossRef]
- Hartl, D.L.; Clark, A.G. Principles of Population Genetics; Sinauer: Sunderland, MA, USA, 1977. [Google Scholar]
- Bertolini, M.; Franchi, R.; Frisanco, F. Il Mais: Una Storia Anche Trentina, 1st ed.; Istituto Agrario di San Michele all’Adige: San Michele all’Adige, Italy, 2005; ISBN 88-7843-003-X. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Accession Name | Sampling Location | Plant Height (cm) | Ear Height (cm) | Tasseling (GDD) | Physiological Maturity (GDD) |
|---|---|---|---|---|---|---|
| EMR01 | Mais Tagliolino di Vetto | Vetto (Reggio Emilia, Italy) | 232 | 114 | 692 | 1446 |
| EMR03 | Mais Cinquantino Rosso di Ramiseto | Frazione Ramiseto, Ventasso (Reggio Emilia, Italy) | 222 | 111 | 646 | 1446 |
| EMR04 | Mais Rosso di Rasora | Frazione Rasora, Castiglione dei Pepoli (Bologna, Italy) | 190 | 93 | 677 | 1430 |
| EMR06 | Mais da Scoppio di Casola Valsenio | Casola Valsenio (Ravenna, Italy) | 210 | 82 | 739 | 1398 |
| EMR07 * | Mais di Santa Sofia Romualdi | Santa Sofia (Forlì-Cesena, Italy) | 200 | 70 | 618 | 1349 |
| EMR10 | Mais del Principe di Scavolino | Pennabilli (Rimini, Italy) | 226 | 73 | 593 | 1430 |
| EMR13 | Mais Piacentino di Coli | Frazione Fontana, Coli (Piacenza, Italy) | 240 | 90 | 646 | 1430 |
| Marker Name | Locus | Forward Primer 5′-3′ | Reverse Primer 5′-3′ | LG | Ta (°C) | Size (bp) | Reference |
|---|---|---|---|---|---|---|---|
| M302 | phi127 | ATATGCATTGCCTGGAACTGGAAGGA | [VIC] AATTCAAACACGCCTCCCGAGTGT | 2 | 58 | 100–120 | Palumbo et al., 2017 |
| M304 | phi076 | TTCTTCCGCGGCTTCAATTTGACC | [6FAM] GCATCAGGACCCGCAGAGTC | 4 | 58 | 150–200 | Palumbo et al., 2017 |
| M306 | phi031 | GCAACAGGTTACATGAGCTGACGA | [PET] CCAGCGTGCTGTTCCAGTAGTT | 6 | 58 | 180–220 | Palumbo et al., 2017 |
| M308 | umc1075 | GAGAGATGACAGACACATCCTTGG | [6FAM] ACATTTATGATACCGGGAGTTGGA | 8 | 58 | 130–150 | Palumbo et al., 2017 |
| M310 | phi084 | AGAAGGAATCCGATCCATCCAAGC | [PET] CACCCGTACTTGAGGAAAACCC | 10 | 58 | 140–170 | Palumbo et al., 2017 |
| M24 | umc1327 | AGGGTTTTGCTCTTGGAATCTCTC | [NED] GAGGAAGGAGGAGGTCGTATCGT | 8 | 64 | 100–120 | MaizeGDB |
| M33 | p-bnlg176 | AGTTCACGTCCAGCTGAATGACAG | [6FAM] CGCGCATCGCATGCTTATCCTA | 1 | 62 | 140–170 | MaizeGDB |
| M78 | umc1941 | ACGACGAGACTCTGTTCTGGTTCT | [NED] AGGAGGATTACGTCAATCTGTTCG | 5 | 64 | 110–130 | MaizeGDB |
| M90 | umc1401 | CTCTGGTCCATCCTCATCGACT | [PET] TCTCTTGATCACATATCGATCCCA | 7 | 62 | 180–200 | MaizeGDB |
| M193 | umc1786 | ACCGTGACTTCCTCCTCATAACTG | [VIC] CATTTTTCGCATTTAGGAAATCCA | 8 | 60 | 180–220 | MaizeGDB |
| Landrace | Locus | Allele | Allele Frequency |
|---|---|---|---|
| EMR03 | umc1327 | 82 | 0.105 |
| EMR03 | umc1786 | 148 | 0.006 |
| EMR04 | umc1327 | 80 | 0.006 |
| EMR04 | umc1786 | 144 | 0.012 |
| EMR04 | p-bnlg176 | 126 | 0.017 |
| EMR04 | p-bnlg176 | 128 | 0.040 |
| EMR04 | umc1401 | 157 | 0.006 |
| EMR07 | phi031 | 192 | 0.050 |
| EMR07 | umc1327 | 74 | 0.008 |
| EMR07 | umc1786 | 124 | 0.009 |
| EMR10 | phi076 | 153 | 0.042 |
| EMR10 | phi084 | 156 | 0.021 |
| EMR13 | umc1075 | 120 | 0.016 |
| EMR13 | p-bnlg176 | 132 | 0.031 |
| EMR13 | p-bnlg176 | 147 | 0.188 |
| Locus | Na | Ne | I | Ho | uHe | PIC | F | FIS | FST | Nm |
|---|---|---|---|---|---|---|---|---|---|---|
| phi127 | 3.00 | 1.88 | 0.68 | 0.41 | 0.41 | 0.51 | −0.02 | −0.02 | 0.30 | 0.60 |
| phi076 | 2.71 | 2.03 | 0.79 | 0.49 | 0.51 | 0.53 | 0.04 | 0.04 | 0.16 | 1.33 |
| phi031 | 4.00 | 2.10 | 0.84 | 0.39 | 0.46 | 0.59 | 0.13 | 0.15 | 0.30 | 0.60 |
| umc1075 | 4.00 | 2.45 | 0.95 | 0.39 | 0.53 | 0.75 | 0.28 | 0.25 | 0.34 | 0.48 |
| phi084 | 2.43 | 1.46 | 0.44 | 0.28 | 0.27 | 0.38 | 0.05 | −0.03 | 0.41 | 0.36 |
| umc1327 | 3.14 | 1.89 | 0.69 | 0.49 | 0.40 | 0.50 | −0.25 | −0.25 | 0.19 | 1.07 |
| p-bnlg176 | 4.00 | 2.11 | 0.89 | 0.36 | 0.50 | 0.59 | 0.29 | 0.28 | 0.28 | 0.63 |
| umc1941 | 4.00 | 2.38 | 0.93 | 0.51 | 0.50 | 0.72 | −0.01 | −0.03 | 0.34 | 0.49 |
| umc1401 | 3.57 | 2.12 | 0.79 | 0.61 | 0.44 | 0.6 | −0.33 | −0.38 | 0.28 | 0.63 |
| umc1786 | 3.57 | 1.68 | 0.60 | 0.24 | 0.34 | 0.55 | 0.23 | 0.28 | 0.46 | 0.29 |
| All loci | 3.44 | 2.01 | 0.76 | 0.42 | 0.44 | 0.57 | 0.05 | 0.03 | 0.31 | 0.65 |
| dev.st | 0.16 | 0.09 | 0.05 | 0.03 | 0.03 | 0.11 | 0.04 | 0.07 | 0.03 | 0.10 |
| EMR01_Tagliolino | 1.80 | 1.63 | 0.45 | 0.24 | 0.30 | 0.17 | 0.19 | 0.52 | 0.23 | |
| EMR03_Cinquantino_Rosso | 4.20 | 2.32 | 0.95 | 0.49 | 0.53 | 0.11 | 0.09 | 0.15 | 1.46 | |
| EMR04_Rasora | 4.30 | 2.28 | 0.93 | 0.58 | 0.52 | −0.09 | −0.11 | 0.17 | 1.25 | |
| EMR06_Mais_da_Scoppio | 3.60 | 2.14 | 0.80 | 0.48 | 0.46 | −0.04 | −0.04 | 0.27 | 0.68 | |
| EMR07_Santa_Sofia | 3.90 | 1.80 | 0.72 | 0.34 | 0.40 | 0.19 | 0.15 | 0.36 | 0.44 | |
| EMR10_Principe_di_Scavolino | 3.80 | 2.18 | 0.90 | 0.53 | 0.51 | −0.05 | −0.05 | 0.19 | 1.09 | |
| EMR13_Piacentino | 2.50 | 1.72 | 0.56 | 0.26 | 0.34 | 0.14 | 0.23 | 0.46 | 0.29 | |
| All pop | 3.44 | 2.01 | 0.76 | 0.42 | 0.44 | 0.05 | 0.05 | 0.30 | 0.58 | |
| dev.st | 0.16 | 0.09 | 0.05 | 0.03 | 0.03 | 0.04 | 0.12 | 0.14 | 0.46 |
| EMR03 | EMR04 | EMR06 | EMR07 | EMR10 | EMR13 | EMR01 | |
|---|---|---|---|---|---|---|---|
| 0.000 | EMR03 | ||||||
| 0.170 | 0.000 | EMR04 | |||||
| 0.473 | 0.588 | 0.000 | EMR06 | ||||
| 0.537 | 0.609 | 0.395 | 0.000 | EMR07 | |||
| 0.213 | 0.472 | 0.503 | 0.514 | 0.000 | EMR10 | ||
| 0.270 | 0.457 | 0.742 | 0.869 | 0.542 | 0.000 | EMR13 | |
| 0.359 | 0.505 | 0.677 | 0.618 | 0.664 | 0.253 | 0.000 | EMR01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stagnati, L.; Soffritti, G.; Martino, M.; Lanubile, A.; Desiderio, F.; Ravasio, A.; Marocco, A.; Rossi, G.; Busconi, M. Morphological and Genetic Characterization of Local Maize Accessions from Emilia Romagna Region, Italy. Sustainability 2022, 14, 91. https://doi.org/10.3390/su14010091
Stagnati L, Soffritti G, Martino M, Lanubile A, Desiderio F, Ravasio A, Marocco A, Rossi G, Busconi M. Morphological and Genetic Characterization of Local Maize Accessions from Emilia Romagna Region, Italy. Sustainability. 2022; 14(1):91. https://doi.org/10.3390/su14010091
Chicago/Turabian StyleStagnati, Lorenzo, Giovanna Soffritti, Michelangelo Martino, Alessandra Lanubile, Francesca Desiderio, Adriano Ravasio, Adriano Marocco, Graziano Rossi, and Matteo Busconi. 2022. "Morphological and Genetic Characterization of Local Maize Accessions from Emilia Romagna Region, Italy" Sustainability 14, no. 1: 91. https://doi.org/10.3390/su14010091
APA StyleStagnati, L., Soffritti, G., Martino, M., Lanubile, A., Desiderio, F., Ravasio, A., Marocco, A., Rossi, G., & Busconi, M. (2022). Morphological and Genetic Characterization of Local Maize Accessions from Emilia Romagna Region, Italy. Sustainability, 14(1), 91. https://doi.org/10.3390/su14010091