1. Introduction
Beans are among one the most important fabaceous crops grown worldwide. Brazil is the third largest producer of the genus
Phaseolus in the world, behind only Myanmar and India, respectively, and the largest producer and consumer of common bean (
P. vulgaris L.). The crop is of great importance in the diet of more than 500 million people and represents 50% of all legumes present in the world’s various diets [
1].
Due to its short cycle and superficial and small root system, common bean is a crop with demanding nutritional requirements, requiring highly available nutrients at the right time and place. Among the nutrients required by the bean plant, nitrogen (N) can be considered the most important since it is a macronutrient present in the constitution of the plant cell, composing nucleic acids, amino acids, and proteins, in addition to taking part in several processes responsible for cell division and expansion [
2,
3].
The nitrogen needed by the common bean plant can be made available via the soil (through the mineralization of organic matter), industrialized nitrogen fertilizers, non-biological fixation, and biological nitrogen fixation (BNF). Soil N is a limited source that is easily depleted after a few crops [
3]. In addition to their high cost and low efficiency, using nitrogen fertilizers necessitates an additional ecological cost due to the losses experienced through leaching in the form of nitrate and surface runoff and denitrification and volatilization processes; therefore, nitrogen fertilizers are highly pollutive [
4].
Fabaceae such as common beans can fix N from the atmosphere into nitrogen assimilable by plants when in symbiosis with bacteria of the Rhizobium genus, which are present in some inoculants. When in contact with the plant, these induce the formation of root nodules, and within these, the BNF process occurs, making this macronutrient, which is carried by the xylem in the transpiration stream, available. BNF is performed with diazotrophic bacteria that live in symbiosis with the roots of host plants, thus having an effect on the formation of nodules on the roots, which can be influenced by abiotic and biotic factors such soil pH, temperature, water deficit, cultivars and efficiency, and the low competitive capacity of rhizobia introduced via seed inoculation with the population of native soil bacteria [
5].
Although BNF supplies most of the N needed by common bean plants, it is characterized by the rapid senescence of nodules around forty-five days after the emergency, therefore reducing the need for BNF soon after flowering [
6,
7]. Given this fact, other practices not yet widely discussed in research on BNF in common beans include reinoculation, late reinoculation, or supplementary inoculation. These practices consist of carrying out inoculation at the time of sowing directly on the seed (inoculation via seed), together with the application of an inoculant after the implantation of the crop (inoculation via topdressing) by spraying the product directly onto the soil to promote the formation of new nodules and prolong the period and supply of N throughout the crop cycle. Souza [
8] observed that the supplementary inoculations in topdressing in the soybean crop at the V1, V3, V6, and R1 stages provided a greater number of nodules and a higher grain yield.
Other studies have shown that the application of an inoculant in the soybean crop at stages close to R6 promoted the addition of nodules in the root system, increasing fixed N and grain yield [
9,
10]. In common bean, the authors of [
11], working with reinoculation in winter bean, found significant differences for some nodular parameters, as the treatments involving reinoculation with
R. tropici yielded higher values when compared to the control treatment. The limited number of studies on the efficiency of reinoculation and the lack of knowledge of field conditions underscore the need for investigations on this subject. The use of these complementary techniques (seed inoculation and cover reinoculation) appears to be more sustainable, and these complementary techniques could replace the nitrogenous mineral fertilizers commonly used in bean fertilization, thereby reducing costs and alleviating environmental issues.
Therefore, this study aimed to investigate the responses of bean plants subjected to rhizobial cell reinoculation under cover applied at different plant growth stages under in-greenhouse and in-field conditions.
2. Materials and Methods
2.1. General Information
Two experiments were conducted—one in a greenhouse and the other in a field—in between March and April 2020 and in the spring–summer harvest of 2020/2021, respectively, both in the experimental area of the Goiana Agency for Technical Assistance, Rural Extension and Agricultural Research—EMATER, located in the municipality of Anápolis-GO, Brazil, the geographic location of which is as follows: latitude 16°20’12.13″ South and longitude 48°53’15.96″ West and average an altitude of 1058 m. The region’s climate, according to the Köppen classification, is humid tropical Aw [
12]. The prevailing climate data during the field experiment are shown below (
Figure 1).
The soil in the area is classified as dystrophic Red Latossol (Oxisol) and was sampled in the 0–20 cm layer for both experiments. The results of the chemical analysis were as follows: pH (CaCl2): 4.8; Ca: 1.4 cmolc dm−3; Mg: 0.6 cmolc dm−3; Al: 0.2 cmolc dm−3; H + Al: 2.4 cmolc dm−3; S: 1.6 mg dm−3; P (Mehlich): 4.2 mg dm−3; K: 82.6 mg dm−3; B: 0.2 mg dm−3; Cu: 2.8 mg dm−3; Fe: 33.8 mg dm−3; Mn: 17.6 mg dm−3; Zn: 6.3 mg dm−3; base saturation (V%): 48; organic matter: 20.5 g dm−3; sand: 415 g kg−1; silt: 105 g kg−1; clay: 480 g kg−1.
2.2. Experimental Design and Treatments
Both experiments were conducted in a randomized block design with ten treatments and four replications. The treatments were as follows: T1 = uninoculated control; T2 = inoculation via seed(VS); T3 = VS + reinoculation at the V4 stage; T4 = VS + reinoculation at the R5 stage; T5 = VS + reinoculation at the R6 stage; T6 = VS + reinoculation at the V4 and R5 stages; T7 = VS + reinoculation at the V4 and R6 stages; T8 = VS + reinoculation at the R5 and R6 stages; T9 = VS + reinoculation at the V4, R5, and R6 stages; and T10 = mineral nitrogen fertilization.
2.3. Treatment Implementation and Experimental Procedure
The soil used in the greenhouse was taken from the same area where the field experiment was conducted. According to the results of the soil analysis described above, liming was carried out using filler limestone (PRNT 100%) at the recommendation of 1.5 ton ha−1 of limestone, aiming to reach saturation by bases of 70%. After being collected in the 0–20 cm layer, the soil was air-dried, loosened, and sieved through 5 mm meshes. The base fertilizer in the treatments that received the application of rhizobial cells was carried out later using 400 mg of P, simple superphosphate as a source (18% of P2O5), and 100 mg of K in the form of potassium chloride (60% of K2O) without the addition of N. The treatment with mineral fertilizer involved the application of 16 mg of N, 400 mg of P2O5, and 100 mg of K2O in the base, with super simple urea and potassium chloride as the sources, in addition to 50 mg of N in stage V3, as coverage, with urea as the source.
Subsequently, the soil was homogenized and placed in five-kilogram pots. The seeds (except for T1 and T10) were inoculated, and five seeds were immediately sown per pot. At stage V1, the seedlings were thinned, leaving two plants per pot. The certified seeds belonging to category C1 from the BRS FC402 cultivar were used at a density of 12 plants per linear meter and characterized by their high yield and grain quality, normal cycle, semi-erect architecture, indeterminate growth habits (type II), resistance to common mosaic, rust, and moderate resistance to anthracnose (in the case of a common bean from the “carioca” bean group) [
13].
A sprayer with a capacity of 500 mL was used for reinoculation according to the product, dose, and volume of syrup specified previously. Spraying was always carried out in the late afternoon, aiming for better inoculation efficiency, directing the spray towards the plants’ root system.
Irrigations were carried out according to the crop’s water needs; they were monitored by weighing the pots to maintain controlled humidity, seeking 60% of the total pore volume, which was maintained during the experimental period via water replacement. To control the weeds and diseases involved in manual pulling and spraying with Cercobin at a dose of 700 g ha−1, we aimed to control anthracnose (Colletotrichum lindemuthianum) and angular leaf spot (Phaeoisariopsis griseola) after flowering.
The field experiment was implemented in an area that has been predominantly made up of the forage grass Brachiara decumbens for more than 10 years. Thus, it can be inferred that the populations of native bacteria, as well as the species R. tropici used specifically in the inoculation of the common bean plants, were low.
The field experiment consisted of plots of six lines each 5 m long, with a spacing between lines of 0.5 m. In each experimental plot, the four central lines were taken as the useful area. Of these, two plant lines (3 and 4) were intended for productivity analyses, while the other lines (2 and 5) were used for destructive analyses.
After the corrective reaction period, as mentioned above, the soil was prepared with one cycle of plowing and two cycles of harrowing. Then, basic fertilization was carried out in treatments 1 to 9 using 400 kg ha−1 of fertilizer formulated 00-20-20. Treatment 10, which involved mineral fertilization, received the fertilizer formulated 05-25-20 and 50 kg ha−1 of N in stage V3, wherein urea was used as a source. The seeds (except for treatments 1 and 10) were inoculated and sown immediately in the soil.
For inoculation via seeds, a dose of 150 mL was used for 50 kg of seed, and for reinoculation, a dose of 150 mL per hectare was used according to the recommendations of the manufacturer. The treatments that received the reinoculation in more than one bean growth stage received the total dose fractionated in the respective stages in both experiments. The inoculant used was liquid-specific for bean based on Rhizobium tropici (SEMIA 4077 and SEMIA 4088), with a guarantee of 2 × 109 CFU mL−1.
Weed control was carried out through the application of the herbicide Fusiflex® (30 L ha−1) at stage V2. The insecticide Engeo®S (30 mL 100 L−1) was used to control yellow–green cowbird (Diabrotica speciosa) and whitefly (Bemisia tabaci). For the preventive control of fungal diseases caused by anthracnose, angular leaf spot, and powdery mildew (Erysiphe polygoni), the fungicide Approve® (100 g p.c 20 L−1 of water) was applied. Irrigation was not carried out due to good water conditions during the bean cycle in the field. The other cultural treatments used were those normally applied to the crops.
2.4. Analyses Performed
At the R7 stage, in a greenhouse, the nodular and morphological characteristics were evaluated utilizing destructive analyses, with five plants being sampled with the aid of a straight shovel, collecting a block of soil with the plant at a depth of 25 cm with the intention of recovering most of the root system of the plants. The characteristics we evaluated were as follows: total nodules (TN), dry mass of nodules (NDM), taproot length (TRL), root dry mass (RDM), plant height (PH), shoot dry mass (SDM), leaf nitrogen content (NC), and stem diameter (SD).
The agronomic analyses were carried out at the R9 stage (maturation), when the plants showed leaf drops and straw yellow pods. The components were evaluated based on data from 10 plants collected in the central area of each experimental unit. The yield was obtained by collecting the rest of the plants from the central area of the central rows when the grains had 13% of moisture (dry base), where the evaluations of the number of pods per plant (NPP), number of grains per pod (NGP), 100-grain weight (W100), grain yield (YIELD), and final stand (FS) were carried out.
2.5. Statistical Analysis
The data obtained were subjected to an analysis of variance (
p ≤ 0.05) and, when significant, Tukey’s test. Statistical analyses were carried out using Sisvar 5.6 Software [
14].
3. Results
Based on the results of our analysis of variance (
Table 1) for the nodular and morphological characteristics analyses performed at the R7 stage, it was verified that the total nodules (TN), dry mass of nodules (NDM), taproot length (TRL), root dry mass (RDM), and shoot dry mass (SDM) were influenced by the treatments. On the other hand, characteristics such as plant height (PH), leaf nitrogen content (NC), and stem diameter (SD) were not influenced by the treatments studied.
In the case of TN, the inoculation via seed (T2) and the inoculation via seed with reinoculation at V4 stage (T3) treatments presented the highest averages, with 26 nodules plant
−1 in both, showing no differences; however, the other inoculation treatments (
Figure 2) generated an increase of 21.7 nodules plant
−1 in the formation of new nodules when compared to the non-inoculation treatment (T1) (
Figure 2).
Regarding the evaluation of the dry mass of nodules (NDM), it should be noted that the highest averages were found with the inoculation via seed treatment (T2), followed by inoculation via seed with reinoculation at the V4 stage (T3) and inoculation via seed with reinoculation at the V4 and R5 stages (T6) (
Figure 3). The lowest averages were obtained with use the of the inoculation via seed and reinoculation at the R5 stage treatment (T4), inoculation via seed and reinoculation at the V4, R5, and R6 stages treatment (T9), and with the mineral nitrogen fertilization treatment (T10), partially differentiating the results of several produced nodules, especially those derived from the use of the control treatment (T1).
The highest averages of taproot length (TRL) were verified in the treatments in which rhizobium cells were used via seed and in topdressing, equivalent to the treatment in which N was provided by a mineral fertilizer (
Figure 4). On the other hand, the lowest average referring to the TRL was found in the non-inoculation treatment (control), with a value of 16.0 cm. The other treatments generated an increase of up to 13.27 cm.
It was possible to verify that inoculation via seed with reinoculation at the V4 stage (T3) and R5 stages (T4) resulted in the highest mean values for the root dry mass (RDM), with values of 7.1 and 6.3 g plant
−1, respectively (
Figure 5). Except for the control treatment, the other treatments showed no statistical differences. The use of inoculation via seed + reinoculation at the V4 stage (T3) generated an increase of 152% in RDM compared to the control and 17% in the treatment that involved mineral nitrogen fertilization (T10).
Reinoculated treatment at the V4 stage (T3) presented the highest shoot dry mass average of 21.9 g plant
−1 (
Figure 6). The reinoculated treatment at the R5 stage (T4) yielded 15.4 g plant
−1, and the inoculation just via seed (T2) yielded an average of 11.4 g plant
−1 (
Figure 6).
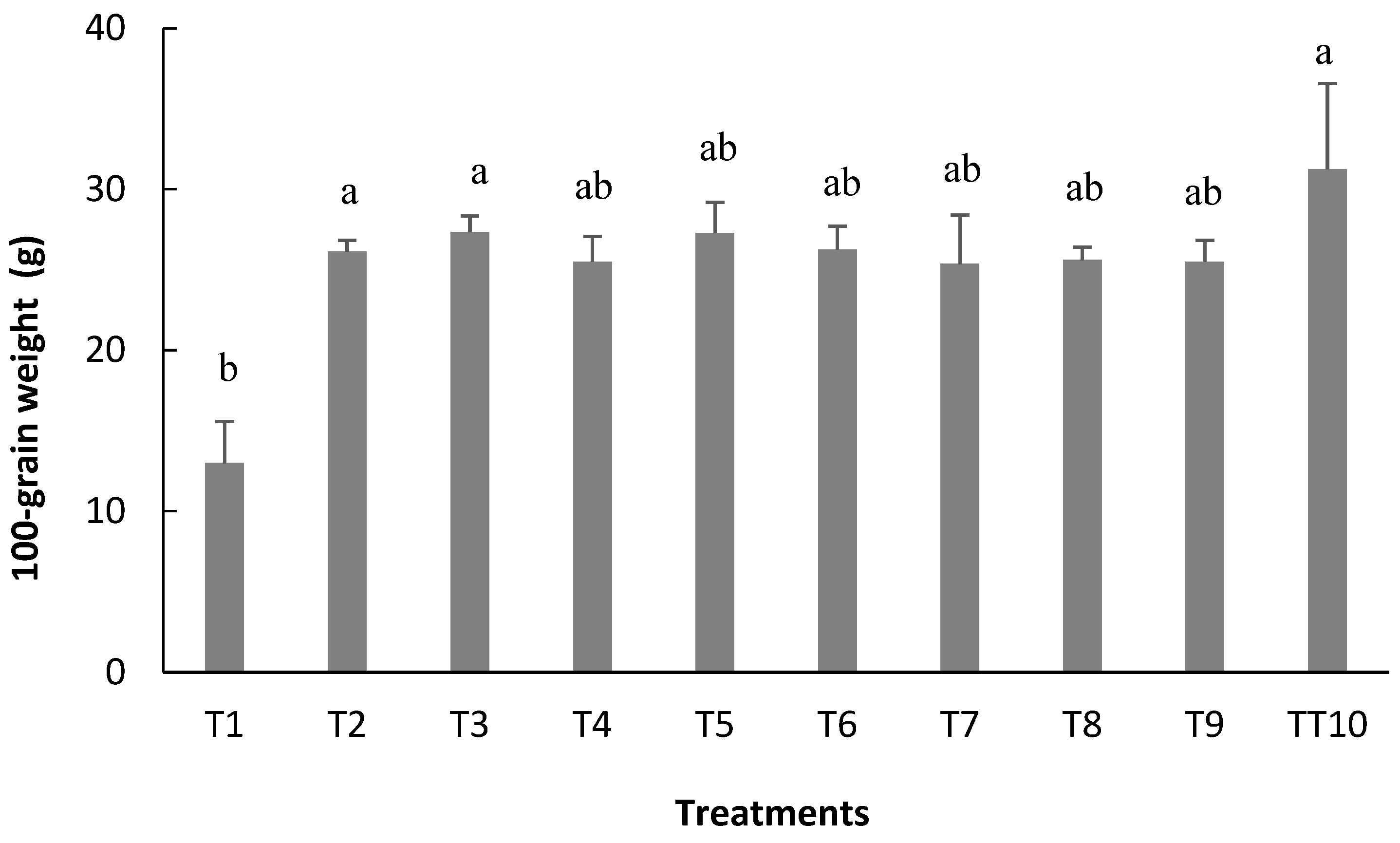
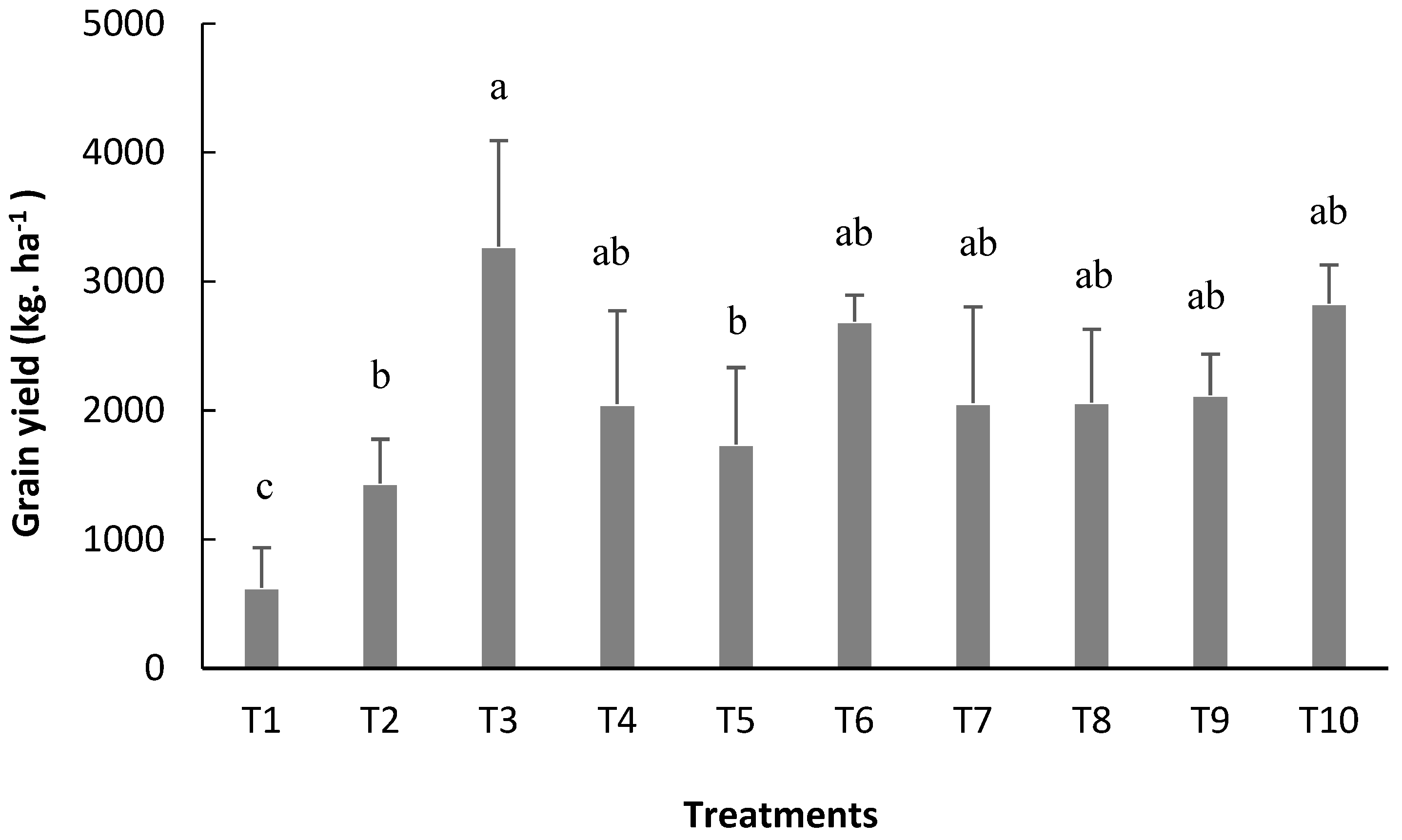
The number of pods per plant (NPP), 100-grain weight (W100), and grain yield (YIELD) were significantly affected the treatments (
Table 2). In contrast, the number of grains per pod (NGP) and the final stand (FS) variables were not influenced by the treatments.
Higher averages for the number of pods per plant (NPP) parameter were found in the inoculation treatments, but they did not differ, especially for inoculation via seed (T2) and inoculation via seed with reinoculation at the V4 stage (T3) (
Figure 7), which were statistically equivalent to the treatment with mineral nitrogen fertilizer (T10).
Regarding 100-grain weight (W100), despite being a variable that suffers little influence from the environment, that is, with variation depending on the plant genotype and the NGP, the average values found in this study ranged between 13.0 g and 27.3 g for the treatments that did not involve inoculation (T1) and T3, respectively, with the latter being statistically equivalent to T10, the treatment involving mineral nitrogen fertilization (
Figure 8).
Following the same trend of its components, the highest grain yield was obtained when inoculation via seed was followed by reinoculation at the V4 stage (T3), which made it possible to obtain a yield of 3.271 kg ha
−1, the highest found in this study, being 15% higher than treatment with nitrogen formula (T10) but not statistically different from this treatment (
Figure 9).
4. Discussion
It is known that the benefits of symbiosis in bean plants begin between 15 and 20 days after sowing and that there is a decrease in biological nitrogen fixation (BNF) activity from the R5 stage, where reinoculation can promote the maintenance of BNF in periods when this activity begins to decline [
15,
16]. Regarding T1, the population of native bacteria in the soil predominated, with low atmospheric N2 fixation potential given the low number of nodules produced in that treatment (statistically equal to T10, the treatment involving the use of a mineral nitrogen fertilizer). Furthermore, as used in the present study, adding mineral N in high doses makes the native fixing bacteria inefficient, thus justifying the low nodulation rate in this treatment.
Based on this context, it can be stated that the non-inoculation treatments showed low nodulation capacity, as previously discussed, contrary to the results reported by the authors of [
17,
18], who observed nodulation in non-inoculated common bean plants and related this to the presence of native rhizobia species in the soil. These results can be justified by the greater responsiveness of current bean genotypes to the use of rhizobia, which is more efficient in fixing atmospheric N
2 than the
R. tropici species but, on the other hand, less attractive to the bacteria native to the soil. It is also worth noting that the cultivar BRS FC402, due to its indeterminate growth habit (type II) and semi-erect architecture, contributed to the success of reinoculation via spraying in topdressing, allowing for the arrival of the product to the soil and subsequently to the root system, enabling the process of infection and the formation of nodules.
Despite the lack of a significant difference between the treatments involving inoculation, the treatments that involving reinoculation in topdressing from the reproductive stage onwards showed a lower nodulation capacity, which can be explained by the proximity of the reinoculations with the phases of greater demand of the plants, which are the flowering and reserve filling phases.
In general, the number of total nodules produced in the present study in the inoculation treatments can be considered high, according to [
19], with an average of 15.2 nodules plant
−1, indicating good symbiotic efficiency. However, it is important to emphasize that although the number of nodules formed was considered large, the dry mass of these nodules should also be analyzed since a significant number of nodules with smaller sizes can limit the efficiency of the BNF because, when very small, properly fixed N may not be able to be returned to the plants [
20].
In a study conducted by Sales [
21], who set out to investigate the ideal time to carry out the reinoculation in topdressing of
Rhizobium tropici in common bean under no-tillage, higher averages were verified for the treatment involving inoculation via seeds with
R. tropici and reinoculation at the V4 or R5 stages, where the reinoculations added to the TN and yielded a greater NDM. This behavior was not observed in this study since the topdressing reinoculations performed in treatments T3 to T9 did not show a cumulative effect on this parameter, and only T2 maintained a higher mean value.
Because of this, it can be seen that the inoculation via seed and reinoculation in V4 allowed for the maintenance of nodules compared to the treatment involving inoculation only via seed. The reinoculations performed at the other bean growth stages were inefficient in promoting more nodules in evaluations at the R7 stage. It is worth noting that the Union of the Network of Laboratories for the Recommendation, Standardization, and Dissemination of Microbial Inoculant Technology of Agricultural Interest (RELARE) uses the evaluation of the number of nodules and dry mass of nodules as one of the criteria for evaluating the symbiotic efficiency between rhizobia and Fabaceae [
22,
23].
As for leaf nitrogen content (NC), according to Rosolem [
24], the translocation of nutrients begins in the period of pod formation, where the N present in the leaves is directed to the formation of pods and later to the filling of grains. As the sampling in this study took place at the R7 stage, characterized as the period of the formation of pods in the common bean (where there is a greater demand for nutrients due to the process of filling grain reserves), NC may have suffered a translocation effect, which occurs with the translocation of the N of the leaf to a higher-concentration place from a place of a lower concentration (grain filling), thus justifying the non-significance of this parameter when analyzed in leaves. The authors of [
25,
26] also did not observe a significant influence on the leaf nitrogen content in common beans when subjected to treatments with inoculation.
The addition of roots can influence bean yield since the tip of the roots and the development of piliferous zones increases the plant’s water absorption and nutrients, helping their development [
27]. The authors of [
28], working with common bean under different inoculant doses based on
R. tropici, obtained mean taproot length (TRL) values close to those found in this study for the same inoculant dosage in analyses carried out at the R6 stage. Since it is the most abundant element in plants and because it participates in processes such as protein synthesis and the formation of important nucleotides for cell metabolism, in addition to being present in the composition of proteins, enzymes, and coenzymes, nitrogen becomes a contributor to the growth of the plants, both in the root system and shoots [
29].
Reinoculation at the V4 stage (T3) and R5 stage (T4) resulted in the highest root dry mass (RDM). This result contradicts what the authors of [
30] found, where the treatment with mineral fertilizer was 25% higher in terms of the same evaluated parameter than the inoculated one. This result could possibly be explained by the greater development of the root system when reinoculation is performed (
Figure 4). It is worth noting that the RDM is an important parameter to observe since a more voluminous root system can provide a greater absorption of nutrients due to the greater contact of the plant’s roots with the soil, thus providing improved plant response regarding yield potential [
31].
In a study conducted by the authors of [
21], it was possible to verify that the reinoculation of
Rhizobium tropici in common bean influenced the shoot dry mass (SDM) parameter, producing an average of 17.9 g plant
−1 in the non-reinoculation treatment, 15.5 g plant
−1 when reinoculated at the V4 stage, and 16.6 g plant
−1 when reinoculated at the R5 stage
The variation found for the final stand (FS) is small and does not influence the bean grain yield, even in the smallest stand, because the bean plants compensate for the existing spaces. Furthermore, an FS between 10 and 12 plants per meter, in combination with a spacing of 0.5 m, allows one to obtain 200 to 220 thousand plants ha
−1, close to the population of 240 thousand plants ha-1 considered suitable for the bean crop (according to what was reported by the authors of [
32]).
The treatments did not affect the number of grains per pod. As this characteristic is more intrinsically linked to the genotypic characteristics, being minimally influenced by the environment, the non-significant result of the treatments applied in this study can be justified, and, normally, these values are between 4 and 5 grains per pod [
33].
The production of pods per plant (NPP) was higher when applying inoculation via an isolated seed (T1) or with reinoculation at stage V4 (T3) and mineral fertilization (T10). Contrary to the results of [
34], supplying N in mineral form, both in sowing and in topdressing, resulted in a higher NPP compared to the inoculated treatment. On the other hand, the control treatment (T1) provided the lowest average of NPP, confirming that the native bacteria in the soil could not supply the necessary N for the development of the bean crop. The satisfactory supply of N via the BNF process, with emphasis on treatments 2 and 3, certainly enabled the supply of this nutrient in adequate amounts, ensuring a greater production of flowers and, consequently, greater production of pods, corroborating what was reported by the authors of [
15].
The highest NPP found in these treatments (around 24 pods per plant) is higher than the average value found by the authors of [
35] in their study involving inoculation using
Rhizobium for the cultivar Carioca, where they obtained an average of 22 pods per plant. It should be noted that, among the yield components, NPP is the one that most influences bean grain yield [
23]. Furthermore, in a study carried out by the authors of [
36] with eight cultivars of common bean and two strains of
Rhizobium, it was possible to verify the dependence of the strain used on the number of pods per plant. In contrast, Ferreira et al. [
37] state that the number of pods per plant is more closely related to the cultivar and that it does not suffer interference from the strain used in the inoculation. It is worth noting that, regardless of the choice of strain and cultivar in the NPP analysis, we can observe significant increases in this parameter when inoculation is performed, ensuring statistical equivalence with mineral nitrogen fertilization.
It is worth noting that the treatments that involved inoculation and T10 presented average values of 25 g, close to the parameter of cultivar BRS FC402, which is 26 g. According to the authors of [
38], this small difference can be attributed to the management employed and the environmental conditions, which can influence the final result for this parameter. According to the authors of [
39], all yield components must reach their maximum levels and proper balance to achieve maximum bean grain yield and its adequate balance.
Compared to the control treatment (T1), which did not involve inoculation or nitrogen mineral fertilization, T3 resulted in an increase of 2.644 kg ha
−1, confirming the efficiency of the use of inoculation to increase bean grain yield. Pelegrin et al. [
4] studied grain productivity from beans cv. Pérola of 3.339 kg ha
−1 in a treatment that involved inoculation with rhizobia via seed (strain SEMIA 4077), equivalent to the application of 80 kg ha
−1 of N and therefore close to the average productivity obtained in this study. Similarly, in a study carried out by Sousa et al. [
40] with beans cv. BRS Valente, a productivity of 3.976 kg ha
−1 was obtained for the treatment that involved cover inoculation + supplementary cover reinoculation carried out at the V4 stage, corroborating the results expressed in this work and confirming the efficiency of the inoculation process in the context of beans when combined with seed inoculation and cover reinoculation carried out at the V4 stage. It is worth noting that the price of inoculants based on
R. tropici for beans currently in Brazil corresponds to less than half of the costs of purchasing nitrogenous mineral fertilizers (such as urea) per hectare, thus confirming that reductions in production costs could be realized via the use of rhizobial inoculants in bean crops.
Based on the results obtained in this study, it can be stated that the bean plant’s nitrogen demand can be completely met by adding an inoculant via seed and re-inoculation under cover at the V4 stage, with this technique being more sustainable than the use of nitrogenous mineral fertilizers in terms of the costs and environmental constraints associated with both.




 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}