
Composition Characteristics of an Urban Forest Soil Seed Bank and Its Influence on Vegetation Restoration: A Case Study in Dadu Terrace, Central Taiwan


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Setting of Sampling Plot and Vegetation Survey
2.3. Sampling and Germination Test of the Soil Seed Bank
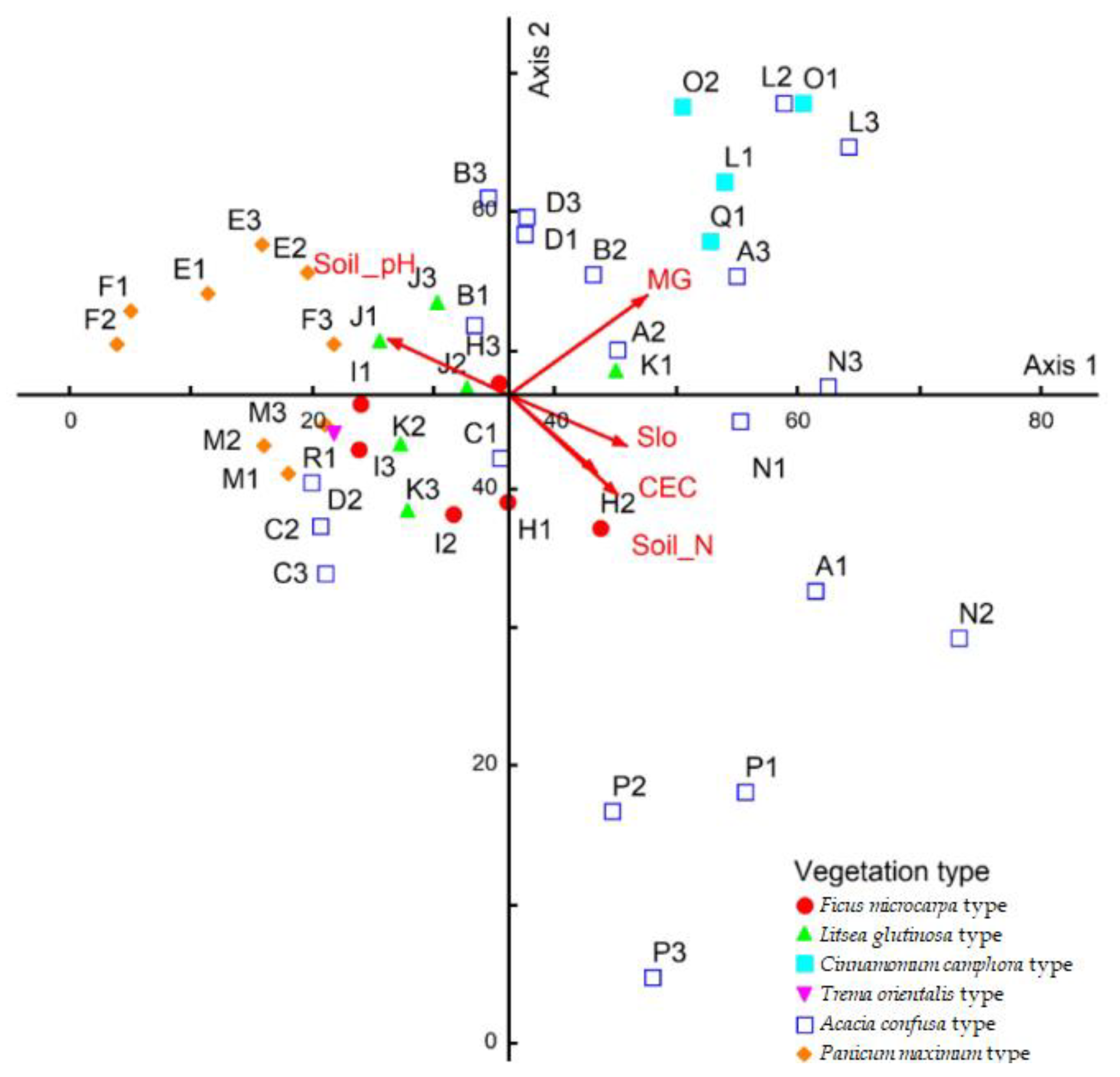
2.4. Measurement of Environmental Factors
2.5. Data Analysis and Statistical Methods
3. Results
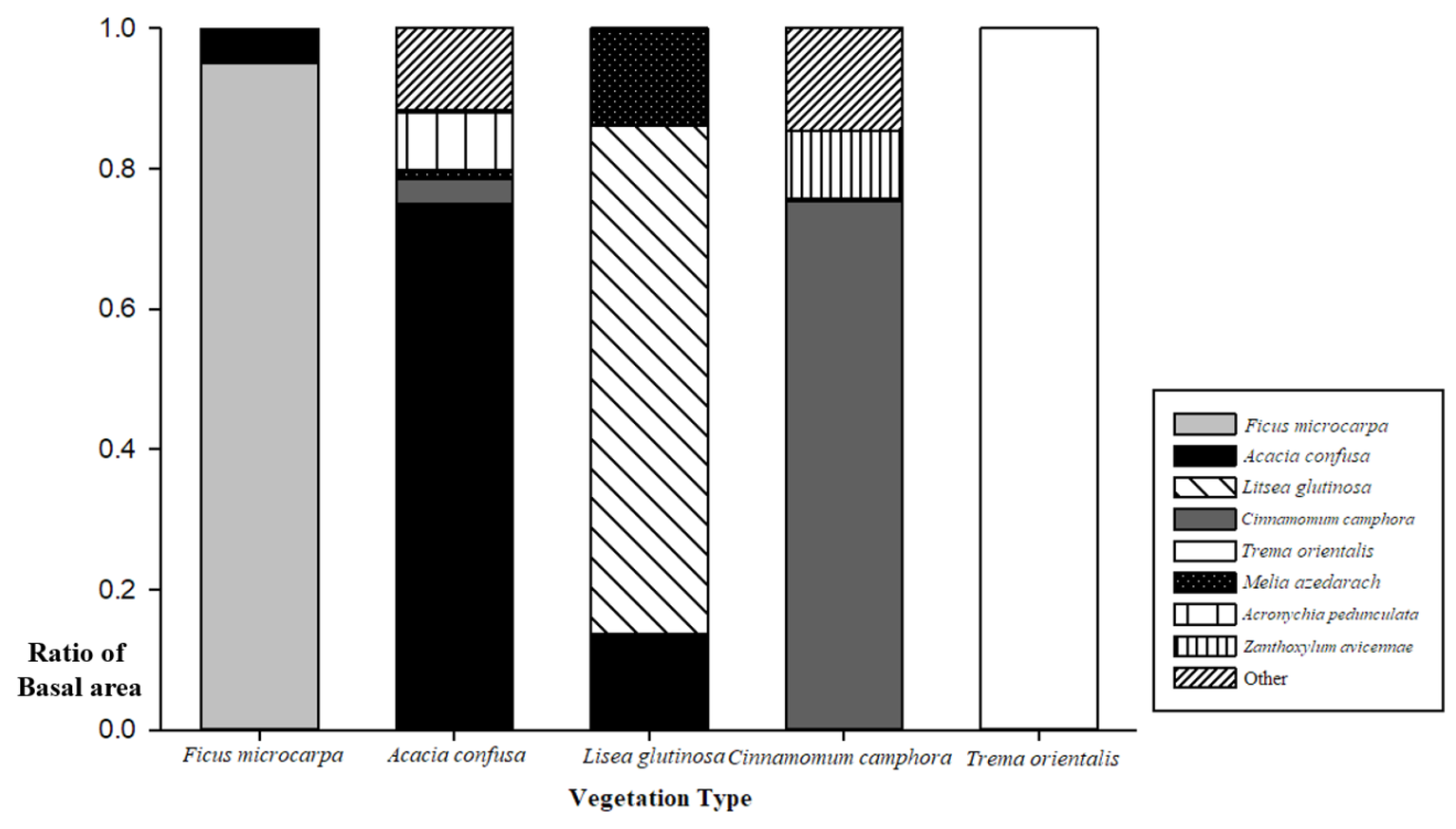
3.1. Composition of Aboveground Plant Community
3.2. Composition of the Soil Seed Bank
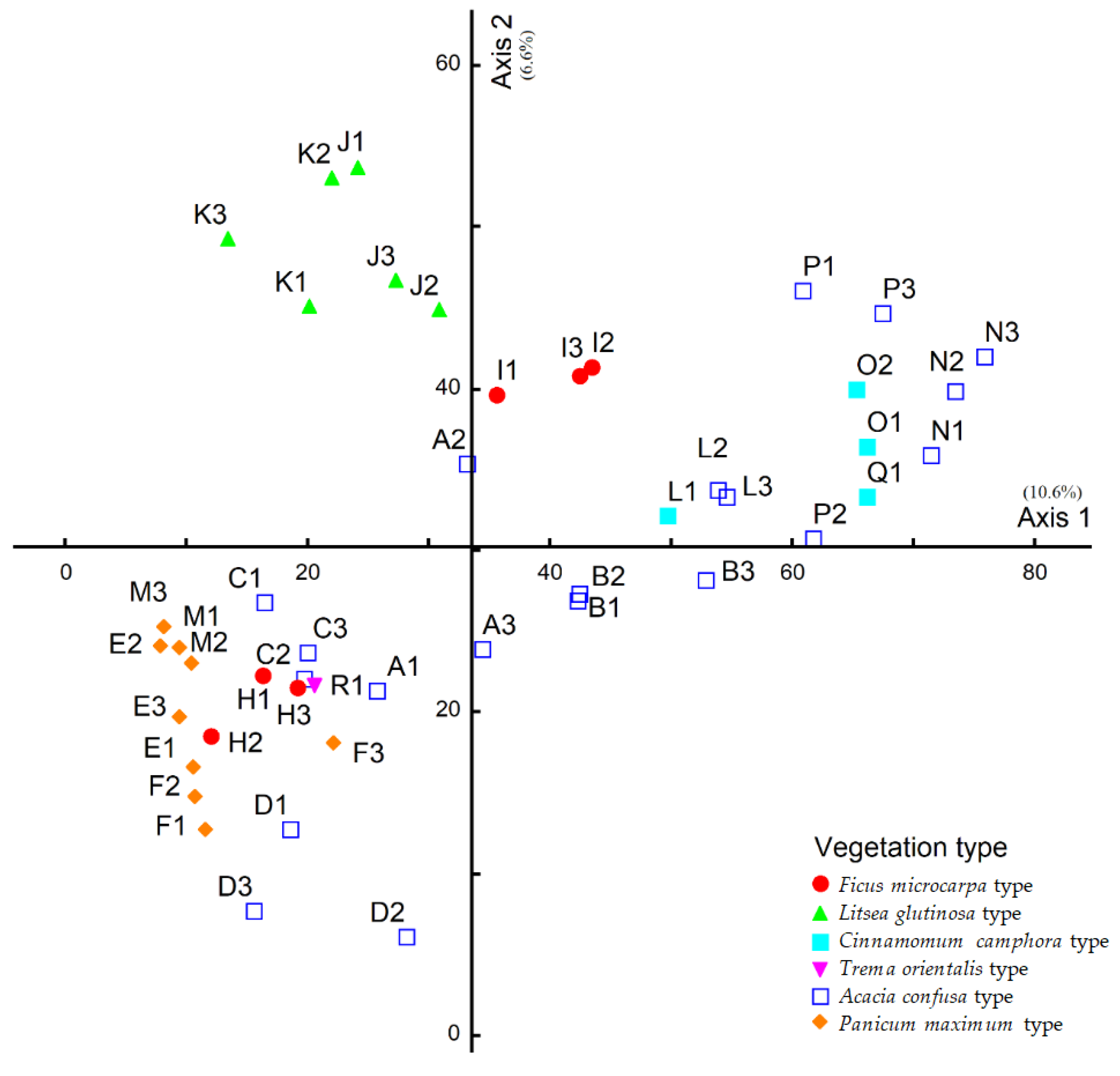
3.3. Similarities between Soil Seed Bank and Aboveground Plant Composition
4. Discussion
4.1. Composition Characteristics of Aboveground Vegetation and Soil Seed Bank
4.2. Feasibility of Soil Seed Banks for Vegetation Restoration
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clark, C.J.; Poulsen, J.R.; Levey, D.J.; Osenberg, C.W. Are plant populations seed limited? A critique and meta-analysis of seed addition experiments. Am. Nat. 2007, 170, 128–142. [Google Scholar] [CrossRef]
- Paul, M.; Catterall, C.P.; Kanowski, J.; Pollard, P.C. Recovery of rain forest soil seed banks under different reforestation pathways in eastern Australia. Ecol. Manag. Restor. 2012, 13, 144–152. [Google Scholar] [CrossRef]
- Simpson, L.R.; Leck, M.A.; Parker, V.T. Ecology of Soil Seed Banks; Academic Press, Inc.: San Diego, CA, USA, 1989; pp. 5–7. [Google Scholar]
- Fenner, M.; Thompson, K. The Ecology of Seed; Cambridge University Press: New York, NY, USA, 2006; pp. 67–92. [Google Scholar]
- Li, K.F.; Wang, W.; Tzeng, H.Y.; Tseng, S.H.; Kuo, C.C.; Lin, C.Y. Composition of the soil seed bank and its effects on forest restoration in Beikeng, Zhukeng Area of Dadu Terrace, Taichung. J. Taiwan Agr. Eng. 2018, 64, 30–48, (In Chinese with English Summary). [Google Scholar]
- Garwood, N.C. Seed germination in a seasonal tropical forest in Panama: A community study. Ecol. Monogr. 1984, 53, 159–181. [Google Scholar] [CrossRef]
- Tzeng, H.Y.; Lan, H.; Wang, W.; Hsu, C.K.; Tseng, Y.H.; Kuo, C.C. Composition and spatial distribution of the soil seed bank at Taiwania plantation in Lienhuachi research center, central Taiwan. Q. J. Chin. For. 2018, 51, 309–329, (In Chinese with English Summary). [Google Scholar]
- Zhang, Z.Q.; Shu, W.S.; Lan, C.Y.; Huang, M.H. The revegetation of a Lead/Zinc mine tailings site with an introduced soil seed bank. Chin. J. Plant Ecol. 2000, 24, 601–607, (In Chinese with English Summary). [Google Scholar]
- Huang, Y.; Ren, H.; Wang, J.; Liu, N.; Jian, S.; Cai, H.; Hui, D.; Guo, Q. Effects of Wollastonia biflora expansion on the soil seed bank in native forest communities on a tropical coral island. Glob. Ecol. Conserv. 2021, 25, e01403. [Google Scholar] [CrossRef]
- Waryszak, P.; Standish, R.J.; Ladd, P.G.; Enright, N.J.; Brundrett, M.; Fontaine, J.B. Best served deep: The seedbank from salvaged topsoil underscores the role of the dispersal filter in restoration practice. Appl. Veg. Sci. 2021, 24, e125392021. [Google Scholar] [CrossRef]
- Wu, T.H.; Tsai, S.T.; Chiu, C.A.; Wang, W.; Tzeng, H.Y.; Lu, K.C. Soil seed bank composition of Hui-Sun Experimental Forest Station. Q. J. For. Res. 2014, 36, 85–100, (In Chinese with English Summary). [Google Scholar]
- Chang, J.L.; Wang, W.; Lin, C.Y.; Tseng, Y.H.; Chiu, C.A.; Lin, C.C.; Tzeng, H.Y. Comparison of soil seed bank compositions between patch cutting of a Chinese fir plantation and nearby forests in HuiSun Forest Station. Q. J. For. Res. 2019, 41, 177–190, (In Chinese with English Summary). [Google Scholar]
- Kuo, C.Y.; Tseng, Y.H.; Tzeng, H.Y. Comparison of soil seed bank in Tungshih Forest Station of National Chung Hsing University. Q. J. For. Res. 2020, 42, 73–90, (In Chinese with English Summary). [Google Scholar]
- Ludewig, K.; Hansen, W.; Klinger, Y.P.; Eckstein, R.L.; Otte, A. Seed bank offers potential for active restoration of mountain meadows. Restor. Ecol. 2021, 29, e13311. [Google Scholar] [CrossRef]
- Valkó, O.; Deák, B.; Török, P.; Tóth, K.; Kiss, R.; Kelemen, A.; Tóthmérész, B. Dynamics in vegetation and seed bank composition highlight the importance of post-restoration management in sown grasslands. Restor. Ecol. 2021, 29, e13192. [Google Scholar] [CrossRef]
- Lamb, N.; Havens, K.; Holloway, J.; Steffen, J.F.; Zeldin, J.; Kramer, A.T. Low passive restoration potential following invasive woody species removal in oak woodlands. Restor. Ecol. 2021, 29, e13568. [Google Scholar] [CrossRef]
- Wang, N.; He, X.; Zhao, F.; Wang, D.; Jiao, J. Soil seed bank in different vegetation types in the Loess Plateau region and its role in vegetation restoration. Restor. Ecol. 2020, 28, A5–A12. [Google Scholar] [CrossRef]
- Liao, C.C.; Lu, F.Y.; Ou, C.H. The study of vegetational ecology and flora analysis in the area of Toukoshan. Bull. Exper. For. Nat. Chung Hsing Univ. 1987, 8, 43–65, (In Chinese with English Summary). [Google Scholar]
- Wang, C.M.; Chiu, C.A.; Tseng, Y.H.; Tzeng, H.Y.; Lu, K.C. Vegetation ecology at Dakeng Area, Taichung. Q. J. For. Res. 2010, 32, 7–22. [Google Scholar]
- Aganga, A.A.; Tshwenyane, S. Potentials of guinea grass (Panicum maximum) as forage crop in livestock production. Pak. J. Nutr. 2004, 3, 1–4. [Google Scholar]
- Williams, D.G.; Baruch, Z. African grass invasion in the Americas: Ecosystem consequences and the role of ecophysiology. Biol. Invasions 2000, 2, 123–140. [Google Scholar] [CrossRef]
- Portela, R.C.Q.; Matos, D.M.S.; Siqueira, L.P.D.; Braz, M.I.G.; Silva-Lima, L.; Marrs, R.H. Variation in aboveground biomass and necromass of two invasive species in the Atlantic rainforest, southeast Brazil. Acta Bot. Bras. 2009, 23, 571–577. [Google Scholar] [CrossRef]
- Hsu, C.C. Taiwan Grasses; Taiwan Provincial Education Association: Taipei, Taiwan, 1975; 884p. [Google Scholar]
- Ho, C.Y.; Tsai, M.Y.; Huang, Y.L.; Kao, W.Y. Ecophysiological factors contributing to the invasion of Panicum maximum into native Miscanthus sinensis grassland in Taiwan. Weed Res. 2016, 56, 69–77. [Google Scholar] [CrossRef]
- Chang, C.Y.; Tzeng, H.Y.; Lu, K.C.; Tseng, Y.H. Invasiveness assessment system of naturalized plants in Taiwan. Q. J. For. Res. 2008, 30, 29–40, (In Chinese with English Summary). [Google Scholar]
- Chiu, C.A.; Wang, C.C.; Lu, K.C.; Lin, P.H.; Tzeng, H.Y. Examination of semi-arid regions and potential savanna vegetation in Taiwan. Taiwan J. For. Sci. 2008, 23, S23–S36, (In Chinese with English Summary). [Google Scholar]
- Ho, C.S. An Introduction to the Geology of Taiwan-Explanatory Text of the Geologic Map of Taiwan; Central Geological Survey, MOEA: New Taipei, Taiwan, 1986; pp. 100–104. [Google Scholar]
- Forestry Bureau. The Fourth Forest Resource Survey Report; Forestry Bureau: Taipei, Taiwan, 2020; 79p.
- Chiou, C.R.; Hsueh, I.C.; Liu, Y.A.; Lai, Y.J. Trend analysis of land-use and land-cover changes in Dadu Hill, Taichung. City Plan. 2012, 39, 25–50, (In Chinese with English Summary). [Google Scholar]
- Tang, K.C.; Tseng, Y.H.; Tzeng, H.Y.; Lu, K.C. Soil seed bank composition of secondary forest in Dakeng Area, Taichung. Q. J. For. Res. 2011, 33, 35–48, (In Chinese with English Summary). [Google Scholar]
- Chang, C.H.; Chang, C.Y. A study on the relationship between the grass fire and the plants in the Dadu Mountain Area. Q. J. For. Res. 2004, 26, 1–10, (In Chinese with English Summary). [Google Scholar]
- Chuang, Y.L.; Chompuchan, C.; Lin, C.Y. Modeling for wildfire risk evaluation in Dadu Terrace. J. Soil Water Conser. 2016, 48, 1681–1696, (In Chinese with English Summary). [Google Scholar]
- Ri, X.; Yang, J.; Zhao, L.Q.; Qing, H.; Latanzhula, A.; Yao, Z.Y.; Chang, G. Establishment, development, and decline of Salix psammophila communities: Changes in soil conditions and floristic composition in dune slacks. Glob. Ecol. Conserv. 2020, 22, e00967. [Google Scholar] [CrossRef]
- Mwendwa, B.A.; Kilawe, C.J.; Treydte, A.C. Effect of seasonality and light levels on seed germination of the invasive tree Maesopsis eminii in Amani Nature Forest Reserve, Tanzania. Glob. Ecol. Conserv. 2020, 21, e00807. [Google Scholar] [CrossRef]
- Su, H.J. Forest Habitat Factors and Their Quantitative Assessment. Q. J. Chin. For. 1985, 20, 1–14, (In Chinese with English Summary). [Google Scholar]
- Hsia, Y.J.; Wang, W.S. Calculation of Potential Solar Irradiance on Slopes; Research Note 001; Forest Research Institute: Taipei, Taiwan, 1985; p. 28, (In Chinese with English Summary). [Google Scholar]
- Day, F.P.; Monk, C.D. Vegetation patterns on a southern Appalachian watershed. Ecology 1974, 55, 1064–1074. [Google Scholar] [CrossRef] [Green Version]
- McLean, E.O. Soil pH and lime requirement. Method Soil. Anal. 1983, 9, 199–224. [Google Scholar]
- MacDonald, C.C. Methods of Soil and Tissue Analysis Used in the Analytical Laboratory; Maritimes Forest Research Centre, Canadian Forestry Service, Department of the Environment; Centre; Fredericton, NB, Canada, 1972; 127p. [Google Scholar]
- Avramidis, P.; Nikolaou, K.; Bekiari, V. Total organic carbon and total nitrogen in sediments and soils: A comparison of the wet oxidation–titration method with the combustion-infrared method. Agric. Sci. Proc. 2015, 4, 425–430. [Google Scholar] [CrossRef] [Green Version]
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Methods of Soil Analysis Part II. Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy; Soil Science Society of America: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Rhoades, J.D. Chemical and microbiological properties. In Cation Exchange Capacity; American Society of Agronomy: Madison, WI, USA, 1982; pp. 149–157. [Google Scholar]
- Editorial Committee of the Flora of Taiwan. Flora of Taiwan Second Edition; Department of Botany, National Taiwan University: Taipei, Taiwan, 2003; Volumes 1–6. [Google Scholar]
- Editorial Committee of the Red List of Vascular Plants of Taiwan. The Red List of Vascular Plants of Taiwan, 2017; Endemic Species Research Institute, Forestry Bureau, Council of Agriculture, Executive Yuan and Taiwan Society of Plant Systematics: Nantou, Taiwan, 2017; p. 187. [Google Scholar]
- Wu, S.H.; Hsieh, C.F.; Rejmánek, M. Catalogue of the naturalized flora of Taiwan. Taiwania 2004, 49, 16–31. [Google Scholar]
- Chen, S.H. Naturalized Plants of Eastern Taiwan; National Hualien University of Education: Hualien, Taiwan, 2008; p. 422. (In Chinese) [Google Scholar]
- Shiu, H.J.; Lee, P.F. Influence of gradient length on community ordinations. Taiwan J. For. Sci. 2003, 18, 201–211, (In Chinese with English Summary). [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD Multivariate Analysis of Ecological Data. Ver. 6.08; MjM Software: Gleneden Beach, OR, USA, 2011. [Google Scholar]
- Hyatt, L.A. Differences between seed bank composition and field recruitment in a temperate zone deciduous forest. Am. Midl. Nat. 1999, 142, 31–38. [Google Scholar] [CrossRef]
- IBM Corp. IBM SPSS Statistics for Windows, Version 22.0. Armonk; IBM Corp: New York, NY, USA, 2013. [Google Scholar]
- Sørensen, T. A Method of Establishing Groups of Equal Amplitude in Plant Sociology Based on Similarity of Species and Its Application to Analyses of the Vegetation on Danish Commons. Biol. Skar. 1948, 5, 1–34. [Google Scholar]
- Chang, N.H.; Ma, F.C.; Yu, H.M.; Hsui, Y.R. Dynamics of soil seed bank and seedlings in the Fushan broadleaf forest. Taiwan J. For. Sci. 1998, 13, 279–289, (In Chinese with English Summary). [Google Scholar]
- Gioria, M.; Osborne, B.A. Similarities in the impact of three large invasive plant species on soil seed bank communities. Biol. Invasions 2010, 12, 1671–1683. [Google Scholar] [CrossRef]
- Gioria, M.; Pyšek, P.; Moravcová, L. Soil seed banks in plant invasions: Promoting species invasiveness and long-term impact on plant community dynamics. Preslia 2012, 84, 327–350. [Google Scholar]
- Gioria, M.; Pyšek, P. The legacy of plant invasions: Changes in the soil seed bank of invaded plant communities. BioScience 2016, 66, 40–53. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.M.A.; Loneragan, W.A.; Grant, C.D.; Koch, J.M. Effect of fire on the topsoil seed banks of rehabilitated bauxite mine sites in the Jarrah forest of Western Australia. Ecol. Manag. Restor. 2000, 1, 50–60. [Google Scholar] [CrossRef]
- O’Loughlin, L.S.; Green, P.T.; Morgan, J.W. What potential is there for regeneration of native species from the soil seed bank in coast tea tree-dominated scrub? Ecol. Manag. Restor. 2014, 15, 80–83. [Google Scholar] [CrossRef]
- Kohagura, T.D.C.; Souza, E.B.D.; Bao, F.; Ferreira, F.A.; Pott, A. Flood and fire affect the soil seed bank of riparian forest in the Pantanal wetland. Rodriguésia 2020, 71. [Google Scholar] [CrossRef] [Green Version]
- Zida, D.; Sanou, L.; Diawara, S.; Savadogo, P.; Thiombiano, A. Herbaceous seeds dominates the soil seed bank after long-term prescribed fire, grazing and selective tree cutting in savanna-woodlands of West Africa. Acta Oecol. 2020, 108, 103607. [Google Scholar] [CrossRef]
- González-Muñoz, N.; Costa-Tenorio, M.; Espigares, T. Invasion of alien Acacia dealbata on Spanish Quercus robur forests: Impact on soils and vegetation. For. Ecol. Manag. 2012, 269, 214–221. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Huenneke, L.F. Disturbance, diversity, and invasion: Implications for conservation. Conserv. Biol. 1992, 6, 324–337. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, R.J. Invasive Species in a Changing World; Island Press: Washington, DC, USA, 2000; pp. 65–93. [Google Scholar]
- Ehrenfeld, J.G. Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems 2003, 6, 503–523. [Google Scholar] [CrossRef]
- Cheng, C.H.; Chen, Y.S.; Huang, Y.H.; Chiou, C.R.; Lin, C.C.; Menyailo, O.V. Effects of repeated fires on ecosystem C and N stocks along a fire induced forest/grassland gradient. J. Geophys. Res. Biogeosci. 2013, 118, 215–225. [Google Scholar] [CrossRef]
- D’antonio, C.M.; Dudley, T.I.; Mack, M. Disturbance and biological invasions: Direct effects and feedbacks. In Ecosystems of Disturbed Ground; Walker, L.R., Ed.; Elsevier: New York, NY, USA, 1999; pp. 413–452. [Google Scholar]
- Fusco, E.J.; Finn, J.T.; Balch, J.K.; Nagy, R.C.; Bradley, B.A. Invasive grasses increase fire occurrence and frequency across US ecoregions. Proc. Natl. Acad. Sci. USA 2019, 116, 23594–23599. [Google Scholar] [CrossRef] [PubMed]
- María, U.; Emilio, M.B.; Paulo, R.; Marina, A.; Inge, J. Effects of forest fragmentation on the seedling recruitment of a tropical herb: Assessing seed vs. safe-site limitation. Ecology 2010, 91, 1317–1328. [Google Scholar]
- Chang, N.H. Effects of light on seed germination of three pioneer tree species. Taiwan J. For. Sci. 1996, 11, 195–199, (In Chinese with English Summary). [Google Scholar]
- Hsu, P.C. The study of natural succession of the man-made forest of Acacia confusa in Hwa-Lin Experimental Forest of Chinese Culture University. Hwa Kang J. Agric. 2003, 11, 85–106. [Google Scholar]
- Wang, Z.J.; Cao, M.; Li, G.F.; Men, L.; Duo, G.; Zha, T.; Zong, W. Trema orientalis seeds dispersed by birds and its ecological role. Zool. Res. 2002, 23, 214–219. [Google Scholar]
- Sheu, B.H.; Chang, C.T. Experiment on hastening germination and storage of India-charcoal Trema (II). Bull. Exp. For. Natl. Chung Hsing Univ. 1981, 3, 41–62. [Google Scholar]
- Lu, F.Y.; Ou, C.H.; Liao, C.C. The succession of India-charcoal Trema vegetation on the alluvium Lau-Dau Brook, Huey-Suen Forest Station. Bull. Exp. For. Natl. Chung Hsing Univ. 1984, 5, 11–24. [Google Scholar]
- Tang, Y.; Cao, M.; Zhang, J.H.; Ren, Y.H. The impact of slash-and -burn agriculture on the soil seed bank of Trema orientalis forest. Acta Bot. Yunnanica 1997, 19, 423–428. [Google Scholar]
- Goodale, U.M.; Ashton, M.S.; Berlyn, G.P.; Gregoire, T.G.; Singhakumara, B.M.P.; Tennakoon, K.U. Disturbance and tropical pioneer species: Patterns of association across life history stages. For. Ecol. Manag. 2012, 227, 54–66. [Google Scholar] [CrossRef]
- Wang, W.; Chiu, C.A.; Tsai, S.T.; Kuo, C.C.; Tseng, Y.H.; Tzeng, H.Y. Vegetation classification of Huoyanshan Area, Miaoli, Taiwan. Q. J. Chin. For. 2016, 49, 131–149. [Google Scholar]
- Rodrigues, C.R.; Rodrigues, B.F. Enhancement of seed germination in Trema orientalis (L.) Blume—Potential plant species in revegetation of mine wastelands. J. Sustain. For. 2014, 3, 46–58. [Google Scholar] [CrossRef]
- Chiu, C.A.; Hsu, H.S.; Lin, S.H.; Chen, W.C. The trajectory and goal of landslide revegetation: A proposal from the viewpoints of restoration ecology. J. Chin. Soil Water Conserv. 2016, 47, 104–110. [Google Scholar]
- Goebel, P.C.; Wyse, T.C.; Corace, R.G., III. Determining reference ecosystem conditions for disturbed landscapes within the context of contemporary resource management issues. J. For. 2005, 103, 351–356. [Google Scholar]
- Miyawaki, A. Restoration of living environment based on vegetation ecology: Theory and practice. Ecol. Res. 2004, 19, 83–90. [Google Scholar] [CrossRef]
- Chiu, C.A.; Tzeng, H.Y.; Wang, C.M.; Wu, Y.H.; Tzeng, Y.H. Survey of plant resources and proposition of ecological restoration in Tungshih Forest Station. Q. J. For. Res. 2012, 34, 13–38, (In Chinese with English Summary). [Google Scholar]
- Chiu, C.A. A preliminary exploration of restoration ecology. Q J. Chin. For. 2012, 45, 289–297, (In Chinese with English Summary). [Google Scholar]
- Lu, Y.; Ranjitkar, S.; Harrison, R.D.; Xu, J.; Ou, X.; Ma, X.; He, J. Selection of native tree species for subtropical forest restoration in southwest China. PLoS ONE 2017, 12, e0170418. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, J.C.; Sabogal, C. Restoring degraded forest land with native tree species: The experience of “Bosques Amazónicos” in Ucayali, Peru. Forests 2019, 10, 851. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Q.; Shu, W.S.; Lan, C.Y.; Wong, M.H. Soil seed bank as an input of seed source in revegetation of Lead/Zinc mine tailings. Restor. Ecol. 2001, 9, 378–385, (In Chinese with English Summary). [Google Scholar] [CrossRef]
- Van Etten, E.J.; Neasham, B.; Dalgleish, S. Soil seed banks of fringing salt lake vegetation in arid Western Australia–density, composition and implications for postmine restoration using topsoil. Ecol. Manag. Restor. 2014, 15, 239–242. [Google Scholar] [CrossRef] [Green Version]
- Taiwo, D.M.; Oyelowo, O.J.; Ogedengbe, T.C.; Woghiren, A.I. The role of soil seed bank in forest regeneration. Asian J. Res. Agric. For. 2018, 1, 1–10. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Type | Grassland | Forest | ||||
|---|---|---|---|---|---|---|
| Panicum maximum Type (n = 9) | Acacia confusa Type (n = 20) | Ficus microcarpa Type (n = 6) | Litsea glutinosa Type (n = 6) | Cinnamomum camphora Type (n = 4) | Trema orientalis Type (n = 1) | |
| Species number of aboveground vegetation | 11.4± 3.8 b | 23.6 ± 11.2 a | 14.5 ± 10.0 ab | 17.2 ± 7.7 ab | 35.0 ± 6.7 a | 16.0 |
| Percentage of naturalized species of aboveground vegetation (%) | 56.7 ± 11.6 a | 26.3 ± 16.5 b | 30 ± 10.7 ab | 49.4 ± 18.7 ab | 23.1 ± 2.2 ab | 56.3 |
| Percentage of naturalized species covered of understory (%) | 96.8 ± 5 a | 51.5 ± 41.9 b | 45.6 ± 22.6 ab | 88.2 ± 12.1 ab | 10.3 ± 6.1 b | 97.1 |
| Average munber of species in soil seed bank | 13.8 ± 4.4 a | 15.1 ± 5.2 a | 14.8 ± 1.7 a | 15.7 ± 1.5 a | 13.8 ± 3.2 a | 10.0 |
| Average seed reserve in soil seed bank (seeds/m2) | 771.0 ± 494.5 a | 523.5 ± 482.6 a | 618.3 ± 460.9 a | 748.8 ± 398.3 a | 928.5 ± 324.4 a | 589.0 |
| Average number of tree species in soil seed bank | 0.4 ± 0.5 b | 2.6 ± 1.4 a | 2.3 ± 0.8 a | 2.5 ± 0.5 a | 2.5 ± 1.5 ab | 1.0 |
| Average tree seed storage in soil seed bank (seed/m2) | 0.7 ± 1.0 b | 19.2 ± 18.9 a | 6.3 ± 3.9 ab | 10.2 ± 10.4 ab | 7.0 ± 4.1 ab | 404.0 |
| Percentage of naturalized species in soil seed bank (%) | 73.0 ± 5.5 a | 56.4 ± 12.2 b | 60.3 ± 8.1 ab | 65.5 ± 7.7 ab | 52.4 ± 11.6 ab | 90.0 |
| Percentage of soil seed reserves of naturalized species (%) | 97.3 ± 2.3 a | 78.0 ± 21.9 a | 95.7 ± 2.6 a | 95.6 ± 4.2 a | 87.7 ± 11.8 a | 31.4 |
| The average Sørensen similarity index between aboveground vegetation and the soil seed bank | 0.28 ± 0.11 a | 0.20 ± 0.12 a | 0.16 ± 0.05 a | 0.50 ± 0.12 a | 0.19 ± 0.08 a | 0.31 |
| Range of Sørensen similarity index between aboveground vegetation and soil seed bank | 0.15–0.45 | 0.00–0.50 | 0.09–0.23 | 0.36–0.64 | 0.09–0.26 | 0.31 |
| Species | Above Ground Vegetation 1 | Growth Type | Native/ Naturalized | Life Span 2 | Panicum maximum Type (n = 9) 3 | Acacia confusa Type (n = 20) | Ficus microcarpa Type (n = 6) | Litsea glutinosa Type (n = 6) | Cinnamomum camphora Type (n = 4) | Trema orientalis Type (n = 1) | Total | Ratio (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Spermacoce latifolia | + | Herb | Naturalized | A | 3739.9 | 4357.0 | 7437.0 | 9022.2 | 10940.7 | 1481.5 | 5756.5 | 59.75 |
| Solanum americanum | + | Herb | Naturalized | A | 2953.1 | 463.7 | 884.0 | 548.1 | 448.1 | 59.3 | 1006.4 | 10.45 |
| Praxelis clematidea | + | Herb | Naturalized | A | 2388.5 | 854.1 | 143.2 | 390.1 | 392.6 | 148.1 | 945.6 | 9.81 |
| Panicum maximum | + | Herb | Naturalized | P | 1121.0 | 739.3 | 145.7 | 118.5 | 322.2 | 148.1 | 606.4 | 6.29 |
| Kyllinga brevifolia | − | Herb | Native | P | 3.3 | 521.5 | 2.5 | 0.0 | 11.1 | 0.0 | 228.7 | 2.37 |
| Trema orientalis | + | Tree | Native | P | 6.6 | 93.3 | 64.2 | 74.1 | 66.7 | 5985.2 | 195.8 | 2.03 |
| Conyza sumatrensis | − | Herb | Naturalized | A | 261.7 | 15.6 | 49.4 | 434.6 | 0.0 | 0.0 | 121.1 | 1.26 |
| Oplismenus compositus | + | Herb | Native | P | 37.9 | 113.3 | 7.4 | 0.0 | 518.5 | 0.0 | 102.7 | 1.07 |
| Mikania micrantha | + | Vine | Naturalized | P | 14.8 | 68.9 | 22.2 | 113.6 | 118.5 | 666.7 | 75.4 | 0.78 |
| Soliva anthemifolia | − | Herb | Naturalized | A | 339.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 66.3 | 0.69 |
| Acacia confusa | + | Tree | Native | P | 0.0 | 123.7 | 17.3 | 61.7 | 14.8 | 0.0 | 65.4 | 0.68 |
| Bidens pilosa var. radiata | + | Herb | Naturalized | A | 174.5 | 42.2 | 19.8 | 12.3 | 40.7 | 118.5 | 62.8 | 0.65 |
| Gnaphalium purpureum | − | Herb | Naturalized | A | 107.0 | 29.6 | 29.6 | 39.5 | 11.1 | 0.0 | 43.8 | 0.45 |
| Oxalis corniculata | + | Herb | Native | A | 31.3 | 15.6 | 51.9 | 79.0 | 151.9 | 0.0 | 43.2 | 0.45 |
| Centella asiatica | + | Herb | Native | P | 0.0 | 1.5 | 192.6 | 4.9 | 118.5 | 0.0 | 36.7 | 0.38 |
| Toddalia asiatica | + | Vine | Native | P | 0.0 | 0.0 | 0.0 | 0.0 | 400.0 | 0.0 | 34.8 | 0.36 |
| Acronychia pedunculata | + | Tree | Native | P | 0.0 | 49.6 | 0.0 | 0.0 | 3.7 | 0.0 | 21.9 | 0.23 |
| Urena lobata | + | Shrub | Native | P | 1.6 | 40.7 | 9.9 | 2.5 | 7.4 | 0.0 | 20.3 | 0.21 |
| Lindernia crustacea | − | Herb | Native | A | 6.6 | 0.7 | 14.8 | 61.7 | 70.4 | 0.0 | 17.7 | 0.18 |
| Youngia japonica | + | Herb | Native | A | 46.1 | 10.4 | 0.0 | 0.0 | 0.0 | 0.0 | 42 | 0.14 |
| Eleusine indica | − | Herb | Native | A | 24.7 | 13.3 | 0.0 | 0.0 | 0.0 | 0.0 | 33 | 0.11 |
| Polygonum chinense | + | Herb | Native | A | 1.6 | 23.0 | 0.0 | 0.0 | 0.0 | 0.0 | 32 | 0.11 |
| Cyperus compressus | − | Herb | Native | A | 13.2 | 16.3 | 0.0 | 0.0 | 0.0 | 0.0 | 30 | 0.10 |
| Cardamine flexuosa | − | Herb | Naturalized | A | 0.0 | 18.5 | 4.9 | 4.9 | 0.0 | 0.0 | 29 | 0.10 |
| Sida alnifolia | − | Shrub | Native | P | 0.0 | 19.3 | 0.0 | 0.0 | 0.0 | 0.0 | 26 | 0.09 |
| Pluchea sagittalis | − | Herb | Naturalized | P | 13.2 | 5.9 | 0.0 | 19.8 | 7.4 | 0.0 | 26 | 0.09 |
| Elephantopus mollis | − | Herb | Naturalized | P | 1.6 | 14.1 | 0.0 | 0.0 | 0.0 | 29.6 | 22 | 0.07 |
| Miscanthus floridulus | + | Herb | Native | P | 0.0 | 2.2 | 4.9 | 2.5 | 48.1 | 0.0 | 19 | 0.06 |
| Vernonia cinerea | + | Herb | Native | A | 16.5 | 5.9 | 0.0 | 0.0 | 0.0 | 0.0 | 18 | 0.06 |
| Rhynchelytrum repens | + | Herb | Naturalized | P | 28.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 17 | 0.06 |
| Phytolacca americana | − | Herb | Naturalized | P | 23.0 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 | 16 | 0.05 |
| Flueggea suffruticosa | − | Shrub | Native | P | 0.0 | 9.6 | 0.0 | 0.0 | 0.0 | 0.0 | 13 | 0.04 |
| Cyperus esculentus | − | Herb | Naturalized | P | 4.9 | 0.0 | 7.4 | 17.3 | 0.0 | 0.0 | 13 | 0.04 |
| Ixeris chinensis | − | Herb | Native | P | 18.1 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 12 | 0.04 |
| Chloris barbata | − | Herb | Naturalized | P | 0.0 | 5.9 | 4.9 | 0.0 | 3.7 | 0.0 | 11 | 0.04 |
| Boehmeria nivea | − | Shrub | Naturalized | P | 0.0 | 5.2 | 0.0 | 0.0 | 14.8 | 0.0 | 11 | 0.04 |
| Scoparia dulcis | − | Herb | Naturalized | A | 0.0 | 0.0 | 0.0 | 9.9 | 25.9 | 0.0 | 11 | 0.04 |
| Ipomoea obscura | + | Vine | Naturalized | P | 0.0 | 3.0 | 0.0 | 0.0 | 0.0 | 88.9 | 10 | 0.03 |
| Hedyotis corymbosa | − | Herb | Native | A | 1.6 | 5.2 | 0.0 | 4.9 | 0.0 | 0.0 | 10 | 0.03 |
| Conyza canadensis | − | Herb | Naturalized | A | 13.2 | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 9 | 0.03 |
| Scleria terrestris | − | Herb | Native | P | 0.0 | 4.4 | 7.4 | 0.0 | 0.0 | 0.0 | 9 | 0.03 |
| Ficus microcarpa | + | Tree | Native | P | 0.0 | 1.5 | 2.5 | 12.3 | 3.7 | 0.0 | 9 | 0.03 |
| Ipomoea nil | − | Vine | Naturalized | A | 0.0 | 5.9 | 0.0 | 0.0 | 0.0 | 0.0 | 8 | 0.03 |
| Symplocos chinensis | + | Shrub | Native | P | 0.0 | 0.7 | 2.5 | 14.8 | 0.0 | 0.0 | 8 | 0.03 |
| Axonopus compressus | − | Herb | Naturalized | P | 0.0 | 5.9 | 0.0 | 0.0 | 0.0 | 0.0 | 8 | 0.03 |
| Zanthoxylum avicennae | + | Tree | Native | P | 0.0 | 3.7 | 0.0 | 0.0 | 7.4 | 0.0 | 7 | 0.02 |
| Mallotus japonicus | + | Tree | Native | P | 0.0 | 5.2 | 0.0 | 0.0 | 0.0 | 0.0 | 7 | 0.02 |
| Ficus subpisocarpa | − | Tree | Native | P | 0.0 | 3.0 | 4.9 | 0.0 | 3.7 | 0.0 | 7 | 0.02 |
| Ageratum houstonianum | + | Herb | Naturalized | A | 0.0 | 3.0 | 4.9 | 0.0 | 0.0 | 0.0 | 6 | 0.02 |
| Mimosa pudica | + | Shrub | Naturalized | P | 0.0 | 3.7 | 0.0 | 0.0 | 0.0 | 0.0 | 5 | 0.02 |
| Lantana camara | + | Shrub | Naturalized | P | 0.0 | 2.2 | 0.0 | 4.9 | 0.0 | 0.0 | 5 | 0.02 |
| Morus alba | + | Shrub | Native | P | 1.6 | 3.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5 | 0.02 |
| Tephrosia noctiflora | − | Herb | Naturalized | A | 6.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4 | 0.01 |
| Lepidagathis inaequalis | − | Herb | Native | P | 1.6 | 1.5 | 2.5 | 0.0 | 0.0 | 0.0 | 4 | 0.01 |
| Litsea glutinosa | + | Shrub | Naturalized | P | 0.0 | 0.0 | 0.0 | 9.9 | 0.0 | 0.0 | 4 | 0.01 |
| Mussaenda parviflora | + | Vine | Native | P | 0.0 | 2.2 | 2.5 | 0.0 | 0.0 | 0.0 | 4 | 0.01 |
| Hedyotis dichotoma | − | Herb | Native | A | 0.0 | 0.7 | 0.0 | 7.4 | 0.0 | 0.0 | 4 | 0.01 |
| Drymaria diandra | − | Herb | Naturalized | A-P | 0.0 | 3.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4 | 0.01 |
| Ixeris polycephala | − | Herb | Native | A | 4.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3 | 0.01 |
| Broussonetia papyrifera | + | Tree | Native | P | 0.0 | 1.5 | 2.5 | 0.0 | 0.0 | 0.0 | 3 | 0.01 |
| Chromolaena odorata | + | Herb | Naturalized | P | 1.6 | 0.0 | 2.5 | 2.5 | 0.0 | 0.0 | 3 | 0.01 |
| Rubus parvifolius | − | Shrub | Native | P | 0.0 | 0.0 | 4.9 | 0.0 | 0.0 | 0.0 | 2 | 0.01 |
| Liquidambar formosana | − | Tree | Native | P | 1.6 | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 2 | 0.01 |
| Cyperus cyperoides | − | Herb | Native | P | 3.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2 | 0.01 |
| Melochia corchorifolia | − | Shrub | Naturalized | P | 0.0 | 0.7 | 2.5 | 0.0 | 0.0 | 0.0 | 2 | 0.01 |
| Solanum trianthum | − | Shrub | Naturalized | P | 1.6 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 2 | 0.01 |
| Maesa perlaria var. formosana | − | Shrub | Native | P | 0.0 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 | 2 | 0.01 |
| Macaranga tanarius | + | Tree | Native | P | 1.6 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 2 | 0.01 |
| Alpinia zerumbet | + | Herb | Native | P | 0.0 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 | 2 | 0.01 |
| Dianella ensifolia | + | Herb | Native | P | 0.0 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 | 2 | 0.01 |
| Passiflora suberosa | + | Vine | Naturalized | P | 0.0 | 0.7 | 0.0 | 2.5 | 0.0 | 0.0 | 2 | 0.01 |
| Bridelia monoica | + | Tree | Native | P | 0.0 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 1 | <0.01 |
| Sarcandra glabra | − | Shrub | Native | P | 0.0 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 1 | <0.01 |
| Cinnamomum camphora | + | Tree | Native | P | 0.0 | 0.0 | 0.0 | 0.0 | 3.7 | 0.0 | 1 | <0.01 |
| Crotalaria zanzibarica | + | Shrub | Naturalized | A | 0.0 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 1 | <0.01 |
| Sapium sebiferum | + | Tree | Naturalized | P | 0.0 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 1 | <0.01 |
| Mallotus repandus | + | Vine | Native | P | 0.0 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 1 | <0.01 |
| Fimbristylis aestivalis | − | Herb | Native | A | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1 | <0.01 |
| Polygonum plebeium | − | Herb | Naturalized | A | 0.0 | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 1 | <0.01 |
| Epaltes australis | − | Herb | Native | A | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1 | <0.01 |
| Morinda parvifolia | + | Vine | Native | P | 0.0 | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 1 | <0.01 |
| Melia azedarach | + | Tree | Native | P | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 0.0 | 1 | <0.01 |
| Gnaphalium purpureum | − | Herb | Naturalized | A | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 0.0 | 1 | <0.01 |
| Lophatherum gracile | + | Herb | Native | P | 0.0 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 1 | <0.01 |
| Clerodendrum cyrtophyllum | + | Shrub | Native | P | 0.0 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 1 | <0.01 |
| Sonchus oleraceus | − | Herb | Naturalized | A | 0.0 | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 1 | <0.01 |
| Momordica charantia var. abbreviata | + | Vine | Naturalized | A | 0.0 | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 1 | <0.01 |
| Pericampylus glaucus | + | Vine | Native | P | 0.0 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 1 | <0.01 |
| Ageratum conyzoides | − | Herb | Naturalized | A | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 0.0 | 1 | <0.01 |
| Bothriospermum zeylanicum | − | Herb | Native | A | 0.0 | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 1 | <0.01 |
| Duchesnea indica | − | Herb | Naturalized | P | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1 | <0.01 |
| Total | 11,422.2 | 7754.8 | 9160.5 | 11,093.8 | 13,755.6 | 8725.9 | 29,914 | 100.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.-Y.; Liao, M.-C.; Wang, W.; Tzeng, H.-Y. Composition Characteristics of an Urban Forest Soil Seed Bank and Its Influence on Vegetation Restoration: A Case Study in Dadu Terrace, Central Taiwan. Sustainability 2022, 14, 4178. https://doi.org/10.3390/su14074178
Lin C-Y, Liao M-C, Wang W, Tzeng H-Y. Composition Characteristics of an Urban Forest Soil Seed Bank and Its Influence on Vegetation Restoration: A Case Study in Dadu Terrace, Central Taiwan. Sustainability. 2022; 14(7):4178. https://doi.org/10.3390/su14074178
Chicago/Turabian StyleLin, Chia-Yen, Min-Chun Liao, Wei Wang, and Hsy-Yu Tzeng. 2022. "Composition Characteristics of an Urban Forest Soil Seed Bank and Its Influence on Vegetation Restoration: A Case Study in Dadu Terrace, Central Taiwan" Sustainability 14, no. 7: 4178. https://doi.org/10.3390/su14074178
APA StyleLin, C. -Y., Liao, M. -C., Wang, W., & Tzeng, H. -Y. (2022). Composition Characteristics of an Urban Forest Soil Seed Bank and Its Influence on Vegetation Restoration: A Case Study in Dadu Terrace, Central Taiwan. Sustainability, 14(7), 4178. https://doi.org/10.3390/su14074178