
Thauera sp. in Hydrogen-Based Denitrification: Effects of Plentiful Bicarbonate Supplementation on Powerful Nitrite Reducer


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation
2.2. Identification of Bacterial Strain
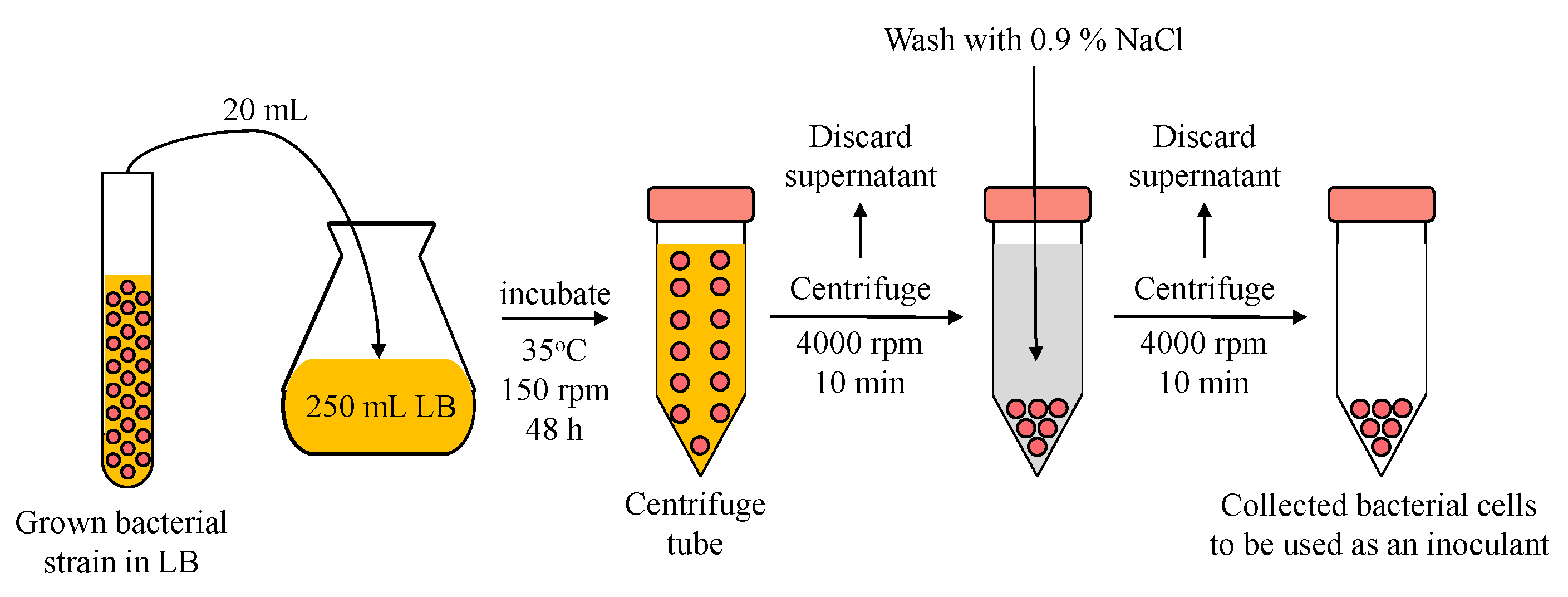
2.3. Inoculants Preparation
2.4. Effects of HCO3− on Bacterial Behavior in Performing Nitrogen Removal
2.5. Analytical Methods for Water Samples and Necessary Calculations
3. Results and Discussion
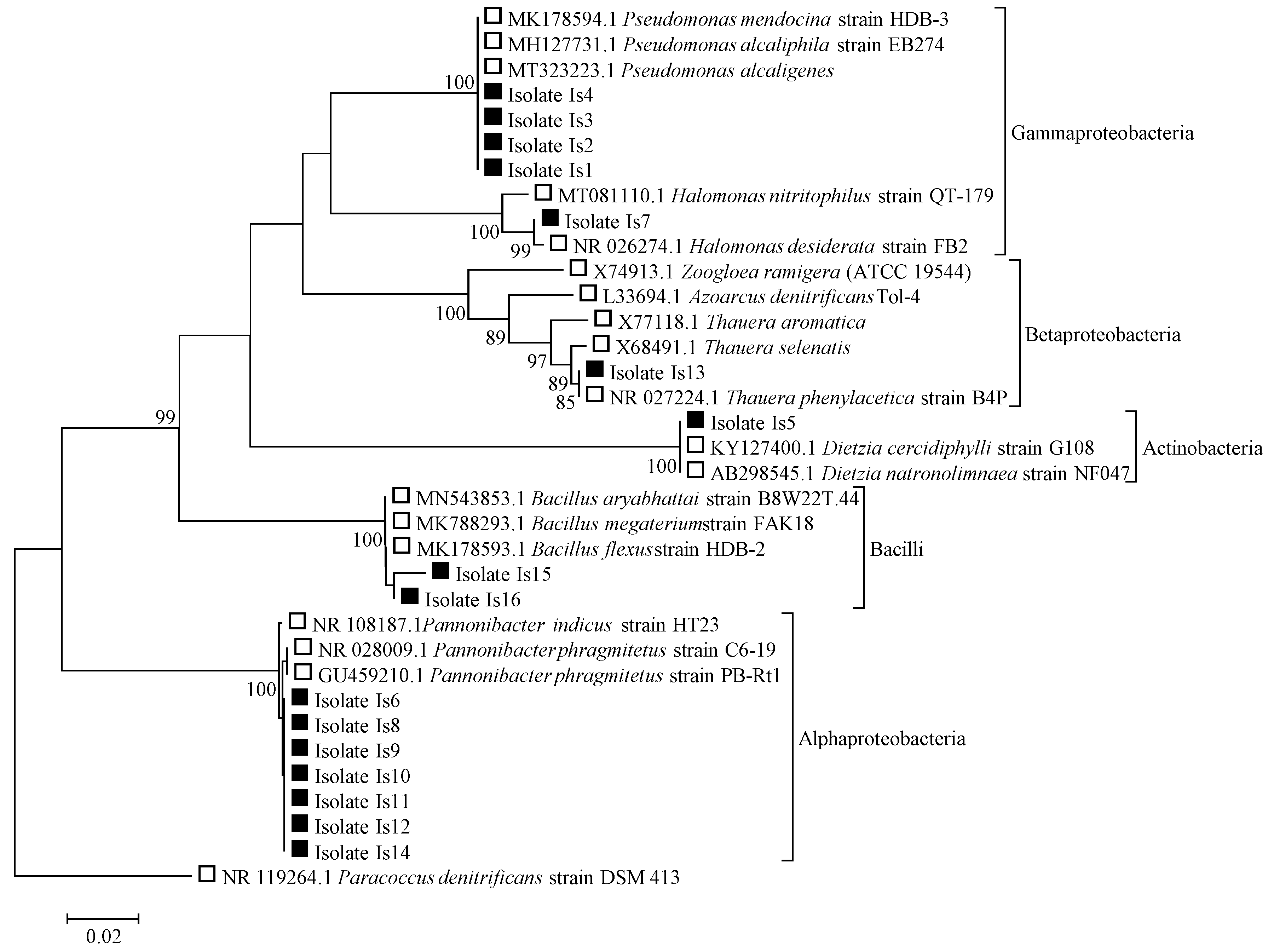
3.1. Isolation of Single Bacterial Strains from the Mixed-Culture Sludge
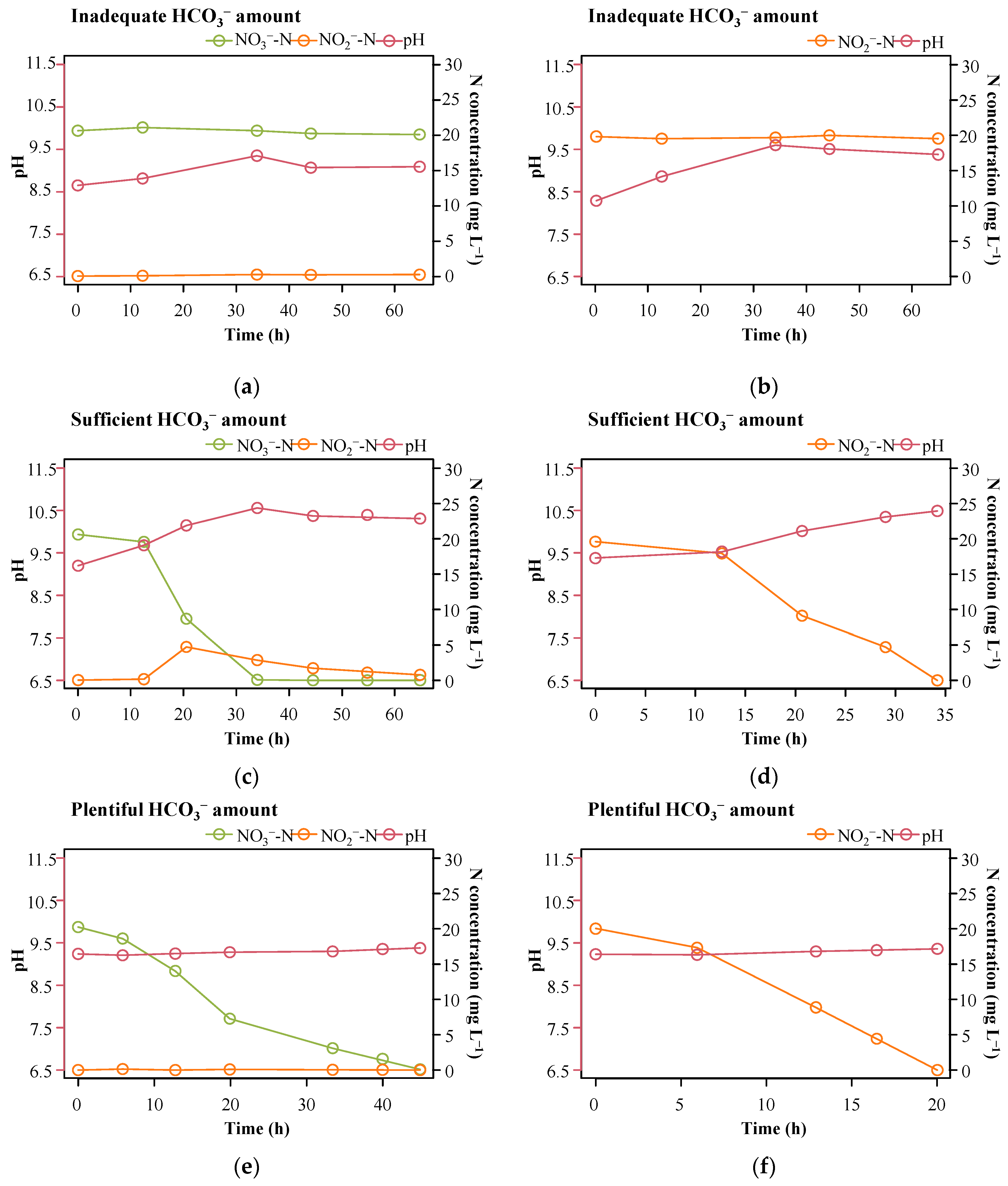
3.2. Behavior of Single Isolated Strains in HD under Plentiful HCO3− Supplementary Conditions
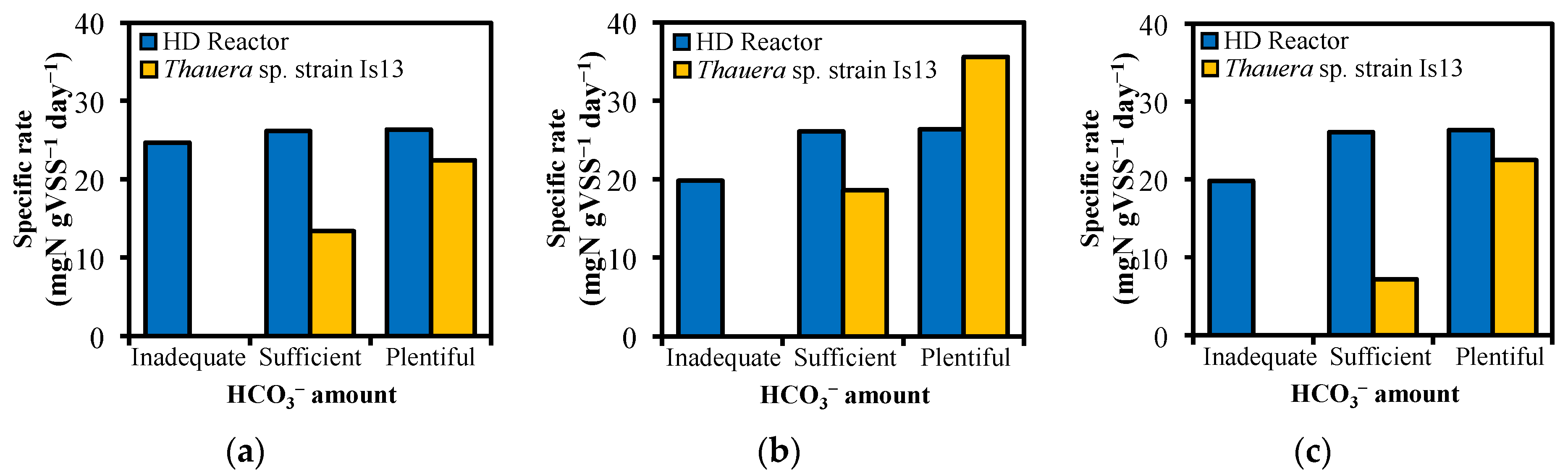
3.3. Effects of HCO3− on Thauera sp. in Performing Nitrogen Removal
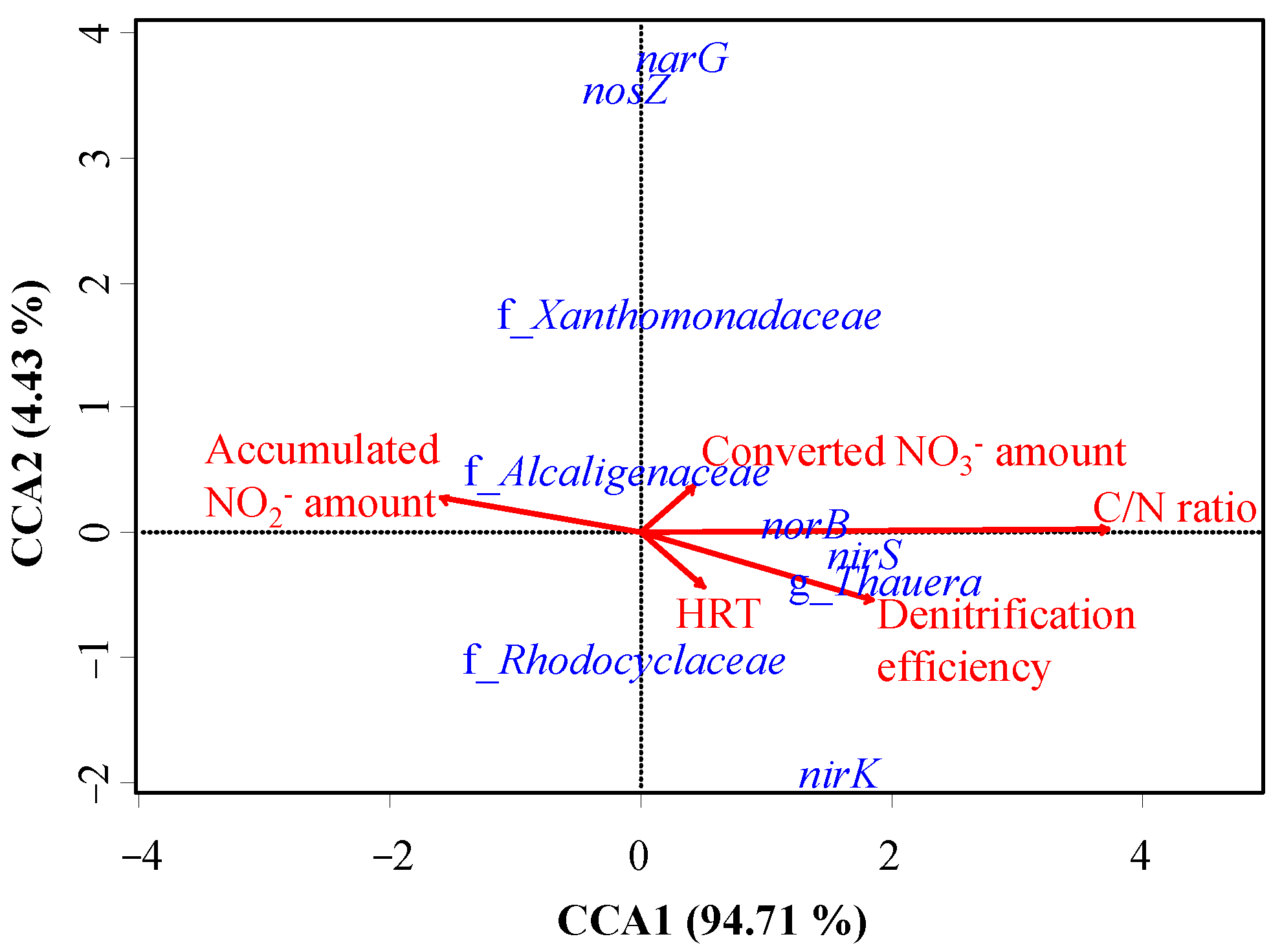
3.4. Contribution of Thauera sp. on Nitrogen Removal in Actual HD Reactor
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, S.S.; Cheng, H.Y.; Zhang, H.; Su, S.G.; Sun, Y.L.; Wang, H.C.; Han, J.L.; Wang, A.J.; Guadie, A. Sulfur autotrophic denitrification filter and heterotrophic denitrification filter: Comparison on denitrification performance, hydrodynamic characteristics and operating cost. Environ. Res. 2021, 197, 111029. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.; Li, N.; Luo, H.; Luo, X.; Shen, T.; Yang, Y.; Jiang, J. Autotrophic Fe-Driven Biological Nitrogen Removal Technologies for Sustainable Wastewater Treatment. Front. Microbiol. 2022, 13, 895409. [Google Scholar] [CrossRef] [PubMed]
- Rujakom, S.; Shinoda, K.; Kamei, T.; Kazama, F. Investigation of hydrogen-based denitrification performance on nitrite accumulation under various bicarbonate doses. Environ. Asia 2019, 12, 54–63. [Google Scholar] [CrossRef]
- Lau, G.N.; Sharma, K.R.; Chen, G.H.; Van Loosdrecht MC, M. Integration of sulphate reduction, autotrophic denitrification and nitrification to achieve low-cost excess sludge minimisation for Hong Kong sewage. Water Sci. Technol. 2006, 53, 227–235. [Google Scholar] [CrossRef]
- Hu, Y.; Wu, G.; Li, R.; Xiao, L.; Zhan, X. Iron sulphides mediated autotrophic denitrification: An emerging bioprocess for nitrate pollution mitigation and sustainable wastewater treatment. Water Res. 2020, 179, 115914. [Google Scholar] [CrossRef] [PubMed]
- Xing, W.; Li, J.; Li, P.; Wang, C.; Cao, Y.; Li, D.; Yang, Y.; Zhou, J.; Zuo, J. Effects of residual organics in municipal wastewater on hydrogenotrophic denitrifying microbial communities. J. Environ. Sci. 2018, 65, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Sunger, N.; Bose, P. Autotrophic denitrification using hydrogen generated from metallic iron corrosion. Bioresour. Technol. 2009, 100, 4077–4082. [Google Scholar] [CrossRef] [PubMed]
- Di Capua, F.; Pirozzi, F.; Lens, P.N.L.; Esposito, G. Electron donors for autotrophic denitrification. Chem. Eng. J. 2019, 362, 922–937. [Google Scholar] [CrossRef]
- Rujakom, S.; Shinoda, K.; Singhopon, T.; Nakano, M.; Kamei, T.; Kazama, F. Effect of bicarbonate on the performance of hydrogen-based denitrification at different hydraulic retention times. J. Water Treat. Biol. 2020, 56, 33–45. [Google Scholar] [CrossRef]
- Chen, D.; Yang, K.; Wang, H. Effects of important factors on hydrogen-based autotrophic denitrification in a bioreactor. Desalination Water Treat. 2016, 57, 3482–3488. [Google Scholar] [CrossRef]
- Ghafari, S.; Hasan, M.; Aroua, M.K. Effect of carbon dioxide and bicarbonate as inorganic carbon sources on growth and adaptation of autohydrogenotrophic denitrifying bacteria. J. Hazard. Mater. 2009, 162, 1507–1513. [Google Scholar] [CrossRef] [PubMed]
- Ghafari, S.; Hasan, M.; Aroua, M.K. Improvement of autohydrogenotrophic nitrite reduction rate through optimization of pH and sodium bicarbonate dose in batch experiments. J. Biosci. Bioeng. 2009, 107, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Rezania, B.; Oleszkiewicz, J.A.; Cicek, N.; Mo, H. Hydrogen-dependent denitrification in an alternating anoxic-aerobic SBR membrane bioreactor. Water Sci. Technol. 2005, 51, 403–409. Available online: https://iwaponline.com/wst/article-pdf/51/6-7/403/435278/403.pdf (accessed on 12 October 2022). [CrossRef] [PubMed]
- Visvanathan, C.; Hung, N.Q.; Jegatheesan, V. Hydrogenotrophic denitrification of synthetic aquaculture wastewater using membrane bioreactor. Process Biochem. 2008, 43, 673–682. [Google Scholar] [CrossRef]
- Lee, J.W.; Lee, K.H.; Park, K.Y.; Maeng, S.K. Hydrogenotrophic denitrification in a packed bed reactor: Effects of hydrogen-to-water flow rate ratio. Bioresour. Technol. 2010, 101, 3940–3946. [Google Scholar] [CrossRef]
- Li, P.; Xing, W.; Zuo, J.; Tang, L.; Wang, Y.; Lin, J. Hydrogenotrophic denitrification for tertiary nitrogen removal from municipal wastewater using membrane diffusion packed-bed bioreactor. Bioresour. Technol. 2013, 144, 452–459. [Google Scholar] [CrossRef]
- Inagaki, Y.; Yamada, D.; Komori, M.; Sakakibara, Y. Field application of hydrogenotrophic denitrification with two-stage injection of electrolytic hydrogen. J. Water Process Eng. 2020, 38, 101685. [Google Scholar] [CrossRef]
- Mao, Y.; Xia, Y.; Zhang, T. Characterization of Thauera-dominated hydrogen-oxidizing autotrophic denitrifying microbial communities by using high-throughput sequencing. Bioresour. Technol. 2013, 128, 703–710. [Google Scholar] [CrossRef]
- Macy, J.M.; Rech, S.; Auling, G.; Dorsch, M.; Stackebrandt, E.; Sly, L.I. Thauera selenatis gen. nov., sp., nov., a Member of the Beta Subclass of Proteobacteria with a Novel Type of Anaerobic Respiration. Int. J. Syst. Bacteriol. 1993, 43, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Song, B.; Young, L.Y.; Palleroni, N.J. Identification of denitrifier strain T I as Thauera arornatica and proposal for emendation of the genus Thauera definition. Int. J. Syst. Bacteriol. 1998, 48, 889–894. [Google Scholar] [CrossRef]
- Jeter, R.M.; Ingraham, J.L. The Denitrifying Prokaryotes. In The Prokaryotes; Starr, M.P., Stolp, H., Trüper, H.G., Balows, A., Schlegel, H.G., Eds.; Springer: Berlin/Heidelberg, Germany, 1981; pp. 913–925. [Google Scholar] [CrossRef]
- Liang, Z.; Sun, J.; Zhan, C.; Wu, S.; Zhang, L.; Jiang, F. Effects of sulfide on mixotrophic denitrification by: Thauera-dominated denitrifying sludge. Environ. Sci. Water Res. Technol. 2020, 6, 1186–1195. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S Ribosomal DNA Amplification for Phylogenetic Study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, R.; Imai, I.; Fukami, K.; Minami, A.; Hiroishi, S. Phylogenetic analysis of algicidal bacteria (family Flavobacteriaceae) and selective detection by PCR using a specific primer set. Fish. Sci. 1999, 65, 432–435. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Annapurna, D.; Rajkumar, M.; Prasad, M.N.V. Potential of castor bean (Ricinus communis L.) for phytoremediation of metalliferous waste assisted by plant growth-promoting bacteria: Possible cogeneration of economic products. In Bioremediation and Bioeconomy; Prasad, M.N.V., Ed.; Elsevier Inc: Amsterdam, The Netherlands, 2016; pp. 149–175. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water and Wastewater, 22nd ed.; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]
- Arat, S.; Bullerjahn, G.S.; Laubenbacher, R. A network biology approach to denitrification in Pseudomonas aeruginosa. PLoS ONE 2015, 10, e0118235. [Google Scholar] [CrossRef] [Green Version]
- Jun, K.I.; Abraham, A.; Choi, O.; Sang, B.I. Aerobic denitrification by a novel Pseudomonas sp. JN5 in different bioreactor systems. Water-Energy Nexus 2019, 2, 37–45. [Google Scholar] [CrossRef]
- von der Weid, I.; Marques, J.M.; Cunha, C.D.; Lippi, R.K.; dos Santos, S.C.C.; Rosado, A.S.; Lins, U.; Seldin, L. Identification and biodegradation potential of a novel strain of Dietzia cinnamea isolated from a petroleum-contaminated tropical soil. Syst. Appl. Microbiol. 2007, 30, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Bødtker, G.; Hvidsten, I.V.; Barth, T.; Torsvik, T. Hydrocarbon degradation by Dietzia sp. A14101 isolated from an oil reservoir model column. Antonie Van Leeuwenhoek 2009, 96, 459–469. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, H.; Shutes, B.; Fu, B.; Yan, B.; Yu, X.; Wen, H.; Chen, X. Identification and denitrification characteristics of a salt-tolerant denitrifying bacterium Pannonibacter phragmitetus F1. AMB Express 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, Y.; Bohu, T.; Wu, S.; Bai, Z.; Zhuang, X. Nitrogen Removal Characteristics and Constraints of an Alphaproteobacteria with Potential for High Nitrogen Content Heterotrophic Nitrification-Aerobic Denitrification. Microorganisms 2022, 10, 235. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhou, X.; Li, Y.; Li, K.; Wang, C.; Liu, J.; Yan, D.; Liu, Y.; Yang, D.; Xing, J. Heterotrophic nitrification and aerobic denitrification by a novel Halomonas campisalis. Biotechnol. Lett. 2013, 35, 2045–2049. [Google Scholar] [CrossRef]
- Wang, L.; Shao, Z. Aerobic Denitrification and Heterotrophic Sulfur Oxidation in the Genus Halomonas Revealed by Six Novel Species Characterizations and Genome-Based Analysis. Front. Microbiol. 2021, 12, 652766. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Mao, Y.; Bergaust, L.; Bakken, L.R.; Frostegård, Å. Strains in the genus Thauera exhibit remarkably different denitrification regulatory phenotypes. Environ. Microbiol. 2013, 15, 2816–2828. [Google Scholar] [CrossRef]
- Sun, Y.; Shen, D.; Zhou, X.; Shi, N.; Tian, Y. Microbial diversity and community structure of denitrifying biological filters operated with different carbon sources. Springerplus 2016, 5, 652766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Chen, Y.; Xiao, L. Metagenomic insights into mixotrophic denitrification facilitated nitrogen removal in a full-scale A2/O wastewater treatment plant. PLoS ONE 2021, 16, e0250283. [Google Scholar] [CrossRef]
- Kim, J.K.; Park, K.J.; Cho, K.S.; Wan, N.S.; Park, T.J.; Bajpai, R. Aerobic nitrification-denitrification by heterotrophic Bacillus strains. Bioresour. Technol. 2005, 96, 1897–1906. [Google Scholar] [CrossRef]
- Barman, P.; Bandyopadhyay, P.; Kati, A.; Paul, T.; Mandal, A.K.; Mondal, K.C.; Mohapatra, P.K.D. Characterization and Strain Improvement of Aerobic Denitrifying EPS Producing Bacterium Bacillus cereus PB88 for Shrimp Water Quality Management. Waste Biomass Valorization 2018, 9, 1319–1330. [Google Scholar] [CrossRef]
- Yang, T.; Yang, Q.; Shi, Y.; Xin, Y.; Zhang, L.; Gu, Z.; Shi, G. Insight into the denitrification mechanism of Bacillus subtilis JD-014 and its application potential in bioremediation of nitrogen wastewater. Process Biochem. 2021, 103, 78–86. [Google Scholar] [CrossRef]
- Ghafari, S.; Hasan, M.; Aroua, M.K. A kinetic study of autohydrogenotrophic denitrification at the optimum pH and sodium bicarbonate dose. Bioresour. Technol. 2010, 101, 2236–2242. [Google Scholar] [CrossRef] [PubMed]
- Sander, E.M.; Virdis, B.; Freguia, S. Bioelectrochemical denitrification for the treatment of saltwater recirculating aquaculture streams. ACS Omega 2018, 3, 4252–4261. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Accession No. | % Similarity | Closest Match | Phylogenetic Group (Family) |
|---|---|---|---|---|
| Is1 | OP676119 | 100.00 | Pseudomonas mendocina strain HDB-3 | Pseudomonadaceae |
| Is2 | OP676120 | 100.00 | Pseudomonas mendocina strain HDB-3 | Pseudomonadaceae |
| Is3 | OP676121 | 100.00 | Pseudomonas mendocina strain HDB-3 | Pseudomonadaceae |
| Is4 | OP676122 | 100.00 | Pseudomonas mendocina strain HDB-3 | Pseudomonadaceae |
| Is5 | OP676123 | 100.00 | Dietzia natronolimnaea strain NF047 | Dietziaceae |
| Is6 | OP676124 | 99.89 | Pannonibacter phragmitetus strain PB-Rt1 | Rhodobacteraceae |
| Is7 | OP676125 | 99.89 | Halomonas desiderata strain FB2 16S | Halomonadaceae |
| Is8 | OP676126 | 99.88 | Pannonibacter phragmitetus strain PB-Rt1 | Rhodobacteraceae |
| Is9 | OP676127 | 99.88 | Pannonibacter phragmitetus strain PB-Rt1 | Rhodobacteraceae |
| Is10 | OP676128 | 99.88 | Pannonibacter phragmitetus strain PB-Rt1 | Rhodobacteraceae |
| Is11 | OP676129 | 99.88 | Pannonibacter phragmitetus strain PB-Rt1 | Rhodobacteraceae |
| Is12 | OP676130 | 99.88 | Pannonibacter phragmitetus strain PB-Rt1 | Rhodobacteraceae |
| Is13 | OP676131 | 99.89 | Thauera phenylacetica strain B4P | Zoogloeaceae |
| Is14 | OP676132 | 99.88 | Pannonibacter phragmitetus strain PB-Rt1 | Rhodobacteraceae |
| Is15 | OP676133 | 99.34 | Bacillus flexus strain HDB-2 | Bacillaceae |
| Is16 | OP676134 | 99.83 | Bacillus aryabhattai strain B8W22T.44 | Bacillaceae |
| Bacterial Inocula | HCO3− Amount | Specific Rate (mgN gVSS−1 day−1) | ||
|---|---|---|---|---|
| NO3− Removal | NO2− Removal | Denitrification | ||
| Pseudomonas sp. strain Is1 | Plentiful | 4.15 ± 0.84 | 3.94 ± 0.80 | 4.14 ± 0.84 |
| Pannonibacter sp. strain Is6 | 15.94 ± 4.19 | n.d. * | n.d. * | |
| Bacillus sp. strain Is15 | n.d. * | n.d. * | n.d. | |
| Thauera sp. strain Is13 | Plentiful | 22.59 ± 2.66 | 36.02 ± 5.66 | 22.61 ± 2.66 |
| Sufficient | 14.32 ± 5.16 | 18.72 ± 2.14 | 7.66 ± 2.76 | |
| Inadequate | n.d. * | n.d. * | n.d. * | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rujakom, S.; Kamei, T.; Kazama, F. Thauera sp. in Hydrogen-Based Denitrification: Effects of Plentiful Bicarbonate Supplementation on Powerful Nitrite Reducer. Sustainability 2023, 15, 277. https://doi.org/10.3390/su15010277
Rujakom S, Kamei T, Kazama F. Thauera sp. in Hydrogen-Based Denitrification: Effects of Plentiful Bicarbonate Supplementation on Powerful Nitrite Reducer. Sustainability. 2023; 15(1):277. https://doi.org/10.3390/su15010277
Chicago/Turabian StyleRujakom, Suphatchai, Tatsuru Kamei, and Futaba Kazama. 2023. "Thauera sp. in Hydrogen-Based Denitrification: Effects of Plentiful Bicarbonate Supplementation on Powerful Nitrite Reducer" Sustainability 15, no. 1: 277. https://doi.org/10.3390/su15010277
APA StyleRujakom, S., Kamei, T., & Kazama, F. (2023). Thauera sp. in Hydrogen-Based Denitrification: Effects of Plentiful Bicarbonate Supplementation on Powerful Nitrite Reducer. Sustainability, 15(1), 277. https://doi.org/10.3390/su15010277