
Land Use, Landform, and Soil Management as Determinants of Soil Physicochemical Properties and Microbial Abundance of Lower Brahmaputra Valley, India

 , , , ,
, , , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Statistical Analysis
3. Results
3.1. Soil Physicochemical Properties
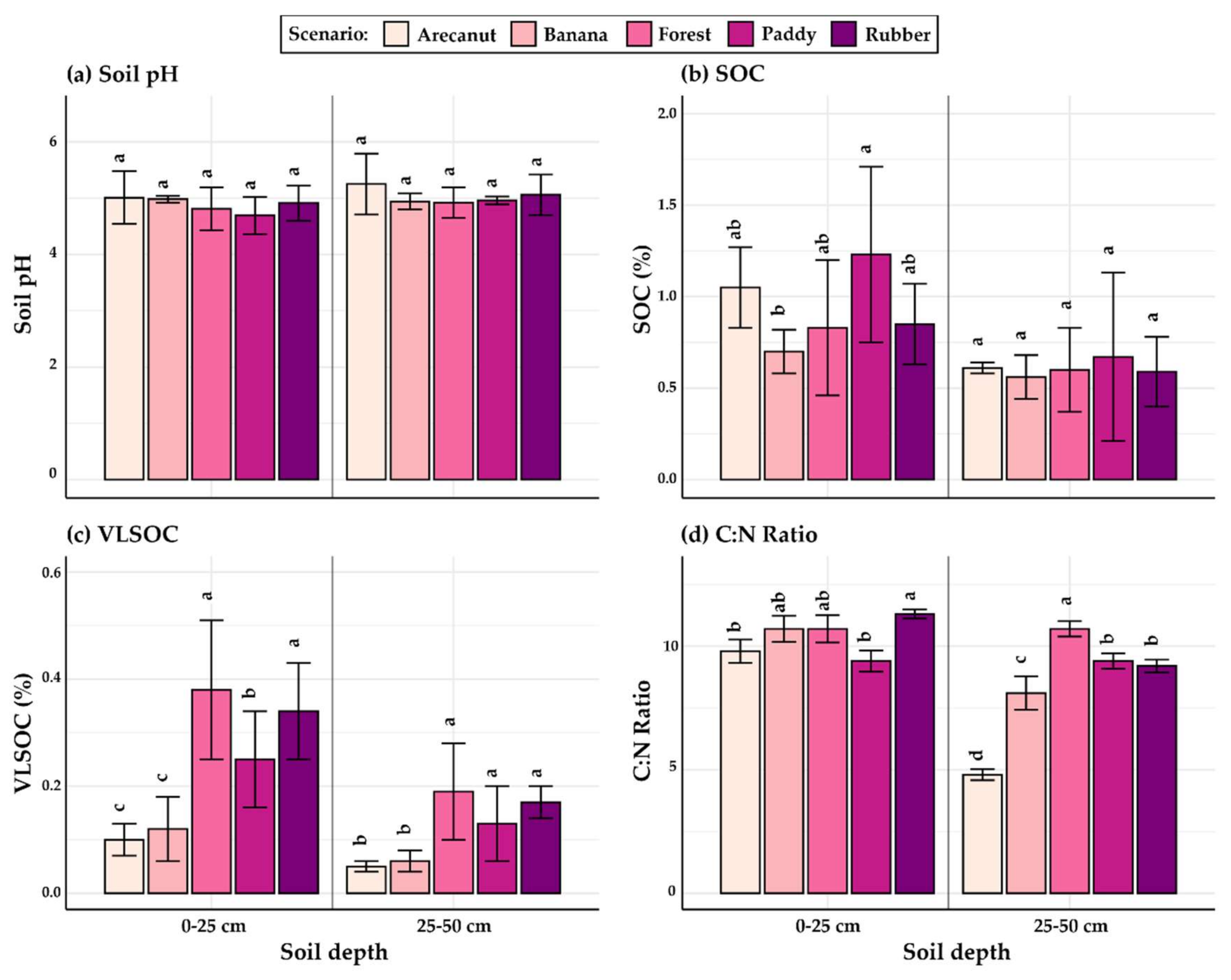
3.1.1. 0 to 25 cm Depth
3.1.2. 25 to 50 cm Depth
3.2. Microbial Population
3.3. Correlation Analysis
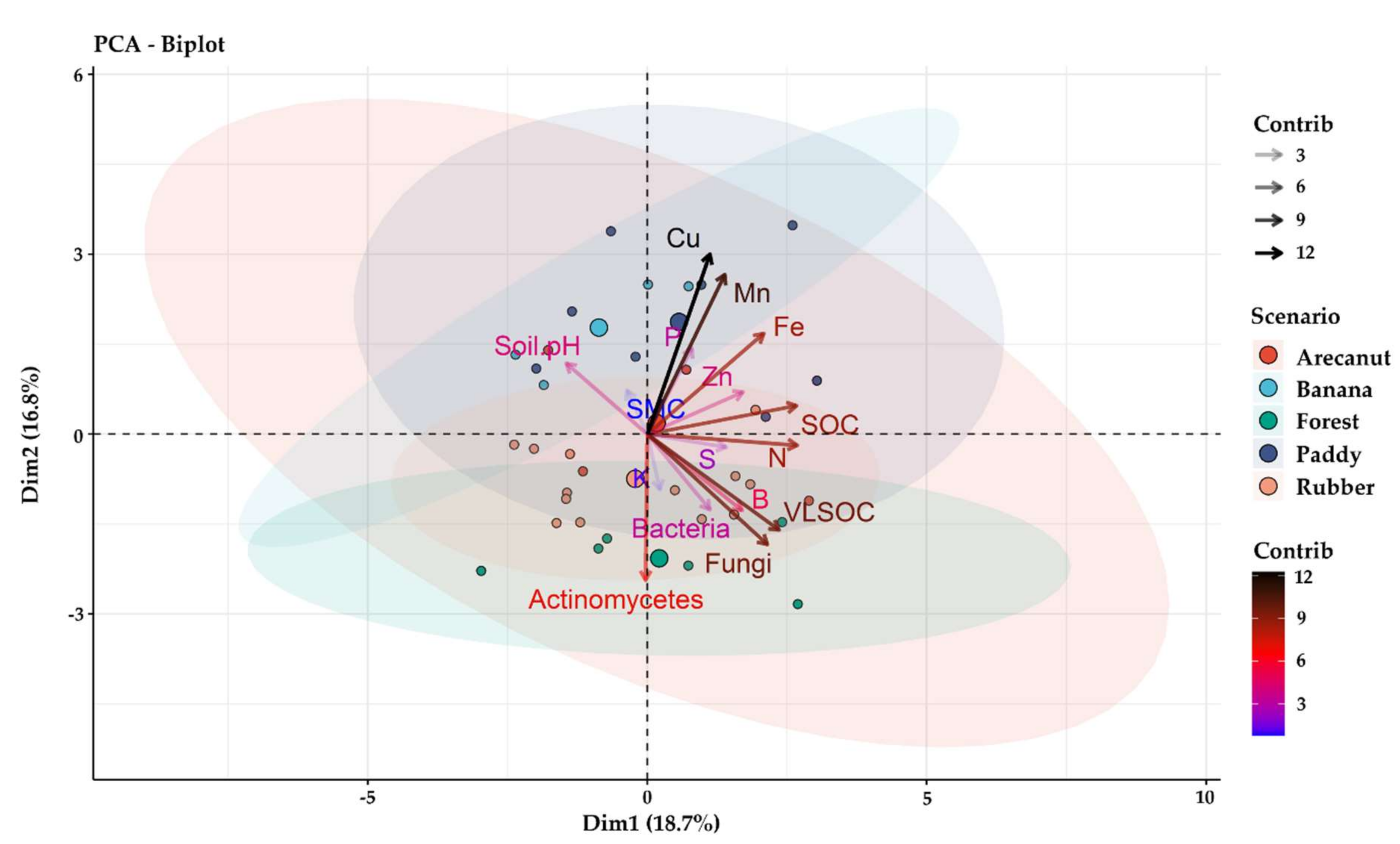
3.4. Principal Component Analysis (PCA) of Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Leeuwen, J.P.; Djukic, I.; Bloem, J.; Lehtinen, T.; Hemerik, L. Effects of land use on soil microbial biomass, activity, and community structure at different soil depths in Danube floodplain. Eur. J. Soil. Biol. 2017, 79, 14–20. [Google Scholar] [CrossRef]
- Bini, D.; dos Santos, C.A.; do Carmo, K.B.; Kishino, N.; Andrade, G.; Zangaro, W.; Nogueira, M.A. Effects of land use on soil organic carbon and microbial process associated with soil health in Southern Brazil. Eur. J. Soil. Biol. 2013, 55, 117–123. [Google Scholar] [CrossRef]
- Cheng, X.; Yang, Y.; Li, M.; Dou, X.; Zhang, Q. The impact of agricultural land use changes on soil organic carbon dynamics in the Danjiangkou Reservoir area of China. Plant. Soil 2013, 366, 415–424. [Google Scholar] [CrossRef]
- García-Palacios, P.; Maestre, F.T.; Kattge, J.; Wall, D.H. Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecol. Lett. 2013, 16, 1045–1053. [Google Scholar] [CrossRef] [Green Version]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setcalca, H.; van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2003, 304, 1629–1633. [Google Scholar] [CrossRef]
- Kardol, P.; Wardle, D.A. How understanding aboveground-belowground linkages can assist restoration ecology. Trends Ecol. Evol. 2010, 25, 670–679. [Google Scholar] [CrossRef]
- Lepcha, N.T.; Devi, N.B. Effect of land use, season, and soil depth on soil microbial biomass carbon of Eastern Himalayas. Ecol. Process. 2020, 9, 65. [Google Scholar] [CrossRef]
- Pennanen, T.; Fritze, H.; Vanhala, P.; Kiikkila, O.; Neuvonen, S.; Baath, E. Structure of a microbial community in soil after prolonged addition of low levels of simulated acid rain. Appl. Environ. Microbiol. 1998, 64, 2173–2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, J.S.; Gupta, V.K. Soil microbial biomass: A key soil driver in management of ecosystem functioning. Sci. Total Environ. 2018, 634, 497–500. [Google Scholar] [CrossRef]
- Mahato, M.K.; Jana, N.C. Impact of landform on agricultural land use pattern: A case study of Salda River Basin in Purulia District, West Bengal. J. Geogr. Environ. Earth Sci. Internat. 2019, 21, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Zackary, G.; Mainuri, J.O. Linking landforms and land use to land degradation in the Middle River Njoro Watershed. Internat. Soil Water Conservat. Res. 2014, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tischer, A.; Potthast, K.; Hamer, U. Land-use and soil depth affect resource and microbial stoichiometry in a tropical mountain rainforest region of southern Ecuador. Oecologia 2014, 175, 375–393. [Google Scholar] [CrossRef] [PubMed]
- Spohn, M.; Chodak, M. Microbial respiration per unit biomass increases with carbon-to-nutrient ratios in forest soils. Soil Biol. Biochem. 2015, 81, 128–133. [Google Scholar] [CrossRef]
- Saikia, L.; Mahanta, C.; Mukherjee, A. Erosion–deposition and land use/land cover of the Brahmaputra River in Assam. India. J. Earth Syst. Sci. 2011, 128, 211. [Google Scholar] [CrossRef] [Green Version]
- Hazarika, N.; Das, A.K.; Borah, S.B. Assessing land-use changes driven by river dynamics in chronically flood affected Upper Brahmaputra plains, India, using RS-GIS techniques. Egypt. J. Remote Sens. Space Sci. 2015, 18, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Sinha, R.; Tandon, S.K. Indus-Ganga-Brahmaputra plains: The alluvial landscape. Landsc. Lanforms India 2014, 53–63. [Google Scholar] [CrossRef]
- Baruah, B.K.; Das, B.; Medhi, C.; Mishra, A.K. Fertility status of soil in the tea garden belts of Golaghat district, Assam, India. J. Chem. 2013, 2013, 983297. [Google Scholar] [CrossRef] [Green Version]
- Reza, S.K.; Baruah, U.; Nayak, D.C.; Dutta, D.; Singh, S.K. Effects of land-use on soil physical, chemical and microbial properties in humid subtropical Northeastern India. Natl. Acad. Sci Lett. 2018, 41, 141–145. [Google Scholar] [CrossRef]
- Choudhury, B.U.; Ansari, M.A.; Chakraborty, M. Effect of land-use change along altitudinal gradients on soil micronutrients in the mountain ecosystem of Indian (Eastern) Himalaya. Sci. Rep. 2021, 11, 14279. [Google Scholar] [CrossRef]
- Gangopadhyay, S.; Ray, S.K. Soil chemical and mineralogical characteristics under major land use systems on upper Brahmaputra valley of Assam. Clay. Res. 2020, 39, 56–66. [Google Scholar] [CrossRef]
- Das Gupta, A.B.; Biswas, A.K. Geology of Assam; Geological Society of India, Bangalore: Bengaluru, India, 2002; pp. 45–83. [Google Scholar]
- Velayutham, M.; Mandal, D.K.; Mandal, C.; Sehgal, J. Agro-ecological sub-regions of India for planning and development. NBSS & LUP Publ. 1999, 35, 372. [Google Scholar]
- Keys to Soil Taxonomy. By Soil Survey Staff. United States Department of Agriculture. Natural Resources Conservation Service. Available online: https://www.nrcs.usda.gov/wps/PA_NRCSConsumption/download?cid=stelprdb1252094&ext=pdf (accessed on 15 July 2021).
- Lindsay, W.L.; Norvell, W.A. Development of a DTPA soil test for zinc, iron, manganese, and copper. Soil Sci. Soc. Am. J. 1978, 42, 421–428. [Google Scholar] [CrossRef]
- Jackson, M.L. Soil Chemical Analysis; Prentice-Hall: Englewood Cliffs, NJ, USA, 1958; p. 498. [Google Scholar]
- Subbiah, B.W.; Asijia, G.L. A rapid procedure for the estimation of available nitrogen in soil. Curr. Sci. 1965, 25, 259–260. [Google Scholar]
- Bray, R.H.; Kurtz, L.T. Determination of total organic and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–45. [Google Scholar] [CrossRef]
- Jackson, M.L. Soil Chemical Analysis; Prentice Hall India Pvt. Ltd.: India, New Delhi, 1967. [Google Scholar]
- Chesnin, L.; Yein, C.H. Turbidimetric determinations of available sulphates. Soil Sci. Soc. Am. Proc. 1951, 15, 149–151. [Google Scholar] [CrossRef]
- Berger, K.C.; Truog, E. Boron determination in soils and plants. Ind. Eng. Chem. Anal. Ed. 1939, 11, 540–545. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of the Degterett method for determining soil organic matter and proposal modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Chan, K.Y.; Bowman, A.; Oates, A. Oxidizible organic carbon fractions and soil quality changes in an oxic paleustalf under different pasture leys. Soil Sci. 2001, 166, 61–67. [Google Scholar] [CrossRef]
- Gardner, W.H. Water content. In. Methods of Soil Analysis, Part I. ed. CA Black et al., (Monograph No. 9) Agronomy Series. Amer. Soc. Agron. 1965, 1, 82–127. [Google Scholar]
- Robinson, G.W. Note on the mechanical analysis of humus soils. Agr. Sci. 1922, 12, 287–291. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley and Sons: New York, NY, USA, 1984; p. 680. [Google Scholar]
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Pearson, K. Notes on regression and inheritance in the case of two parents. Proc. R. Soc. 1895, 58, 240–242. [Google Scholar] [CrossRef] [Green Version]
- Doetterl, S.; Six, J.; Van Wesemael, B.; Van Oost, K. Carbon cycling in eroding landscapes: Geomorphic controls on soil organic C pool composition and C stabilization. Glob. Chang. Biol. 2012, 18, 2218–2232. [Google Scholar] [CrossRef]
- Fissore, C.; Dalzell, B.J.; Berhe, A.A.; Voegtle, M.; Evans, M.; Wu, A. Influence of topography on soil organic carbon dynamics in a Southern California grassland. Catena 2017, 149, 140–149. [Google Scholar] [CrossRef]
- Burst, M.; Chauchard, S.; Dambrine, E.; Dupouey, J.L.; Amiaud, B. Distribution of soil properties along forest-grassland interfaces: Influence of permanent environmental factors or land-use aftereffects? Agric. Ecosyst. Environ. 2020, 289, 106739. [Google Scholar] [CrossRef]
- Paudel, S.; Sah, J.P. Physicochemical characters of soil in tropical sal (Shorea robusta Gaertn.) forests in eastern Nepal. Himal. J. Sci. 2003, 1, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, L.; Lu, J.; Chen, W.; Wei, G.; Lin, Y. Topography affects the soil conditions and bacterial communities along a restoration gradient on Loess-Plateau. Appl. Soil Ecol. 2020, 150, 103471. [Google Scholar] [CrossRef]
- Gonnety, J.T.; Assémien, E.F.; Guéi, A.M.; Aya, A.N.; Djina, Y.; Koné, A.W.; Tondoh, J.E. Effect of land-use types on soil enzymatic activities and chemical properties in semi-deciduous forest areas of Central-West Côte d’Ivoire. Biotechnol. Agron. Soc. Environ. 2013, 4, 478–485. [Google Scholar]
- George, N.; Killur, R.R.B.; Cornelio, D.L. Land use conversion and soil properties in a lowland tropical landscape of Papua New Guinea. J. Manajemen. Hutan Trop. 2013, 1, 39. [Google Scholar] [CrossRef]
- Malik, A.A.; Puissant, J.; Buckeridge, K.M.; Goodall, T.; Jehmlich, N.; Chowdhury, S.; Gweon, H.S.; Peyton, J.M.; Mason, K.E.; van Agtmaal, M.; et al. Land use driven change in soil pH affects microbial carbon cycling processes. Nat. Commun. 2018, 9, 3591. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Gan, P.; Chen, A. Environmental controls on soil pH in planted forest and its response to nitrogen deposition. Environ. Res. 2019, 172, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Han, G.; Liu, M.; Li, X. Effects of soil pH and texture on soil carbon and nitrogen in soil profiles under different land uses in Mun River Basin, Northeast Thailand. Peer J. 2019, 7, e7880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assefa, F.; Elias, E.; Soromessa, T.; Ayele, G.T. Effect of Changes in Land-Use Management Practices on Soil Physicochemical Properties in Kabe Watershed, Ethiopia. Air Soil Water Res. 2020, 13, 1–16. [Google Scholar] [CrossRef]
- Singh, P.; Benbi, D.K. Soil organic carbon pool changes in relation to slope position and land-use in Indian lower Himalayas. Catena 2018, 166, 171–180. [Google Scholar] [CrossRef]
- McGrath, D.; Smith, C.; Gholz, H.; Oliveira, F.d.A. Effects of land-use change on soil nutrient dynamics in Amazônia. Ecosystems 2001, 4, 625–645. Available online: http://www.jstor.org/stable/3658623 (accessed on 18 September 2021). [CrossRef]
- Fontaine, S.; Barot, S.; Barré, P. Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef]
- Smith, S.V.; Renwick, W.H.; Buddemeier, R.W.; Crossland, C.J. Budgets of soil erosion and deposition for sediments and sedimentary organic carbon across the conterminous United States. Glob. Biogeochem. Cycles 2001, 15, 697–707. [Google Scholar] [CrossRef] [Green Version]
- Berhe, A.A.; Kleber, M. Erosion, deposition, and the persistence of soil organic matter: Mechanistic considerations and problems with terminology. Earth Surf. Process. Landf. 2013, 38, 908–912. [Google Scholar] [CrossRef]
- Doetterl, S.; Berhe, A.A.; Nadeu, E.; Wang, Z.; Sommer, M.; Fiener, P. Erosion, deposition, and soil carbon: A review of process-level controls, experimental tools, and models to address C cycling in dynamic landscapes. Earth Sci. Rev. 2016, 154, 102–122. [Google Scholar] [CrossRef]
- Haque, M.M.; Biswas, J.C.; Maniruzaman, M.; Akhter, S.; Kabir, M.S. Carbon sequestration in paddy soil as influenced by organic and inorganic amendments. Carbon Manag. 2020, 11, 231–239. [Google Scholar] [CrossRef]
- Negasa, D.J. Effects of land use types on selected soil properties in Central Highlands of Ethiopia. App. Environ. Soil Sci. 2020, 6, 1–9. [Google Scholar] [CrossRef]
- Uzoho, B.U.; Ekeh, C. Potassium status of soils in relation to land use types in Ngor-Okpala, Southeastern Nigeria. J. Nat. Sci. Res. 2014, 4, 2224–3186. [Google Scholar]
- Erickson, G.; Klopfenstein, T.; Walters, D.; Lesoing, G. Nutrient Balance of Nitrogen, Organic Matter, Phosphorous and Sulfur the Feedlot. Neb. Beef Cattle Rep. 1998, 69, 86–87. [Google Scholar]
- Sharma, Y.K.; Sharma, A.; Sharma, S.K. An appraisal of physico-chemical characteristics and soil fertility status of forest and rice land use systems in Mokokchung district of Nagaland. J. Indian Soc. Soil Sci. 2013, 61, 38–43. [Google Scholar]
- Takkar, P.N. Micronutrient research and sustainable agricultural productivity in India. J. Indian Society Soil Sci. 1996, 44, 562–581. [Google Scholar]
- Ray, P.; Gogoi, S.N.; Bandyopadhyay, S.; Jena, R.K.; Ramachandran, S.; Sharma, G.K.; Sah, K.D.; Singh, S.K.; Ray, S.K. Soil-cum-nutrient management package for achieving high yield and quality of mulberry (Morus indica L.) leaf in acid soil of Northeastern India. J. Plant. Nutr. 2021, 44, 2501–2513. [Google Scholar] [CrossRef]
- Ray, P.; Gogoi, S.N.; Bandyopadhyay, S.; Padua, S.; Jena, R.K.; Roy, P.D.; Ramachandran, S.; Sharma, G.K.; Sah, K.D.; Trivedy, K.; et al. Fertility status of mulberry (Morus indica L.) growing soils of upper Brahmaputra valley region of northeastern India. Range Manag. Agroforest. 2018, 39, 147–155. [Google Scholar] [CrossRef]
- Bansal, R.L.; Takkar, P.N. Micronutrient status of soils in Amritsar district. Indian J. Ecol. 1986, 13, 158–160. [Google Scholar]
- Bhuyan, N.; Barua, N.G.; Borah, D.K.; Bhattacharyya, D.; Basumatari, A. Georeferenced micronutrient status in soils of Lakhimpur district of Assam. J. Indian Soc. Soil Sci. 2014, 62, 102–107. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Ray, P.; Padua, S.; Ramachandran, S.; Jena, R.K.; Roy, P.D.; Dutta, D.P.; Singh, S.K.; Ray, S.K. Priority zoning of available micronutrients in the soils of agro-ecological sub-regions of North-East India using geospatial techniques. Agric. Res. 2018, 7, 200–214. [Google Scholar] [CrossRef]
- Dai, L.; Guo, X.; Ke, X.; Du, Y.; Zhang, F.; Cao, G. The variation in soil water retention of alpine shrub meadow under different degrees of degradation on northeastern Qinghai-Tibetan plateau. Plant. Soil 2021, 458, 231–244. [Google Scholar] [CrossRef]
- Kumar, V.; Gathala, M.K.; Saharawat, Y.S.; Parihar, C.M.; Kumar, R.; Kumar, R.; Jat, M.L.; Jat, A.S.; Mahala, D.M.; Kumar, L.; et al. Impact of tillage and crop establishment methods on crop yields, profitability, and soil physical properties in rice–wheat system of Indo-Gangetic Plains of India. Soil Use Manag. 2019, 35, 303–313. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- LaMontagne, M.G.; Schimel, J.P.; Holden, P.A. Comparison of subsurface and surface soil bacterial communities in California grassland as assessed by terminal restriction fragment length polymorphisms of PCR amplified 16S rRNA genes. Microb. Ecol. 2003, 46, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Xia, B.; Treves, D.S.; Wu, L.Y.; Marsh, T.L.; O’Neill, R.V.; Palumbo, A.V.; Tiedje, J.M. Spatial and resource factors influencing high microbial diversity in soil. Appl. Environ. Microbiol. 2002, 68, 326–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SchnuÈrer, J.; Clarholm, M.; Bastrom, S.; Rosswall, T. 1986. Effects of moisture on soil microorganisms and nematodes: A field experiment. Microb. Ecol. 1986, 12, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.S.; Bais, H.P.; Grotewold, E.; Vivanco, J.M. Root exudation and rhizosphere biology. Plant. Physiol. 2003, 132, 44–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beare, M.H.; Parmelee, R.W.; Hendrix, P.F.; Cheng, W.; Coleman, D.C.; Grossley, D.A., Jr. Microbial and faunal interactions and effects on litter nitrogen and decomposition in agroecosystems. Ecol. Monogr. 1992, 62, 569–591. [Google Scholar] [CrossRef] [Green Version]
- Neely, C.L.; Beare, M.H.; Hargrove, W.H.; Coleman, D.C. Relationships between fungal and bacterial substrate-induced respiration, biomass, and plant residue decomposition. Soil Biol. Biochem. 1991, 23, 947–954. [Google Scholar] [CrossRef]
- Holland, E.A.; Coleman, D.C. Litter placement effects on microbial and organic matter dynamics in an agroecosystem. Ecology 1987, 68, 425–433. [Google Scholar] [CrossRef]
- Frey, S.D.; Elliott, E.T.; Paustian, K. Bacterial and fungal abundance and biomass in conventional and no-tillage agroecosystems along two climatic gradients. Soil Biol. Biochem. 1999, 31, 573–585. [Google Scholar] [CrossRef]
- Unger, I.M.; Kennedy, A.C.; Muzika, R.M. Flooding effects on soil microbial communities. App. Soil Ecol. 2009, 42, 1–8. [Google Scholar] [CrossRef]
- Silva, C.C.; Guido, M.L.; Ceballos, J.M.; Marsch, R.; Dendooven, L. Production of carbon dioxide and nitrous oxide in alkaline saline soil of Texcoco at different water contents amended with urea: A laboratory study. Soil Biol. Biochem. 2008, 40, 1813–1822. [Google Scholar] [CrossRef]
- Hota, S.; Jena, R.K.; Ray, P.; Ramachandran, S.; Mourya, K.K.; Sharma, G.K.; Ray, S.K. Fertility capability classification of soils under dominant land uses of North-Eastern India. In Proceedings of the conference: Soil-Plant-Animal and Human Health Continumm, 5th International Agronomy Congress, Hyderabad, India, 23–27 November 2021. [Google Scholar]
- Zakalyukina, Y.V.; Zenova, G.M.; Zvyagintsev, D.G. Acidophilic Soil Actinomycetes. Microbiology 2002, 71, 342–345. [Google Scholar] [CrossRef]
- Poomthongdee, N.; Duangmal, K.; Pathom-aree, W. Acidophilic actinomycetes from rhizosphere soil: Diversity and properties beneficial to plants. J. Antibiot. 2015, 68, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wang, L.; Cui, Q.; Huang, Y.; Liu, Z.; Zheng, G.; Goodfellow, M. Neutrotolerant acidophilic Streptomyces species isolated from acidic soils in China: Streptomyces guanduensis sp. nov., Streptomyces paucisporeus sp. nov., Streptomyces rubidus sp. nov. and Streptomyces yanglinensis sp. nov. Int. J. Syst. Evol. Microbiol. 2006, 56, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani-Nasrabadi, R.; Greiner, R.; Alikhani, H.A.; Hamedi, J.; Yakhchali, B. Distribution of actinomycetes in different soil ecosystems and effect of media composition on extracellular phosphatase activity. J. Soil Sci. Plant. Nutr. 2013, 13, 223–236. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Scenario | Land Use (Name of Scenario) | Landform | Slope Gradient (%) | Drainage | Fertilization and Management | Cropping System | Other Management Practices |
|---|---|---|---|---|---|---|---|
| 1 | Paddy | Younger and older alluvial plain | 0–1% and 1–3% | Poor | Cow dung manure application @ 15 to 20 q/ha before transplanting. | Kharif monocropping | Incorporation of stubble, puddling, conventional tillage |
| 2 | Banana | Older alluvial plain and Char land | 1–5% | Moderately well | Basal application SSP @ 100 g/plant. Urea @ 100 g/plant 45 days after planting (DAP) and MOP @ 250 g/plant starting from 90 DAP. All are repeated at 45-day intervals. | Perennial monocropping | Earthing-up operations during planting and fertilizer application |
| 3 | Arecanut | Older alluvial plain and Char land | 1–5% | Moderately well | Occasional application of SSP. Cow dung manure @ 5 kg/plant every 5 to 6 months. | Perennial monocropping | No other operations |
| 4 | Rubber | Upland | 3–8% | Well | Urea @ 350 kg/ha and SSP @ 200 to 250 kg during initial plantation. | Perennial monocropping | Leaf litter left on surface |
| 5 | Sal forest | Upland | 3–8% | Well | No fertilizer management. | Recently planted forest | Leaf litter left on surface |
| Scenario | Sand (%) | Silt (%) | Clay (%) | Soil Texture | N (Kg ha−1) | P (Kg ha−1) | K (Kg ha−1) | S (ppm) | B (ppm) | Fe (ppm) | Mn (ppm) | Zn (ppm) | Cu (ppm) | SMC (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Paddy (n = 8) | 21.6 ± 2.1 | 31.2 ± 2.4 | 47.2 ± 3.6 | Clay loam to Clay | 378.9 ± 11.18 a | 49.4 ± 9.28 a | 153.1 ± 33.2 b | 15.1 ± 4.3 a | 0.32 ± 0.03 ab | 71.5 ± 5.30 ab | 21.14 ± 1.8 a | 0.65 ± 0.15 b | 0.74 ± 0.06 a | 23.16 ± 2.5 a |
| Banana (n = 4) | 14.0 ± 3.3 | 51.3 ± 4.1 | 34.7 ± 4.8 | Silty clay loam | 353.1 ± 72.74 a | 55.1 ± 7.50 a | 254.7 ± 85.5 b | 11.4 ± 4.94 a | 0.21 ± 0.04 b | 103.6 ± 4.12 a | 14.45 ± 4.1 ab | 0.60 ± 0.02 b | 2.25 ± 0.05 ab | 12.9 ± 2.87 a |
| Rubber (n = 12) | 68.5 ± 5.1 | 0.8 ± 2.6 | 30.7 ± 3.7 | Sandy clay loam | 338.9 ± 70.03 a | 34.7 ± 9.08 a | 186.6 ± 78.2 b | 13.8 ± 4.18 a | 0.31 ± 0.08 ab | 69.4 ± 8.28 b | 8.52 ± 1.13 b | 0.66 ± 0.21 b | 1.69 ± 0.06 bc | 19.81 ± 2.16 a |
| Arecanut (n = 4) | 27.4 ± 6.2 | 38.1 ± 1.5 | 34.5 ± 11.2 | Clay loam | 333.4 ± 44.9 a | 34.7 ± 2.36 a | 380.4 ± 42.8 a | 13.5 ± 1.8 a | 0.24 ± 0.08 b | 92.8 ± 8.28 ab | 15.46 ± 2.6 ab | 1.61 ± 0.19 a | 0.56 ± 0.06 bc | 11.76 ± 1.81 a |
| Sal forest (n = 6) | 56.8 ± 8.9 | 17.2 ± 3.2 | 26.0 ± 2.5 | Sandy clay loam | 309.5 ± 20.99 a | 30.7 ± 9.57 a | 211.5 ± 35.1 b | 14.8 ± 2.59 a | 0.37 ± 0.08 a | 73.5 ± 9.94 ab | 6.29 ± 1.17 b | 0.52 ± 0.11 b | 0.42 ± 0.06 c | 14.83 ± 1.08 a |
| Scenario | Sand (%) | Silt (%) | Clay (%) | Soil Texture | N (Kg ha−1) | P (Kg ha−1) | K (Kg ha−1) | S (ppm) | B (ppm) | Fe (ppm) | Mn (ppm) | Zn (ppm) | Cu (ppm) | SMC (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Paddy (n = 8) | 28.0 ± 4.3 | 19.9 ± 1.2 | 52.1 ± 5.2 | Clay | 228.2 ± 13.34 a | 40.9 ± 4.14 a | 90.38 ± 11.8 c | 10.5 ± 1.6 a | 0.23 ± 0.05 a | 57.9 ± 3.16 a | 17.44 ± 2.2 a | 0.40 ± 0.05 b | 0.64 ± 0.06 a | 28.14 ± 1.9 a |
| Banana (n = 4) | 25.0 ± 5.7 | 38.0 ± 1.1 | 37.0 ± 6.2 | Clay loam | 231.75 ± 22.74 a | 47.89 ± 5.27 a | 229.3 ± 16.2 abc | 8.3 ± 1.1 a | 0.17 ± 0.03 a | 57.8 ± 4.76 a | 6.15 ± 2.7 bc | 0.32 ± 0.18 b | 0.84 ± 0.07 a | 20.7 ± 2.73 ab |
| Rubber (n = 12) | 9.0 ± 5.6 | 49.5 ± 4.8 | 41.5 ± 3.9 | Silty clay | 217.62 ± 25.5 a | 26.9 ± 7.43 a | 173.8 ± 12.4 bc | 10.7 ± 1.75 a | 0.26 ± 0.09 a | 41.5 ± 6.2 a | 4.93 ± 1.23 bc | 0.37 ± 0.17 b | 0.27 ± 0.24 b | 23.08 ± 2.24 ab |
| Arecanut (n = 4) | 13.9 ± 1.2 | 37.7 ± 3.3 | 48.4 ± 4.5 | Clay | 222.23 ± 11.2 a | 30.7 ± 2.17 a | 359.2 ± 13.2 a | 13.5 ± 1.5 a | 0.19 ± 0.08 a | 63.6 ± 8.28 a | 10.89 ± 2.1 ab | 0.83 ± 0.18 a | 0.31 ± 0.08 b | 17.64 ± 3.12 b |
| Sal forest (n = 6) | 15.4 ± 2.6 | 42.3 ± 3.4 | 42.3 ± 4.6 | Silty clay | 235.5 ± 10.12 a | 25.1 ± 6.78 a | 268.2 ± 20.3 ab | 10.5 ± 1.62 a | 0.27 ± 0.07 a | 51.5 ± 7.87 a | 1.61 ± 0.4 c | 0.28 ± 0.09 b | 0.20 ± 0.03 b | 20.13 ± 2.37 ab |
| Scenario | Bacteria (10 6 CFU/g Soil) | Fungi (10 4 CFU/g Soil) | Actinomycetes (10 4 CFU/g Soil) | |||
|---|---|---|---|---|---|---|
| 0–25 cm | 25–50 cm | 0–25 cm | 25–50 cm | 0–25 cm | 25–50 cm | |
| Paddy (n = 8) | 362.5 ± 17.4 b | 297.4 ± 21.0 b | 80 ± 6.1 b | 32.5 ± 3.04 a | 1129.6 ± 93.4 b | 722.5 ± 74.34 b |
| Banana (n = 4) | 135 ± 14.5 b | 135 ± 14.5 b | 75 ± 6.6 b | 65.0 ± 3.6 a | 195 ± 35.4 c | 157 ± 4.24 c |
| Rubber (n = 12) | 784.3 ± 31.7 a | 485.6 ± 40.9 ab | 142.5 ± 9.8 b | 71.2 ± 7.7 a | 1216.7 ± 109.7 b | 1292.5 ± 86.2 b |
| Arecanut (n = 4) | 400 ± 19.7 b | 290 ± 42.4 b | 510.6 ± 21.1 a | 80.0 ± 2.4 a | 2006 ± 71.6 a | 1566 ± 51.6 b |
| Sal forest (n = 6) | 760 ± 47.6 a | 654 ± 32.5 a | 586.6 ± 25.1 a | 70.0 ± 6.05 a | 1399.3 ± 98.3 b | 2766.3 ± 42.9 a |
| Soil pH | SOC | N | P | K | S | B | Fe | Mn | Zn | Cu | SMC | VLSOC | Bacteria | Fungi | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Soil pH | 1.000 | ||||||||||||||
| SOC | −0.355 * | 1.000 | |||||||||||||
| N | −0.340 * | 0.608 ** | 1.000 | ||||||||||||
| P | 0.095 | 0.013 | 0.064 | 1.000 | |||||||||||
| K | 0.274 | −0.023 | 0.144 | 0.249 | 1.000 | ||||||||||
| S | −0.103 | −0.080 | 0.031 | 0.450 ** | 0.172 | 1.000 | |||||||||
| B | −0.411 ** | −0.048 | 0.166 | 0.018 | −0.267 | 0.350 * | 1.000 | ||||||||
| Fe | 0.183 | 0.430 ** | 0.317 * | 0.137 | 0.049 | −0.074 | −0.006 | 1.000 | |||||||
| Mn | 0.184 | 0.271 | 0.167 | 0.100 | −0.205 | 0.117 | 0.039 | 0.332 * | 1.000 | ||||||
| Zn | 0.129 | 0.314 * | 0.125 | 0.203 | 0.336 * | 0.249 | −0.094 | 0.484 ** | 0.242 | 1.000 | |||||
| Cu | −0.092 | 0.237 | 0.121 | 0.211 | −0.435 ** | 0.042 | 0.058 | 0.365 * | 0.739 ** | 0.021 | 1.000 | ||||
| SMC | −0.093 | 0.407 ** | −0.085 | −0.032 | −0.248 | −0.373 * | −0.227 | −0.015 | 0.031 | −0.078 | 0.088 | 1.000 | |||
| VLSOC | −0.344 * | 0.330 * | 0.376 * | 0.148 | 0.164 | 0.350 * | 0.371 * | 0.001 | −0.044 | 0.014 | −0.117 | 0.016 | 1.000 | ||
| Bacteria | 0.199 | 0.114 | −0.002 | −0.021 | −0.007 | 0.111 | 0.299 * | 0.124 | −0.044 | −0.027 | −0.140 | 0.105 | 0.373 * | 1.000 | |
| Fungi | −0.243 | 0.236 | 0.297 * | −0.262 | 0.079 | 0.100 | 0.360 * | 0.245 | −0.095 | 0.263 | −0.213 | −0.397 ** | 0.344 * | 0.340 * | 1.000 |
| Actinomycetes | −0.158 | 0.146 | 0.065 | −0.470 ** | 0.161 | −0.173 | 0.086 | −0.245 | −0.266 | 0.048 | −0.430 ** | 0.103 | 0.149 | 0.201 | 0.154 |
| Total Variance Explained | ||||||
|---|---|---|---|---|---|---|
| Component | Initial Eigenvalues | Extraction Sums of Squared Loadings | ||||
| Total | % of Variance | Cumulative % | Total | % of Variance | Cumulative % | |
| 1 | 2.986 | 18.660 | 18.660 | 2.986 | 18.660 | 18.660 |
| 2 | 2.684 | 16.773 | 35.433 | 2.684 | 16.773 | 35.433 |
| 3 | 2.153 | 13.458 | 48.891 | 2.153 | 13.458 | 48.891 |
| 4 | 1.907 | 11.920 | 60.811 | 1.907 | 11.920 | 60.811 |
| 5 | 1.401 | 8.757 | 69.568 | 1.401 | 8.757 | 69.568 |
| 6 | 1.279 | 7.991 | 77.559 | 1.279 | 7.991 | 77.559 |
| Component Matrix | ||||||
|---|---|---|---|---|---|---|
| Component | ||||||
| 1 | 2 | 3 | 4 | 5 | 6 | |
| pH | −0.377 | 0.309 | 0.466 | 0.383 | −0.344 | 0.384 |
| SOC | 0.696 | 0.122 | −0.436 | 0.383 | 0.245 | 0.021 |
| N | 0.698 | −0.049 | −0.164 | 0.153 | 0.203 | −0.303 |
| P | 0.209 | 0.374 | 0.538 | −0.188 | 0.515 | 0.211 |
| K | 0.057 | −0.240 | 0.629 | 0.514 | 0.288 | −0.077 |
| S | 0.367 | −0.056 | 0.598 | −0.453 | 0.190 | 0.077 |
| B | 0.441 | −0.332 | −0.033 | −0.628 | −0.226 | 0.080 |
| Fe | 0.543 | 0.436 | 0.072 | 0.379 | −0.320 | 0.016 |
| Mn | 0.363 | 0.692 | −0.069 | −0.084 | −0.237 | 0.064 |
| Zn | 0.446 | 0.183 | 0.383 | 0.481 | −0.076 | −0.105 |
| Cu | 0.294 | 0.781 | −0.239 | −0.314 | −0.109 | −0.020 |
| SMC | −0.093 | 0.191 | −0.612 | 0.255 | 0.394 | 0.483 |
| VLSOC | 0.614 | −0.414 | 0.026 | −0.154 | 0.298 | 0.289 |
| Bacteria | 0.292 | −0.328 | 0.029 | 0.028 | −0.314 | 0.781 |
| Fungi | 0.559 | −0.480 | 0.075 | 0.070 | −0.475 | −0.179 |
| Actinomycetes | −0.009 | −0.634 | −0.283 | 0.341 | −0.028 | 0.034 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hota, S.; Mishra, V.; Mourya, K.K.; Giri, K.; Kumar, D.; Jha, P.K.; Saikia, U.S.; Prasad, P.V.V.; Ray, S.K. Land Use, Landform, and Soil Management as Determinants of Soil Physicochemical Properties and Microbial Abundance of Lower Brahmaputra Valley, India. Sustainability 2022, 14, 2241. https://doi.org/10.3390/su14042241
Hota S, Mishra V, Mourya KK, Giri K, Kumar D, Jha PK, Saikia US, Prasad PVV, Ray SK. Land Use, Landform, and Soil Management as Determinants of Soil Physicochemical Properties and Microbial Abundance of Lower Brahmaputra Valley, India. Sustainability. 2022; 14(4):2241. https://doi.org/10.3390/su14042241
Chicago/Turabian StyleHota, Surabhi, Vidyanand Mishra, Krishna Kumar Mourya, Krishna Giri, Dinesh Kumar, Prakash Kumar Jha, Uday Shankar Saikia, P. V. Vara Prasad, and Sanjay Kumar Ray. 2022. "Land Use, Landform, and Soil Management as Determinants of Soil Physicochemical Properties and Microbial Abundance of Lower Brahmaputra Valley, India" Sustainability 14, no. 4: 2241. https://doi.org/10.3390/su14042241
APA StyleHota, S., Mishra, V., Mourya, K. K., Giri, K., Kumar, D., Jha, P. K., Saikia, U. S., Prasad, P. V. V., & Ray, S. K. (2022). Land Use, Landform, and Soil Management as Determinants of Soil Physicochemical Properties and Microbial Abundance of Lower Brahmaputra Valley, India. Sustainability, 14(4), 2241. https://doi.org/10.3390/su14042241