
Effects of Continuous Tomato Monoculture on Soil Microbial Properties and Enzyme Activities in a Solar Greenhouse

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Experimental Design
2.2. Soil Sampling and Analysis
2.2.1. Soil Sampling
2.2.2. Analysis of Soil Microbial Abundance
2.2.3. Analysis of the Functional Diversity of the Soil Microbial Community
2.2.4. Determination of Soil Chemical Properties
2.2.5. Determination of Soil Enzyme Activities
2.2.6. Measurement of Average Plant Yield
2.3. Statistical Analysis
3. Results
3.1. Soil Microbial Abundance
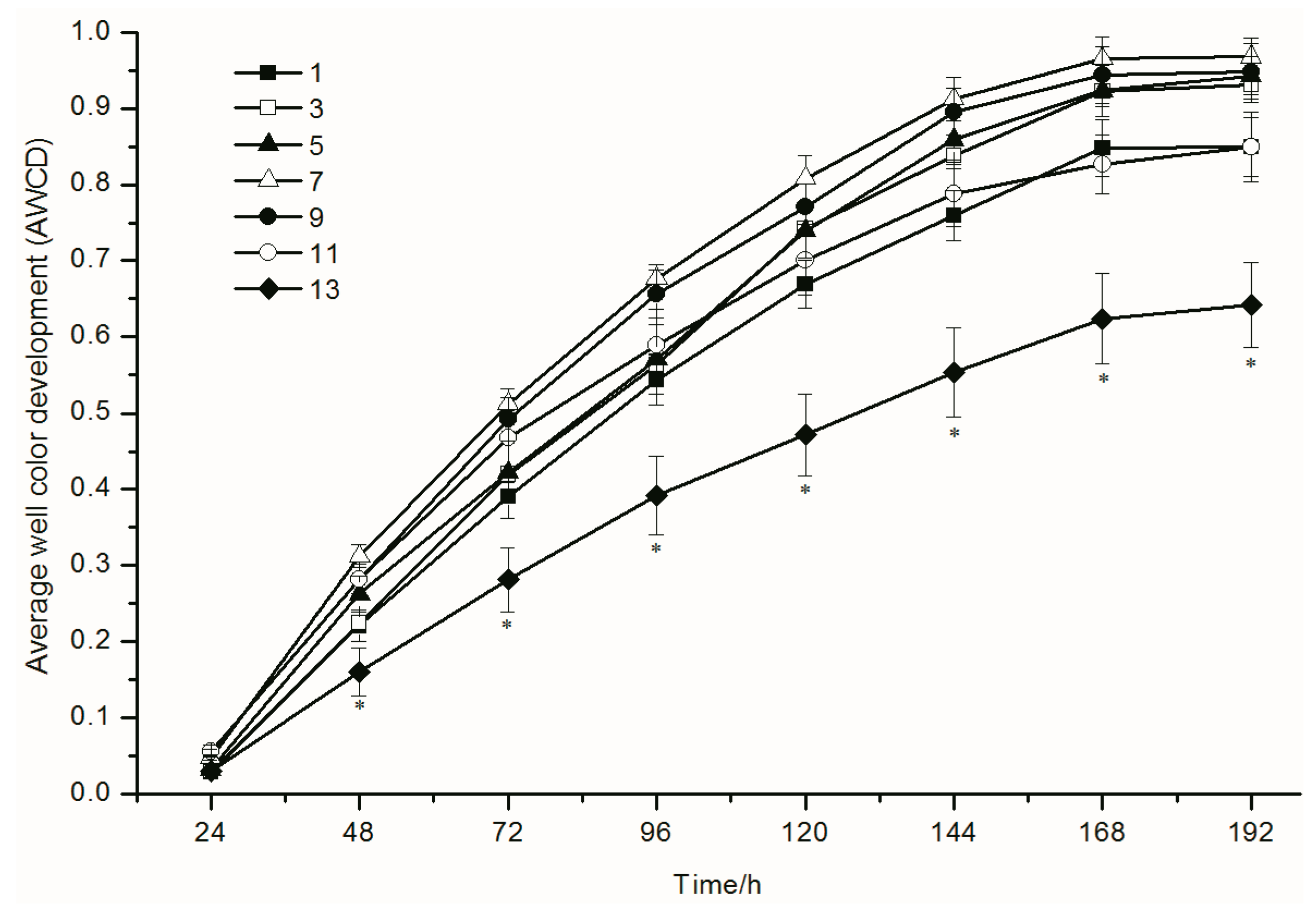
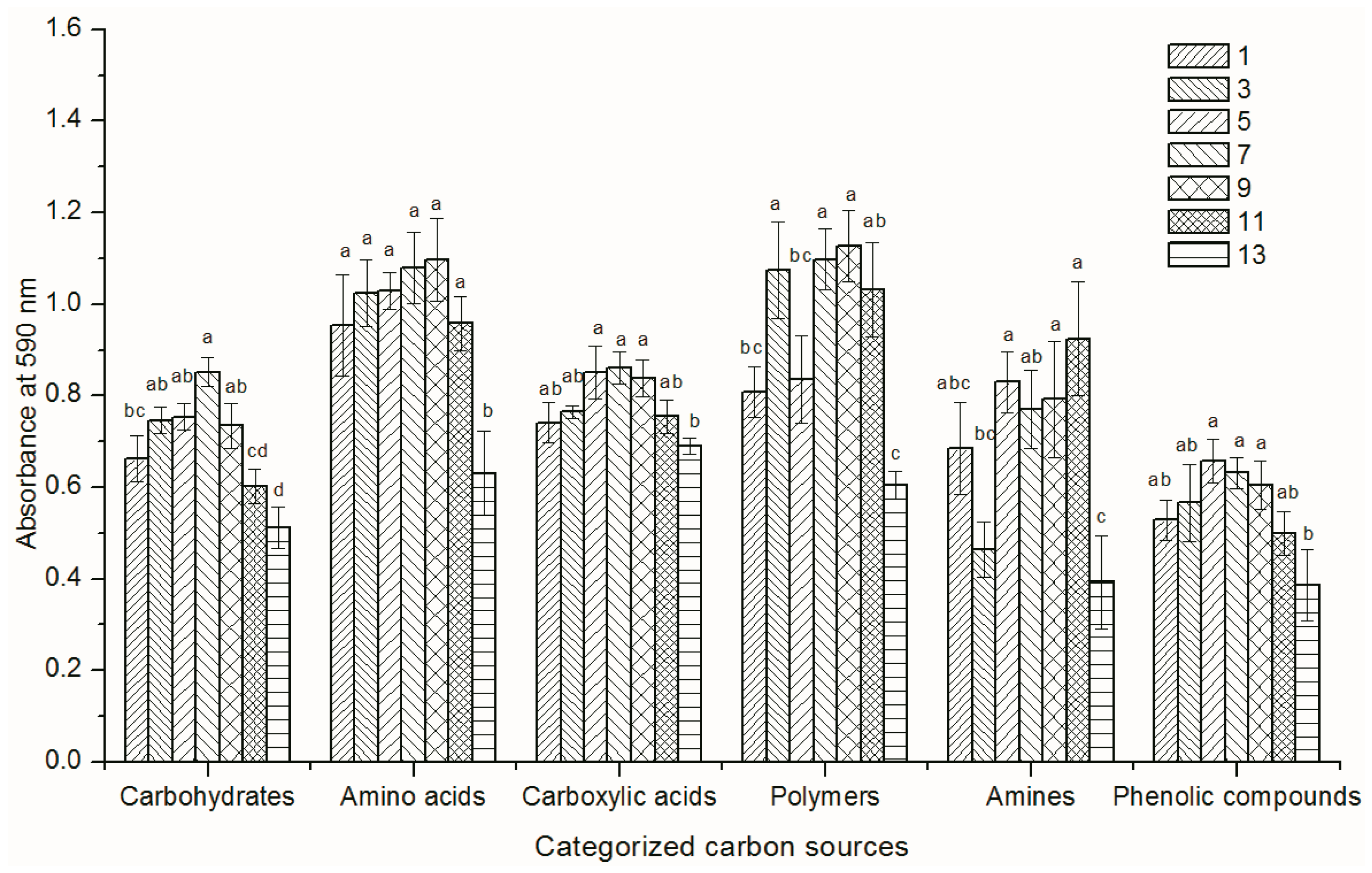
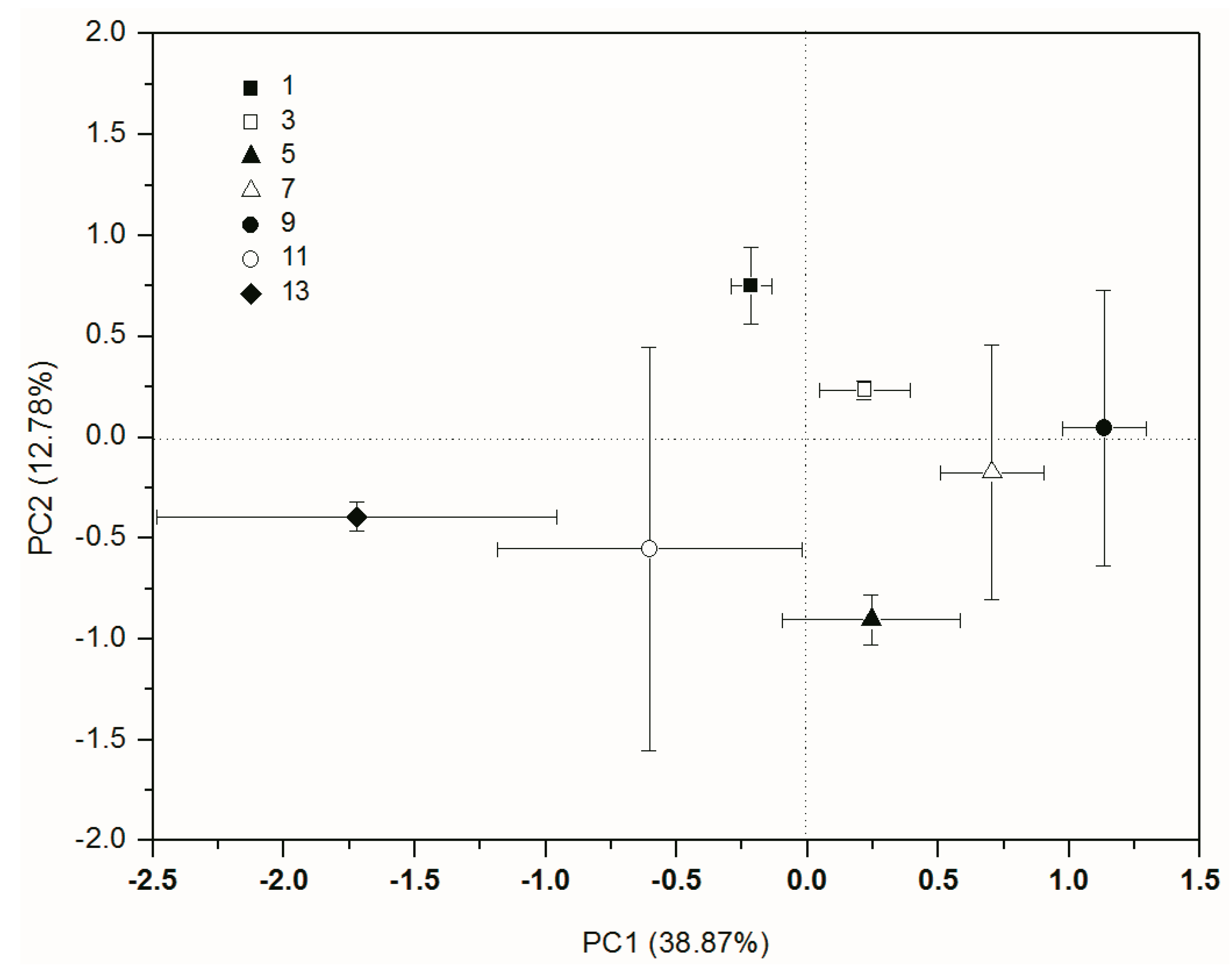
3.2. Functional Diversity of the Soil Microbial Community
3.3. Soil Chemical Properties
3.4. Soil Enzyme Activities
3.5. Correlations of Soil Quality Indicators
3.6. Tomato Yield
3.7. Correlations between Tomato Yield and Soil Quality Indicators
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Panwar, N.L.; Kaushik, S.C.; Kothari, S. Solar greenhouse an option for renewable and sustainable farming. Renew. Sustain. Energy Rev. 2011, 15, 3934–3945. [Google Scholar] [CrossRef]
- Chai, L.; Ma, C.; Ni, J.Q. Performance evaluation of ground source heat pump system for greenhouse heating in northern China. Biosyst. Eng. 2012, 111, 107–117. [Google Scholar] [CrossRef]
- More, T.A.; Chandra, P.; Majumdar, G.; Singh, J.K. Some observation on growing cucumber under greenhouse. In Proceedings of the 11th International Congress on the Use of Plastics in Agriculture, New Delhi, India, 26 February–2 March 1990; pp. 49–55.
- Nimje, P.M.; Wanjari, O.P.; Shyam, M. Greenhouse technology for vegetable crop production. In Proceedings of the 11th International Congress on the Use of Plastics in Agriculture, New Delhi, India, 26 February–2 March 1990; pp. 83–90.
- Han, J.; Shi, J.; Zeng, L.; Xu, J.; Wu, L. Effects of nitrogen fertilization on the acidity and salinity of greenhouse soils. Environ. Sci. Pollut. Res. 2015, 22, 2976–2986. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.H.; Liu, X.J.; Zhang, Y.; Shen, J.L.; Han, W.X.; Zhang, W.F.; Christie, P.; Goulding, K.W.T.; Vitousek, P.M.; Zhang, F.S. Significant Acidification in Major Chinese Croplands. Science 2010, 327, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.G.; Yin, R.; Zhang, H.Y.; Huang, J.F.; Chen, R.R.; Cao, Z.H. Changes of soil microbiological properties caused by land use changing from rice-wheat rotation to vegetable cultivation. Environ. Geochem. Health 2004, 26, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shao, X.H.; Chang, T.T. Subsurface drainage improved the secondary soil salinity in plastic greenhouse. J. Food Agric. Environ. 2012, 10, 1409–1412. [Google Scholar]
- Rakesh, K.; Ram, C.N.; Yadav, G.C.; Deo, C.; Vimal, S.C.; Bhartiya, H.D. Studies on correlation and path coefficient analysis in tomato (Solanumlycopersicon L.). Plant Arch. 2014, 14, 443–447. [Google Scholar]
- Xu, H.L. Introduction and application of TM2-NV gene of tomato resources resistant to tobacco mosaix virus (TMV). In Proceedings of the International Symposium on Cultivar Improvement of Horticultural Crops, Part 1: Vegetable Crops, Beijing, China, 6–10 September 1993.
- Zhao, P.Y. Tomato production situation and development countermeasure in Liaoning province. China Veg. 2015, 10, 6–9. [Google Scholar]
- Sebastian, D.; Hauck, M.; Daniel, S.; Dörfler, I.; Hühne, R.; Bade, C. Separating forest continuity from tree age effects on plant diversity in the ground and epiphyte vegetation of a Central European mountain spruce forest. Flora Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 238–246. [Google Scholar]
- Zhou, X.G.; Wu, F.Z. Dynamics of the diversity of fungal and fusarium communities during continuous cropping of cucumber in the greenhouse. FEMS Microbiol. Ecol. 2012, 80, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.G.; Yu, G.B.; Wu, F.Z. Effects of intercropping cucumber with onion or garlic on soil enzyme activities, microbial communities and cucumber yield. Eur. J. Soil Biol. 2011, 47, 279–287. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, J.L.; Gu, T.Y.; Zhang, W.M.; Shen, Q.R.; Yin, S.X.; Qi, H.Z. Microbial community diversities and taxa abundances in soils along a seven-year gradient of potato monoculture using high throughput pyrosequencing approach. PLoS ONE 2014, 9, e86610. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.G.; Yu, G.B.; Wu, F.Z. Soil phenolics in a continuously mono-cropped cucumber (Cucumissativus L.) system and their effects on cucumber seedling growth and soil microbial communities. Eur. J. Soil Sci. 2012, 63, 332–340. [Google Scholar] [CrossRef]
- Xiong, W.; Li, Z.G.; Liu, H.J.; Xue, C.; Zhang, R.F.; Wu, H.S.; Li, R.; Shen, Q.R. The effect of long-term continuous cropping of black pepper on soil bacterial communities as determined by 454 pyrosequencing. PLoS ONE 2015, 10, e0136946. [Google Scholar] [CrossRef] [PubMed]
- You, C.H.; Jiang, L.F.; Xi, F.H.; Wang, W.W.; Li, M.J.; Xu, Z. Comparative evaluation of different types of soil conditioners with respect to their ability to remediate consecutive tobacco monoculture soil. Int. J. Agric. Biol. 2015, 17, 969–975. [Google Scholar] [CrossRef]
- Zhang, H.J.; Ding, W.X.; He, X.H.; Yu, H.Y.; Fan, J.L.; Liu, D.Y. Influence of 20-year organic and inorganic fertilization on organic carbon accumulation and microbial community structure of aggregates in an intensively cultivated Sandy loam Soil. PLoS ONE 2014, 9, e92733. [Google Scholar] [CrossRef] [PubMed]
- Buller, S.; Inglis, D.; Miles, C. Plant growth, fruit yield and quality, and tolerance to verticillium wilt of grafted watermelon and tomato in field production in the Pacific Northwest. HortScience 2013, 48, 1003–1009. [Google Scholar]
- Sam, C.; Freitas, S.S.; Airoldi, C. Microcalorimetric measurements of the metabolic activity by bacteria and fungi in some Brazilian soils amended with different organic matter. Thermochim. Acta 2004, 417, 275–281. [Google Scholar]
- Green, J.; Bohannan, B.J. Spatial scaling of microbial biodiversity. Trends Ecol. Evol. 2006, 21, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Da, W.; Ghani, A. Why is the strength of relationships between pairs of methods for estimating soil microbial biomass often so variable? Soil Biol. Biochem. 1995, 27, 821–828. [Google Scholar]
- Du, Z.M.; Xie, Y.; Hu, L.Q.; Hu, L.X.; Xu, S.D.; Li, D.X.; Wang, G.F.; Fu, J.M. Effects of fertilization and clipping on carbon, nitrogen storage, and soil microbial activity in a natural grassland in Southern China. PLoS ONE 2014, 9, e99385. [Google Scholar] [CrossRef] [PubMed]
- Jpo, E.; Liu, Y.; Chan, J.W.; Downes, D. Cucumber productivity and soil degradation in recropping system in greenhouse. Commun. Soil Sci. Plant Anal. 2012, 43, 1743–1748. [Google Scholar]
- Cardinale, B.J.; Srivastava, D.S.; Emmett, D.J.; Wright, J.P.; Downing, A.L.; Sankaran, M.; Jouseau, C. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 2006, 443, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.L.; Paixão, S.M.; Caçador, I.; Carolino, M. CLPP and EEA profiles of microbial communities in salt marsh sediments. J. Soils Sediments 2007, 7, 418–425. [Google Scholar] [CrossRef]
- Islam, M.R.; Chauhan, P.S.; Kim, Y.; Kim, M.; Sa, T. Community level functional diversity and enzyme activities in paddy soils under different long-term fertilizer management practices. Biol. Fertil. Soils 2011, 47, 599–604. [Google Scholar] [CrossRef]
- Dick, R.P.; Doran, J.W.; Coleman, D.C.; Bezdicek, D.F.; Stewart, B.A. Soil enzymes activities as indicators of soil quality. In Defining Soil Quality for a Sustainable Environment; Soil Science Society of America, Inc.: Madison, WI, USA, 1994; pp. 107–124. [Google Scholar]
- Zhou, X.G.; Wu, F.Z. Changes in soil chemical characters and enzyme activities during continuous monocropping of cucumber (Cucumissativus). Pak. J. Bot. 2015, 47, 691–697. [Google Scholar]
- Xiao, X.M.; Cheng, Z.H.; Meng, H.W.; Khan, M.A.; Li, H.Z. Intercropping with garlic alleviated continuous cropping obstacle of cucumber in plastic tunnel. Acta Agric. Scand. Sect. B Soil Plant Sci. 2012, 62, 696–705. [Google Scholar] [CrossRef]
- Garland, J.L.; Mills, A.L.; Young, J.S. Relative effectiveness of kinetic analysis vs. single point readings for classifying environmental samples based on community-level physiological profiles (CLPP). Soil Biol. Biochem. 2001, 33, 1059–1066. [Google Scholar] [CrossRef]
- Rogers, B.F.; Tate, R.L., III. Temporal analysis of the soil microbial community along a toposequence in pineland soils. Soil Biol. Biochem. 2001, 33, 1389–1401. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agro-Chemical Analysis; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Zhou, J.; Guo, W.H.; Wang, R.Q.; Han, X.M.; Wang, Q. Microbial community diversity in the profle of an agricultural soil in northern China. J. Environ. Sci. 2008, 20, 981–988. [Google Scholar] [CrossRef]
- Dong, M.; Wang, Y.F.; Kong, F.Z.; Jiang, G.M.; Zhang, Z.B. Survey Observation and Analysis of Terrestrial Biocommunities; Beijing Standards Press: Beijing, China, 1996; pp. 130–132. [Google Scholar]
- Yan, C.S. Research Methods of Soil Fertility; China Agriculture Press: Beijing, China, 1988; pp. 243–280. [Google Scholar]
- Wu, Z.Y.; Haack, S.E.; Lin, W.; Li, B.L.; Wu, L.K.; Fang, C.X.; Zhang, Z.X. Soil microbial community structure and metabolic activity of Pinuselliottii plantations across different stand ages in a subtropical area. PLoS ONE 2015, 10, e0135354. [Google Scholar]
- Corneo, P.E.; Pellegrini, A.; Cappellin, L.; Roncador, M.; Chierici, M.; Gessler, C.; Pertot, I. Microbial community structure in vineyard soils across altitudinal gradients and in different seasons. FEMS Microbiol. Ecol. 2013, 84, 588–602. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.Y.; Xu, W.L.; Mao, J.; Zhang, Z.D.; Tang, G.M.; Ge, C.H. Microbial community diversity of rhizosphere soil in continuous cotton cropping system in Xinjiang. ActaEcol. Sin. 2012, 32, 3031–3040. [Google Scholar]
- Ma, Y.H.; Wei, M.; Wang, X.F. Variation of microflora and enzyme activity in continuous cropping cucumber soil in solar greenhouse. Chin. J. Appl. Ecol. 2004, 15, 1005–1008. [Google Scholar]
- Nie, L.X.; Peng, S.B.; Bouman, B.A.M.; Huang, J.L.; Cui, K.H.; Visperas, R.M.; Xiang, J. Alleviating soil sickness caused by aerobic monocropping: Responses of aerobic rice to various nitrogen sources. Soil Sci. Plant Nutr. 2009, 55, 150–159. [Google Scholar] [CrossRef]
- Wang, W.J.; Su, D.X.; Qiu, L.; Wang, H.Y.; An, J.; Zheng, G.Y. Concurrent changes in soil inorganic and organic carbon during the development of larch, Larixgmelinii, plantations and their effects on soil physicochemical properties. Environ. Earth Sci. 2013, 69, 1559–1570. [Google Scholar] [CrossRef]
- Lu, G.; He, G.X.; Liu, W.G. Long-term cropping effects on agricultural sustainability in Alar Oasis of Xinjiang. Sustainability 2016, 8, 61. [Google Scholar] [CrossRef]
- Sun, S.S.; Feng, X.S.; Wan, S.B.; Zuo, X.Q. Changes of main microbial strains and enzymes activities in peanut continuous cropping soil and their interactions. Acta Agron. Sin. 2001, 27, 617–621. [Google Scholar]
- Sun, R.L.; Zhao, B.Q.; Zhang, L.S. Effects of long term fertilization on soil enzyme activities and its role in adjusting-controlling soil fertility. Plant Nutr. Fertil. Sci. 2003, 9, 406–410. [Google Scholar]
- Frankenberger, W.T.; Dick, W.A. Relationships between enzyme activities and microbial growth and activity indices in soil. Soil Sci. Soc. Am. J. 1983, 47, 945–951. [Google Scholar] [CrossRef]
- Mungai, N.W.; Motavalli, P.P.; Kremer, R.J.; Nelson, K.A. Spatial variation of soil enzyme activities and microbial functional diversity in temperate alley cropping systems. Biol. Fertil. Soils 2005, 42, 129–136. [Google Scholar] [CrossRef]
- Makoi, J.; Ndakidemi, P.A. Selected soil enzymes: Examples of their potential roles in the ecosystem. Afr. J. Biotechnol. 2008, 7, 181–191. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | EC (µs·cm−1) | AvailableNitrogen (AN, mg·kg−1) | Available Phosphorus (AP, mg·kg−1) | Available Potassium (AK, mg·kg−1) | Organic Matter (OM, g·kg−1) |
|---|---|---|---|---|---|
| 7.04 | 290.30 | 95.81 | 94.65 | 255.70 | 20.30 |
| Crop | TMA (×106) | BAC (×106) | FUN (×103) | ACT (×105) | B/F Value |
|---|---|---|---|---|---|
| 1 | 3.19 ± 0.23 bc | 3.01 ± 0.21 bc | 8.84 ± 0.14 b | 1.65 ± 0.28 a | 341.71 bc |
| 3 | 3.23 ± 0.41 bc | 3.09 ± 0.39 bc | 9.41 ± 0.42 b | 1.43 ± 0.17 a | 330.89 bc |
| 5 | 4.82 ± 1.13 ab | 4.72 ± 1.10 ab | 9.80 ± 0.58 b | 1.25 ± 0.38 a | 489.44 ab |
| 7 | 6.24 ± 0.40 a | 6.12 ± 0.42 a | 9.30 ± 0.82 b | 1.15 ± 0.27 a | 641.73 a |
| 9 | 5.97 ± 0.82 a | 5.81 ± 0.84 a | 8.81 ± 0.34 b | 1.52 ± 0.31 a | 667.63 a |
| 11 | 4.83 ± 0.61 ab | 4.67 ± 0.58 ab | 10.15 ± 0.60 b | 1.55 ± 0.62 a | 468.47 abc |
| 13 | 2.45 ± 0.42 c | 2.18 ± 0.38 c | 12.08 ± 0.75 a | 2.51 ± 0.55 a | 183.69 c |
| Crop | S | H | D | U |
|---|---|---|---|---|
| 1 | 25.11 ± 0.77 a | 3.13 ± 0.02 a | 0.95 ± 0.00 ab | 5.23 ± 0.17 b |
| 3 | 26.14 ± 0.63 a | 3.18 ± 0.02 a | 0.96 ± 0.00 a | 5.54 ± 0.11 ab |
| 5 | 26.43 ± 0.57 a | 3.21 ± 0.02 a | 0.96 ± 0.00 a | 5.60 ± 0.14 ab |
| 7 | 26.57 ± 0.65 a | 3.20 ± 0.02 a | 0.96 ± 0.00 a | 6.02 ± 0.13 a |
| 9 | 26.67 ± 0.53 a | 3.12 ± 0.02 a | 0.96 ± 0.00 a | 5.86 ± 0.17 a |
| 11 | 24.11 ± 0.79 a | 3.09 ± 0.03 a | 0.95 ± 0.00 ab | 5.48 ± 0.23 ab |
| 13 | 19.89 ± 1.24 b | 2.88 ± 0.13 b | 0.95 ± 0.01 b | 4.39 ± 0.23 c |
| Crop | pH | OM (g·kg−1) | AN (mg·kg−1) | AP (mg·kg−1) | AK (mg·kg−1) |
|---|---|---|---|---|---|
| 1 | 6.88 ± 0.01 abc | 28.40 ± 3.93 bc | 168.59 ± 2.26 d | 150.48 ± 2.64 d | 304.45 ± 6.17 d |
| 3 | 6.94 ± 0.06 ab | 28.57 ± 1.44 bc | 198.64 ± 1.31 bc | 186.53 ± 5.35 bc | 363.65 ± 10.54 c |
| 5 | 6.96 ± 0.03 a | 33.35 ± 0.55 ab | 192.87 ± 5.76 bc | 206.52 ± 9.09 b | 485.29 ± 5.98 b |
| 7 | 6.87 ± 0.03 abc | 37.62 ± 1.87 a | 183.01 ± 2.60 cd | 211.40 ± 7.87 b | 484.36 ± 2.86 b |
| 9 | 6.87 ± 0.01 abc | 37.48 ± 1.33 a | 220.75 ± 8.59 a | 328.91 ± 15.13 a | 521.38 ± 16.12 a |
| 11 | 6.75 ± 0.08 cd | 30.23 ± 1.64 bc | 203.68 ± 5.41 ab | 203.98 ± 10.42 b | 357.73 ± 13.98 c |
| 13 | 6.63 ± 0.09 d | 25.52 ± 1.27 c | 122.21 ± 9.25 e | 170.45 ± 5.98 cd | 236.03 ± 3.12 e |
| Crop | URE (mg NH3-N/g·24 h) | INV (mg glucose/g·24 h) | N-PHO (mg phenol/g·24 h) | CAT (mg KMnO4/g·20 min) |
|---|---|---|---|---|
| 1 | 0.80 ± 0.03 b | 12.12 ± 1.79 c | 1.35 ± 0.04 ab | 0.61 ± 0.01 a |
| 3 | 0.92 ± 0.03 b | 12.38 ± 0.20 c | 1.34 ± 0.06 ab | 0.59 ± 0.02 a |
| 5 | 0.99 ± 0.09 b | 17.27 ± 0.49 ab | 1.30 ± 0.03 ab | 0.56 ± 0.02 ab |
| 7 | 1.00 ± 0.01 b | 18.72 ± 1.32 a | 1.34 ± 0.16 ab | 0.53 ± 0.05 ab |
| 9 | 1.45 ± 0.22 a | 18.25 ± 1.96 a | 1.51 ± 0.07 a | 0.52 ± 0.03 ab |
| 11 | 0.86 ± 0.09 b | 13.12 ± 0.72 bc | 1.17 ± 0.09 b | 0.47 ± 0.04 b |
| 13 | 0.38 ± 0.02 c | 10.19 ± 1.63 c | 0.57 ± 0.05 c | 0.46±0.05 b |
| URE | INV | N-PHO | CAT | pH | OM | AN | AP | AK | |
|---|---|---|---|---|---|---|---|---|---|
| AWCD | 0.87 * | 0.84 * | 0.92 ** | 0.44 | 0.67 | 0.84 * | 0.86 * | 0.54 | 0.89 ** |
| TMA | 0.79 * | 0.93 ** | 0.62 | −0.11 | 0.18 | 0.96 ** | 0.68 | 0.72 | 0.90 ** |
| BAT | 0.79 * | 0.94 ** | 0.63 | −0.09 | 0.20 | 0.97 ** | 0.69 | 0.71 | 0.91 ** |
| FUN | −0.79 * | −0.56 | −0.98 ** | −0.73 | −0.66 | −0.60 | −0.75 | −0.37 | −0.62 |
| ACT | −0.7 | −0.74 | −0.86 * | −0.51 | −0.80 * | −0.71 | −0.78 * | −0.28 | −0.79 * |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, H.; Zhang, G.; Zhang, F.; Sun, Z.; Geng, G.; Li, T. Effects of Continuous Tomato Monoculture on Soil Microbial Properties and Enzyme Activities in a Solar Greenhouse. Sustainability 2017, 9, 317. https://doi.org/10.3390/su9020317
Fu H, Zhang G, Zhang F, Sun Z, Geng G, Li T. Effects of Continuous Tomato Monoculture on Soil Microbial Properties and Enzyme Activities in a Solar Greenhouse. Sustainability. 2017; 9(2):317. https://doi.org/10.3390/su9020317
Chicago/Turabian StyleFu, Hongdan, Guoxian Zhang, Fan Zhang, Zhouping Sun, Guoming Geng, and Tianlai Li. 2017. "Effects of Continuous Tomato Monoculture on Soil Microbial Properties and Enzyme Activities in a Solar Greenhouse" Sustainability 9, no. 2: 317. https://doi.org/10.3390/su9020317
APA StyleFu, H., Zhang, G., Zhang, F., Sun, Z., Geng, G., & Li, T. (2017). Effects of Continuous Tomato Monoculture on Soil Microbial Properties and Enzyme Activities in a Solar Greenhouse. Sustainability, 9(2), 317. https://doi.org/10.3390/su9020317