1. Introduction
Coastal natural resources, particularly those in low-lying floodplains, are threatened by the most certain, immediate, and widespread result of anthropogenic climate change—accelerated sea-level rise [
1,
2]. The global sea level rose at an average rate of 3.2 mm yr
−1 between 1993 and 2010 [
3], which was up from the rate of 2.0 mm yr
−1 between 1971 and 2010. Compounding the chronic inundation due to tides, storm surges flood coastal ecosystems with increasing frequency, leading to an increase in the salinity and saturation of soils in these habitats, with diminished time between sequential disturbance [
4,
5]. Ecosystems that are unable to migrate landward will be most impacted, resulting in a loss of species diversity [
2,
6] and of the benefits to society that these areas offer. While this element of coastal vulnerability has received a great deal of attention, others have also reported degradation and die-off of coastal vegetation, such as mangroves, in recent years as a result of both chronic stress and acute events [
7].
The response of coastal vegetation to sea-level rise depends on the local relative rate of sea-level change, sediment loading, freshwater hydrology, ability to migrate, and other factors. Donnelly and Bertness [
8] studied the effects of sea-level rise on coastal wetlands in New England, using quadrats to evaluate percent vegetation cover change for two saltmarsh species from 1995 to 1999. In this five-year period alone, both sites experienced a five-fold increase and significant landward migration of cordgrass at the expense of high-marsh species. This coincided with sea-level rise acceleration, as measured at a nearby tide gauge. The authors attributed the increase in cordgrass dominance to region-wide sea-level rise, and connected it to an increased frequency of marsh surface flooding. In the Florida Keys, USA, Ross et al. [
6] found that the salinization of groundwater, caused by a 15 cm rise in the local sea level, was a major factor in the loss of more than half of the pine forest covered area on Sugarloaf Key over a 70 year period. Koch et al. [
9] projected, from regional modelling, future coastal Florida vegetation responses using a sea-level rise estimate of 46 cm by 2060. They predicted that, based on estimated rates of historical sediment accumulation in the region, coastal mangrove forests will not be able to accrete faster than the sea-level rise, and therefore will effectively drown.
Sea-level rise is often studied with regard to the potential inundation of heavily populated metropolitan areas because of the potential for economic devastation. Many cities (e.g., Miami, New York City, and others) are preparing actively to mitigate short-term sea-level rise effects. It is also important to characterize how the rising sea level is impacting low-lying coastal conservation areas.
The Big Bend region of Florida is the largest remaining stretch of undeveloped coastline in the continental United States. Geselbracht et al. [
10] used a Sea Level Affecting Marshes Model (SLAMM) of the southern Big Bend to predict a loss of over 24,000 ha (49%) of coastal forest by 2100, under a scenario of a sea-level rise of one meter. Langston et al. [
11] recently published the results of over 20 years of in-situ coastal forest evaluations in the Big Bend. They observed decreased coastal forest species richness, and decreased
Sabal palmetto (among other species) regeneration and survival, from 1992 to 2014. They attributed the rapid declines to increased tidal flooding that was caused by sea-level rise, as determined by observed and modeled weekly flooding events. Lewis et al. [
7] state that mangrove die-offs in protected areas like South Florida constitute a “heart-attack” dynamic, whereby previous changes to local or regional hydrology have created chronic stress that alone would ultimately lead to eventual ecosystem decline, but are compounded by acute events, such as hurricanes, which may accelerate the decline of the now vulnerable habitats, decrease resilience, and preclude recovery.
A decades-long time series of synoptic observations, collected by a suite of satellite sensors, can be used to better quantify changes estimated by previous modeling efforts [
12]. The Landsat mission series, for example, has acquired optical data of the globe since the early 1980’s at a spatial resolution of 30 m. More recently, very-high-spatial-resolution (i.e., <5 m) satellite data have been collected, and compliment the longer time series with more precise observations of land change.
In this study, we used time series of satellite images and aerial photography to assess change in coastal forests of the Big Bend region of Florida from 1982 to 2017. Our working hypothesis was that the rapid decline of vegetation reported by Langston et al. [
11] as a result of sea-level-rise is exacerbated by one or more acute meteorological events, including extreme cold temperatures that drove some of the vegetation over a threshold and into rapid decline, from which it has yet to recover.
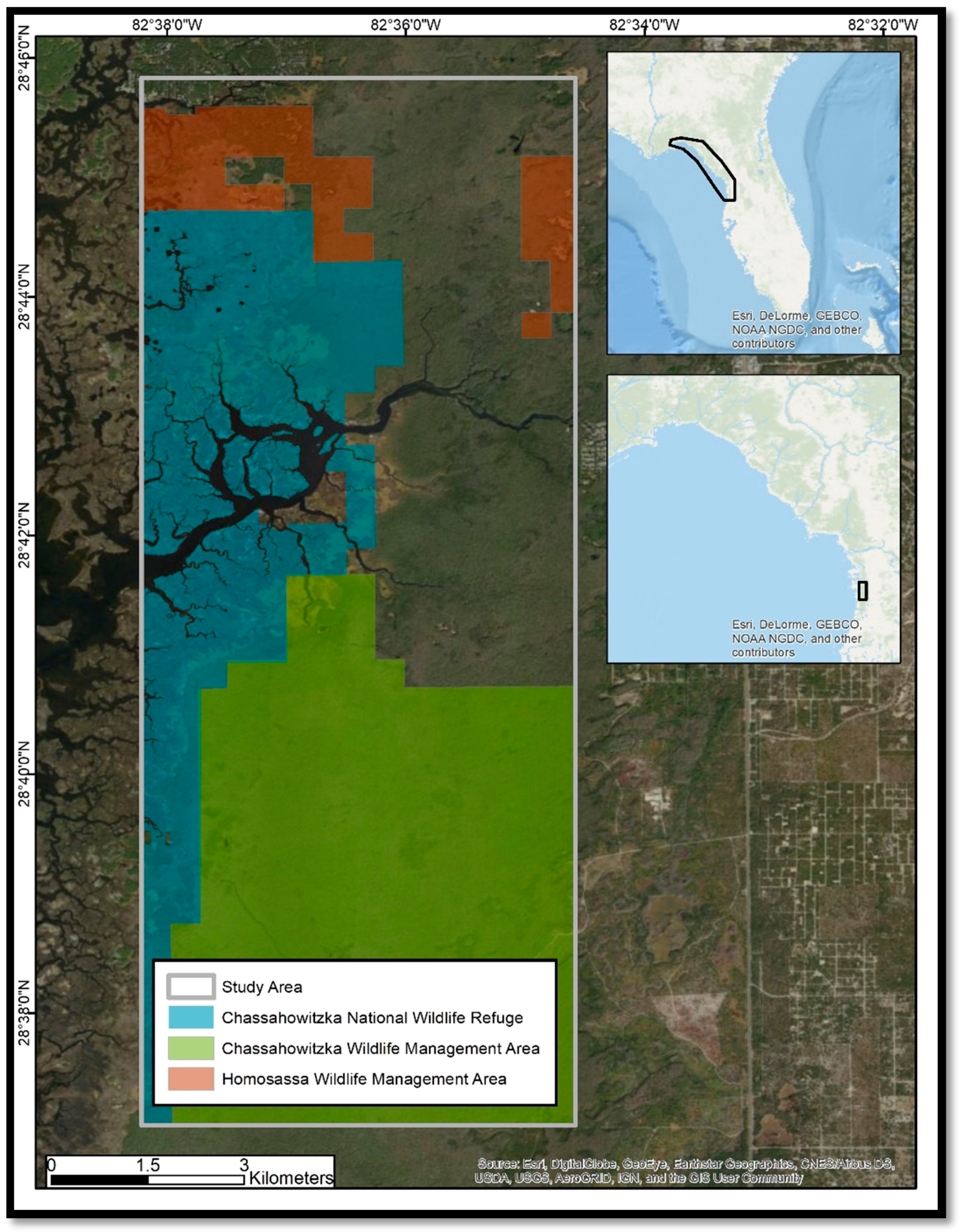
The study area intersects three management areas: The Chassahowitzka National Wildlife Refuge (CNWR), the Chassahowitzka Wildlife Management Area, and the Homosassa Wildlife Management Area (
Figure 1). Swampland extends from the coast, approximately 8 km inland, and ranges in elevation from 0 to less than 3 m. Much of this ecosystem is tidally influenced, and fed upstream by a series of freshwater springs. Urban and other human development is sparse in this region. The natural vegetation inland of the tidal marsh is primarily mixed hardwoods, including
Acer floridanum,
Persea borbonia,
Quercus virginiana, and
Sabal palmetto, which vary in their tolerance to salinity and cold temperatures. Much of the swamp area consists of poorly drained organic soils atop limestone of the Floridan aquifer system [
13].
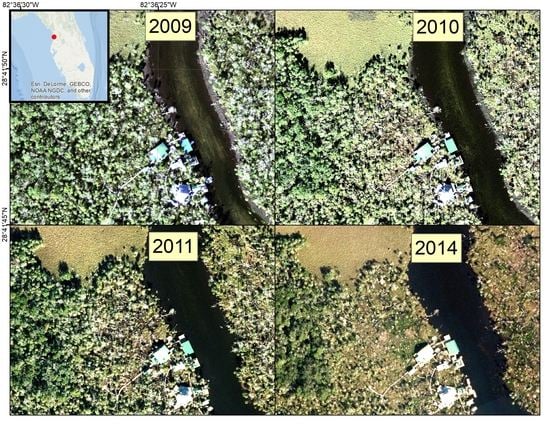
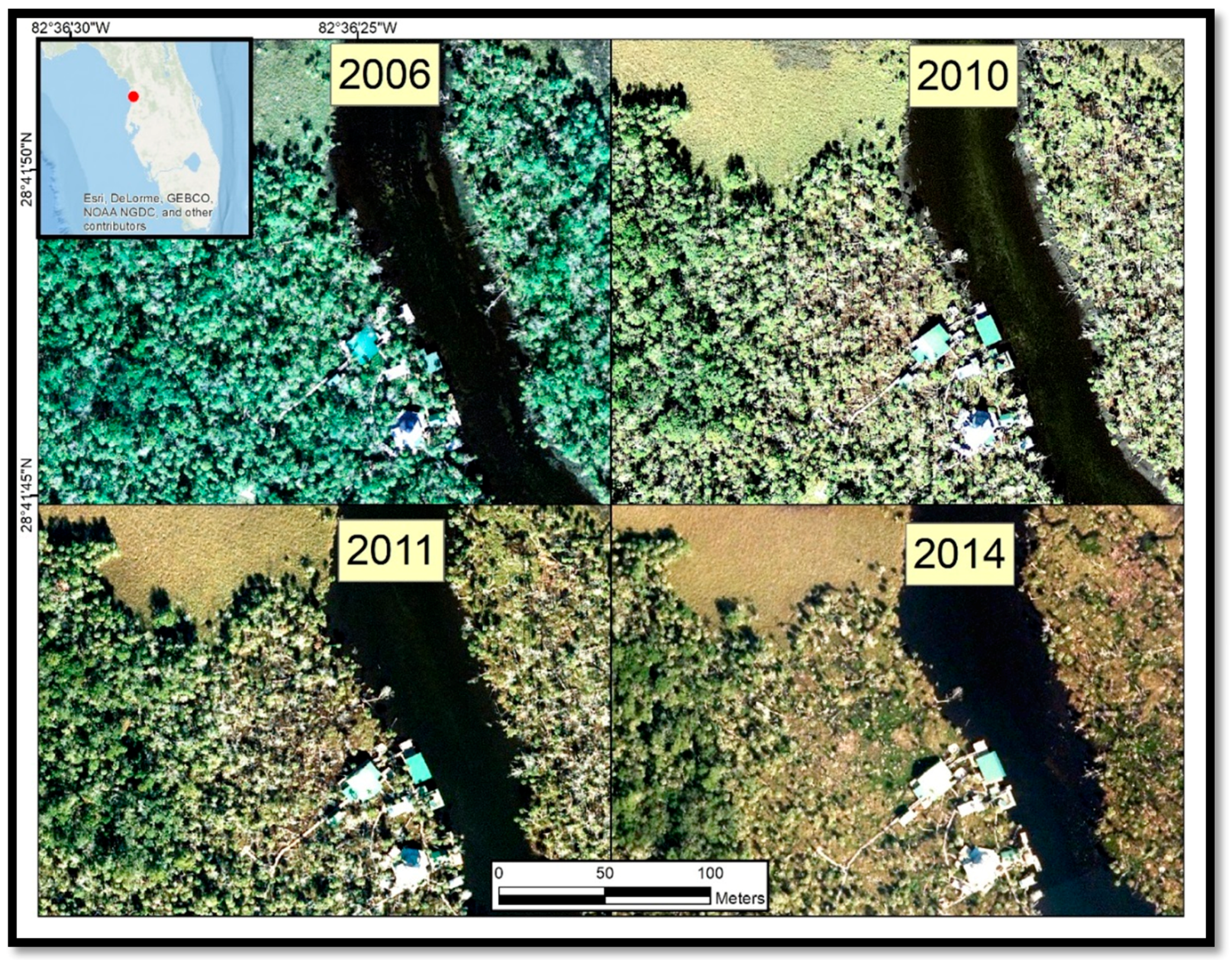
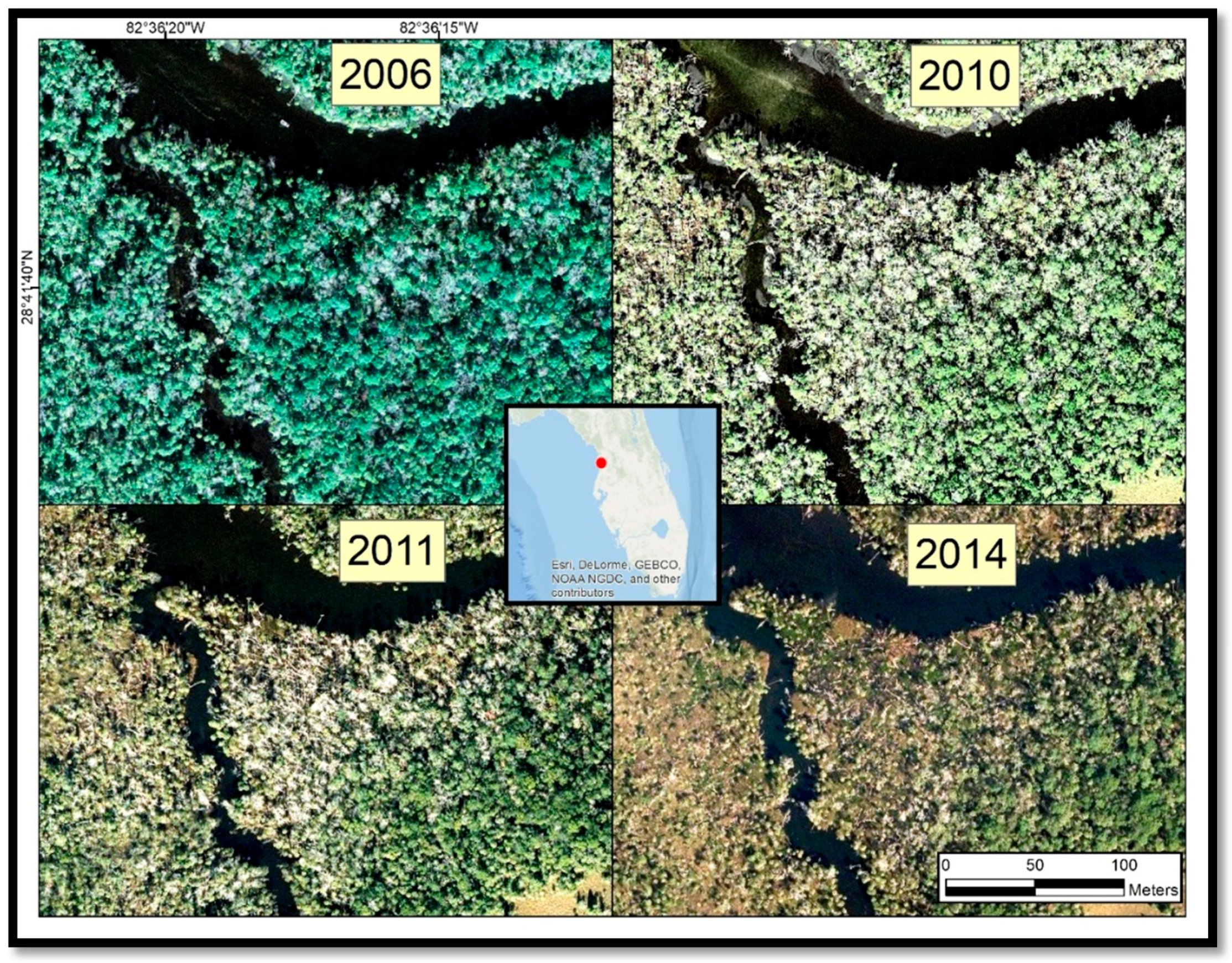
An increasing prevalence of dead palm trees on coastal islands within the study area has been attributed to sea-level rise [
13], but the geographic extent of the damage has not yet been quantified. Between 2009 and 2016, co-author Benjamin Dimmitt documented some of the degradation along waterways in and around CNWR using photographs collected from the ground (
Figure 2). The CNWR Comprehensive Management Plan [
13] has acknowledged that climate change impacts are expected to become more frequent and severe in the 2012–2027 timeframe, although the uncertainty of the severity and extent of the impacts is high. The synergistic effects of sea-level rise and increased storm surges will likely compound the threat to these coastal habitats.
2. Materials and Methods
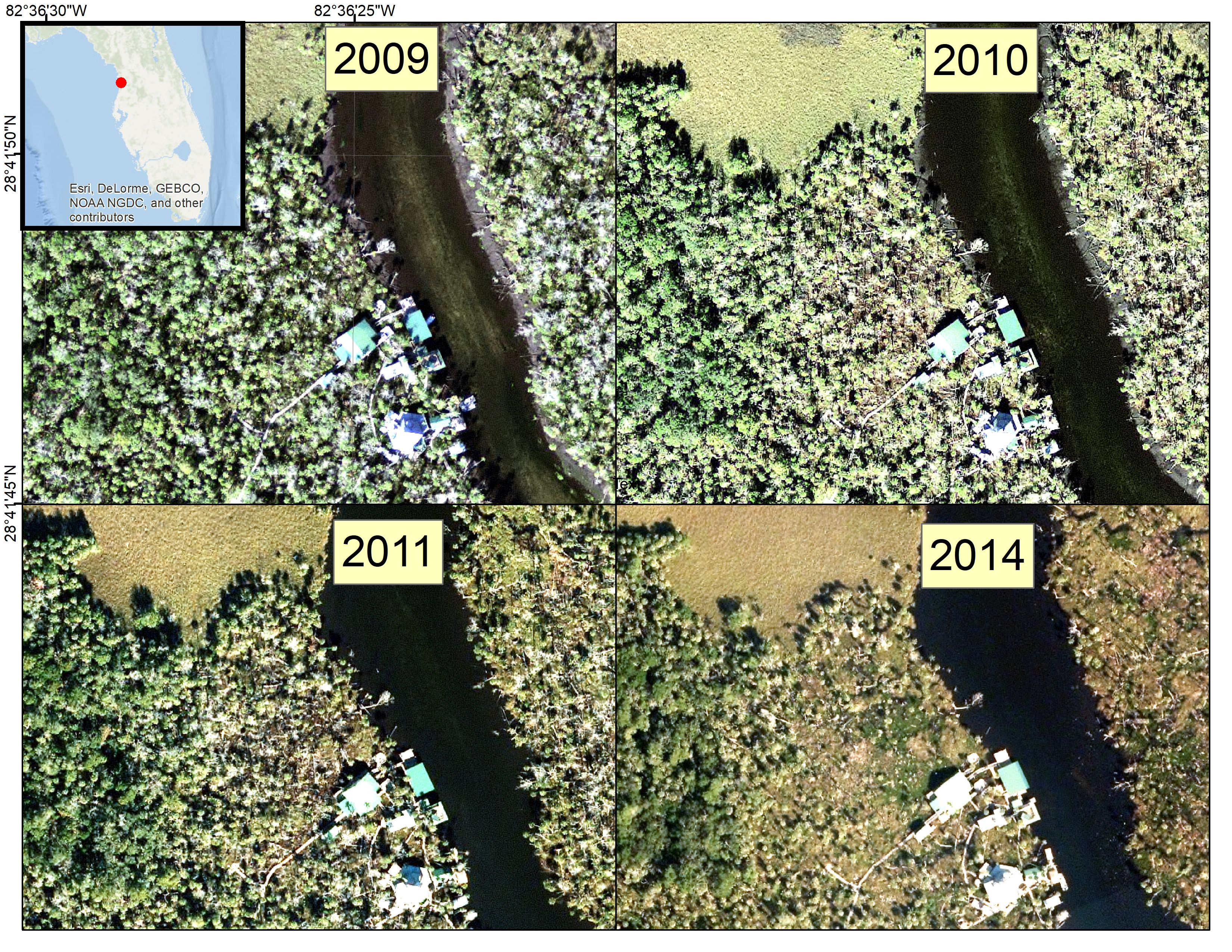
To visually assess the change in vegetation for the areas shown in Mr. Dimmitt’s photos, we examined aerial photographs for the years 2006–2011, and 2014 from the Florida Land Boundary Information Systems (
http://labins.org). To quantify the spatial change between 1982, 2003, and 2015 we obtained three Landsat satellite images (
Table 1; source:
http://EarthExplorer.usgs.gov). The multispectral Landsat data were used to derive normalized difference vegetation index (NDVI) maps of the study area at 30 m spatial resolution. NDVI is a robust method for evaluating vegetation health changes over time as measured by the spectral reflectance patterns of the vegetation, and results in values between −1 and 1 [
14].
To further examine the change of the habitat at scales relevant to individual trees, we derived similar NDVI maps for the study area using very-high-spatial-resolution (2 m) WorldView-2 satellite imagery for the years 2010 and 2017 (source: Digital Globe™).
Landsat images were radiometrically calibrated and converted to top-of-atmosphere reflectance fields using the ENVI™ software, prior to computing NDVI using the red and near-infrared bands. WorldView-2 images were preprocessed to reflectance and corrected for Rayleigh attenuation using a protocol developed by McCarthy et al. [
15], before computing NDVI using the red and near-infrared (band 8) bands. Because WorldView-2 imagery covers a much smaller area per image, multiple images were required to cover the study area. The individual NDVI maps were mosaicked together in ArcGIS v10.1. A coastal forest mask was defined using the criteria of the Cooperative Land Cover Map (v3.2.5) of Geselbracht et al. [
10] for this study area. The masks were used to select NDVI maps of our region of interest. For all Landsat (1982/2003, 2003/2015) and WorldView image pairs, the “before” NDVI maps were subtracted from the “after” maps to evaluate change. To account for noise or ephemeral changes, “vegetation decline” was defined as those pixels containing an NDVI difference value of less (more negative) than −0.05.
Langston et al. [
11] evaluated precipitation, temperature, river discharge, and storm data for their region as potential drivers of the degradation of stands of
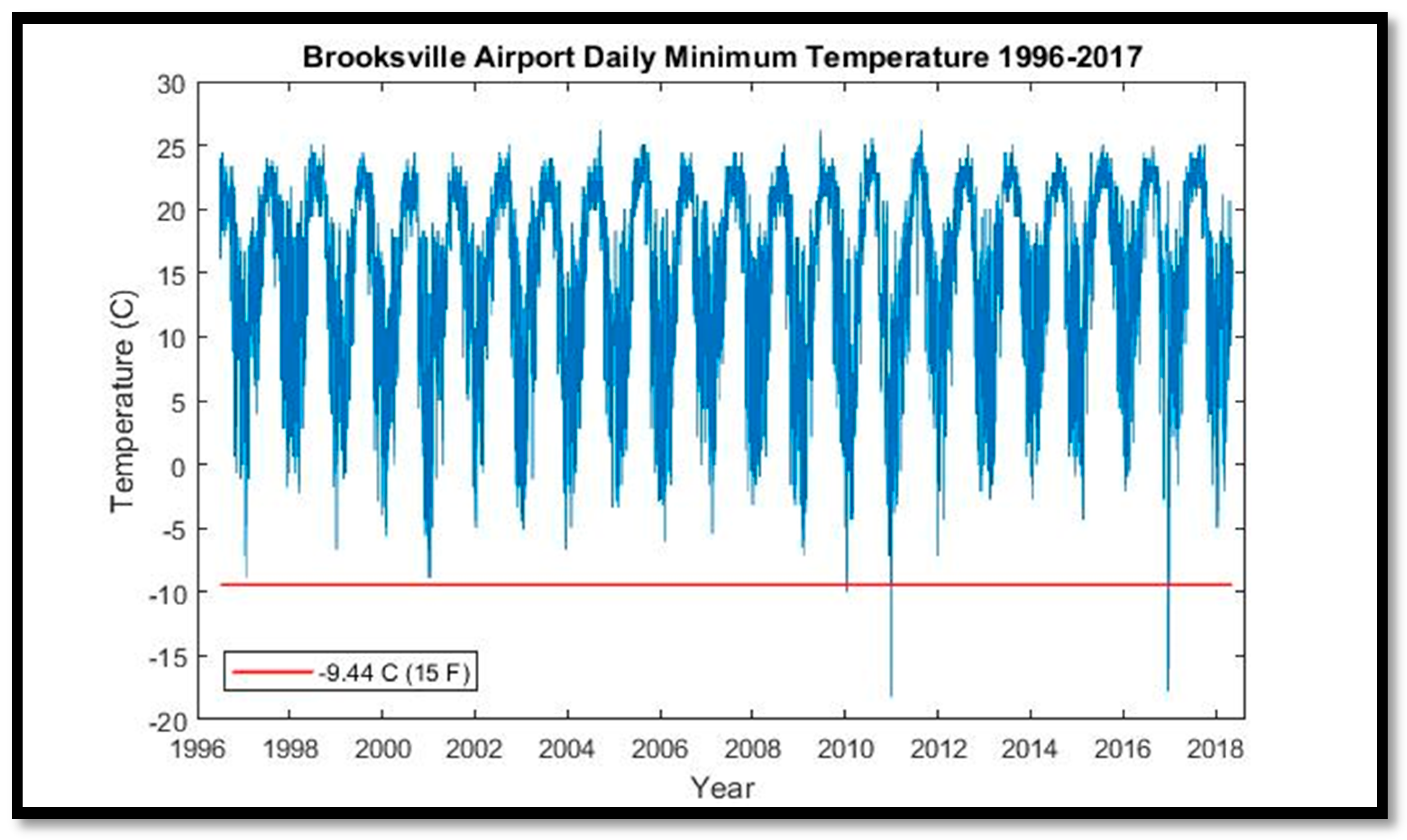
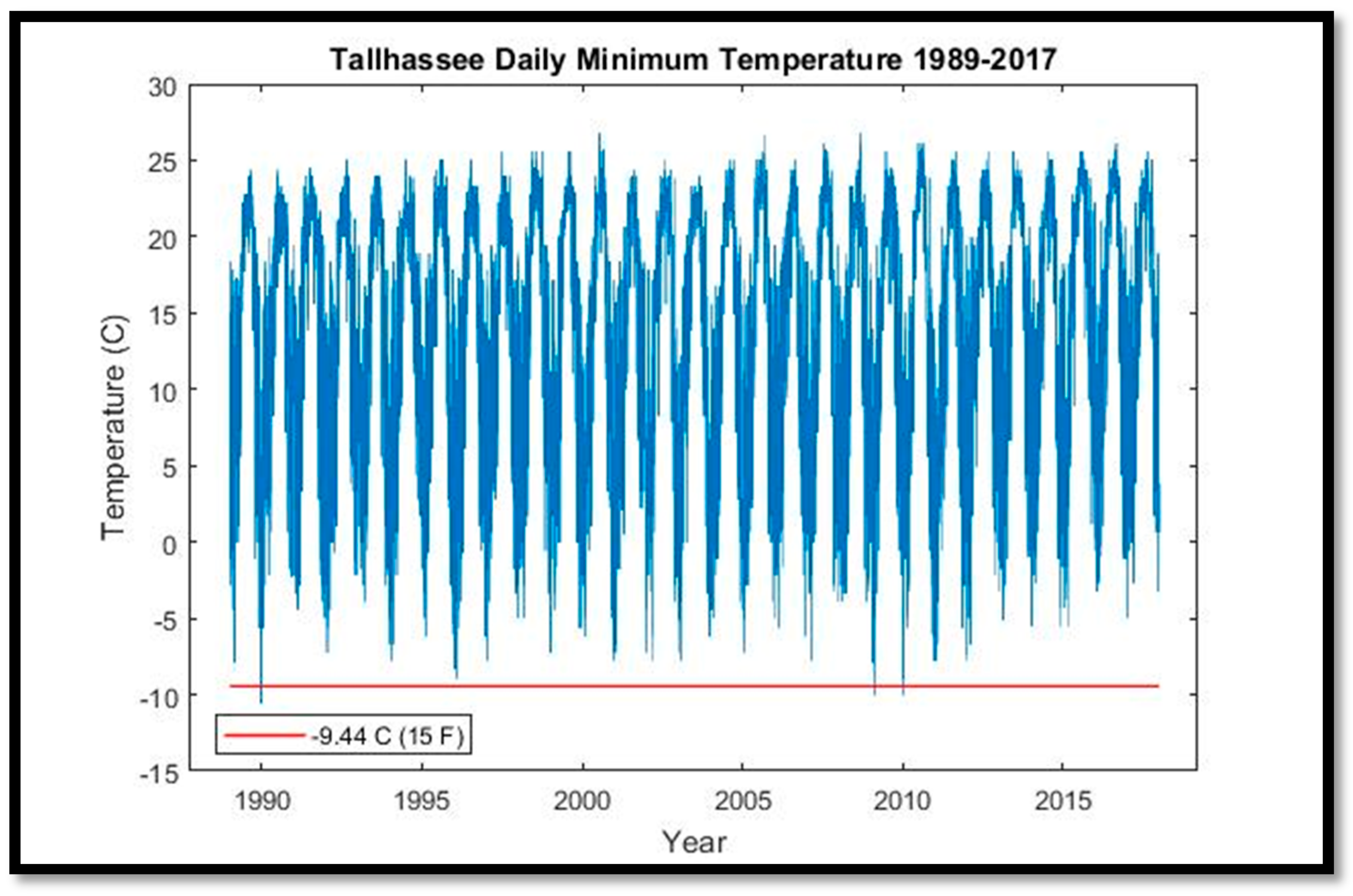
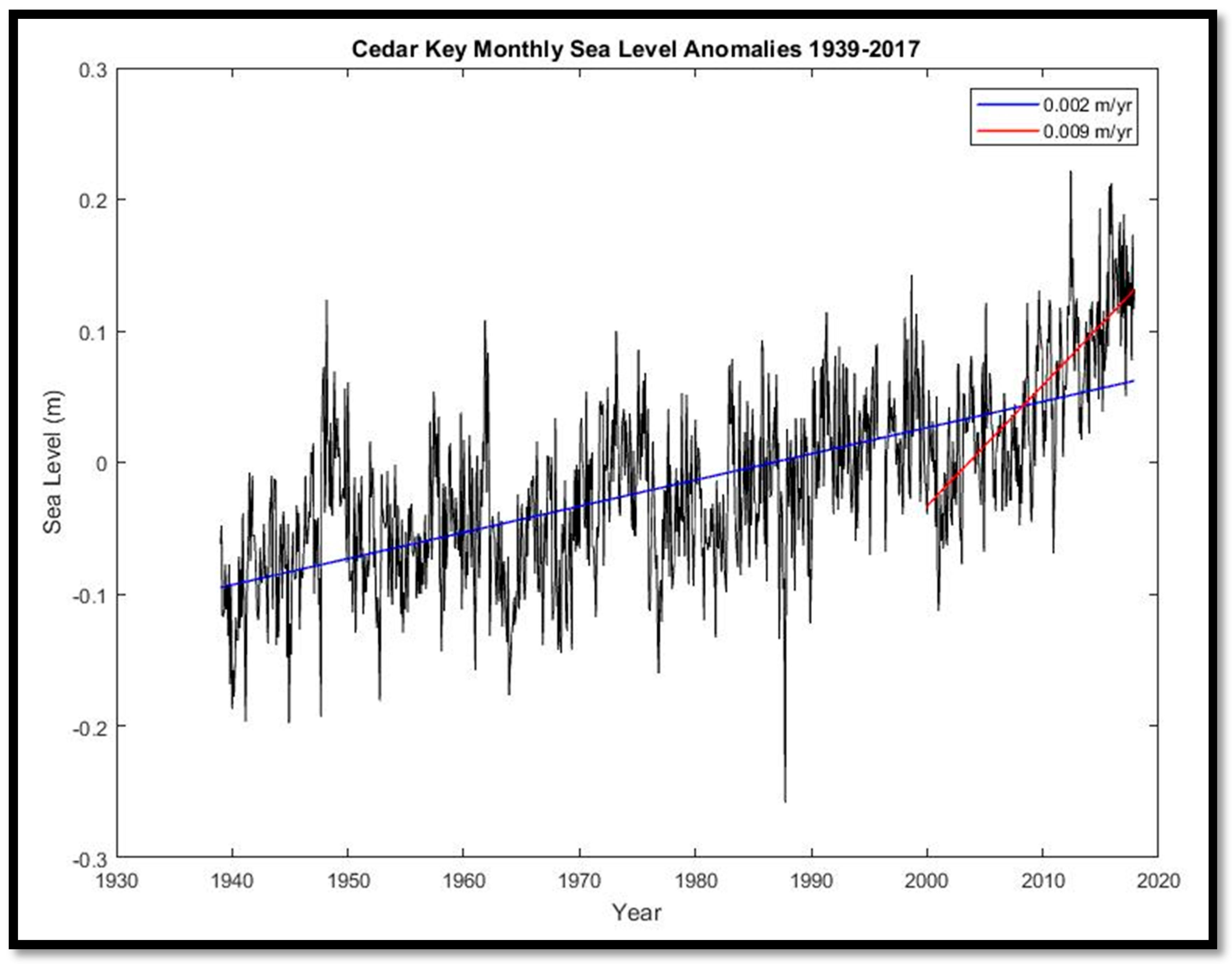
S. palmetto and other species. We attempted to replicate this approach by evaluating minimum and maximum daily temperatures from Brooksville, Florida (~35 km southeast of CNWR) for 1996–2017, and for Tallahassee, Florida (~250 km northwest of CNWR) for its longer time series (1942–2018). Precipitation data were downloaded from Inverness, Florida (~30 km northeast of CNWR). Monthly mean sea-level anomalies (de-seasonalized) were assessed from tide gauge #8727520 at Cedar Key, Florida (~62 km northwest of CNWR) for 1939–2017. River discharge, conductance, stream-level, and gauge-height data were downloaded for two United States Geologic Survey stations located within the Chassahowitzka River for the period 2007–2016.
4. Discussion
Coastal forests are increasingly vulnerable to extreme events in a changing global climate. Those along the coast in the Big Bend region of Florida are predicted to lose upwards of 50% by 2100 [
10,
17]. Burkett and Kusler [
18] summarize studies that specifically predict large-scale losses of Big Bend coastal marsh and
S. palmetto forests as a result of sea-level rise.
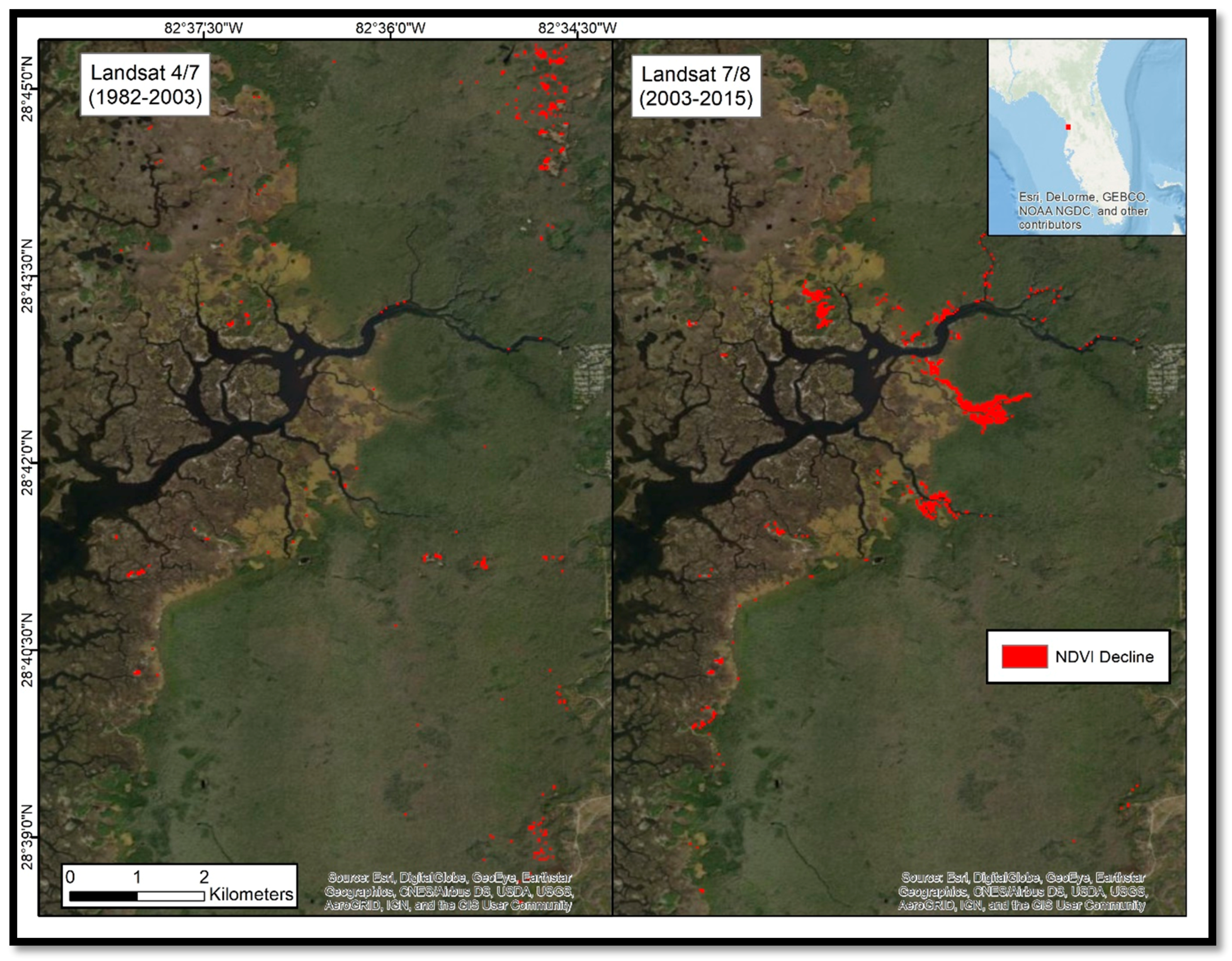
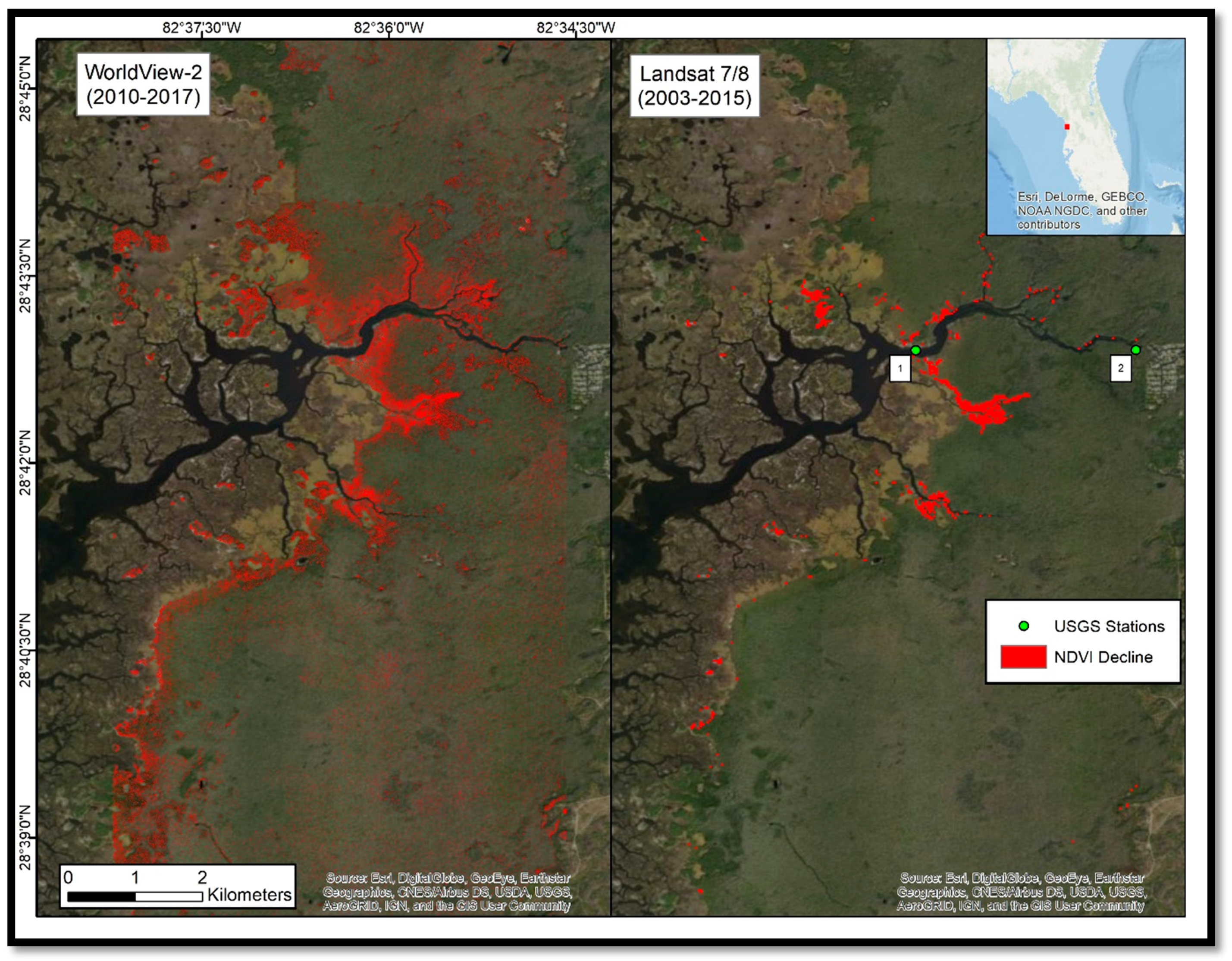
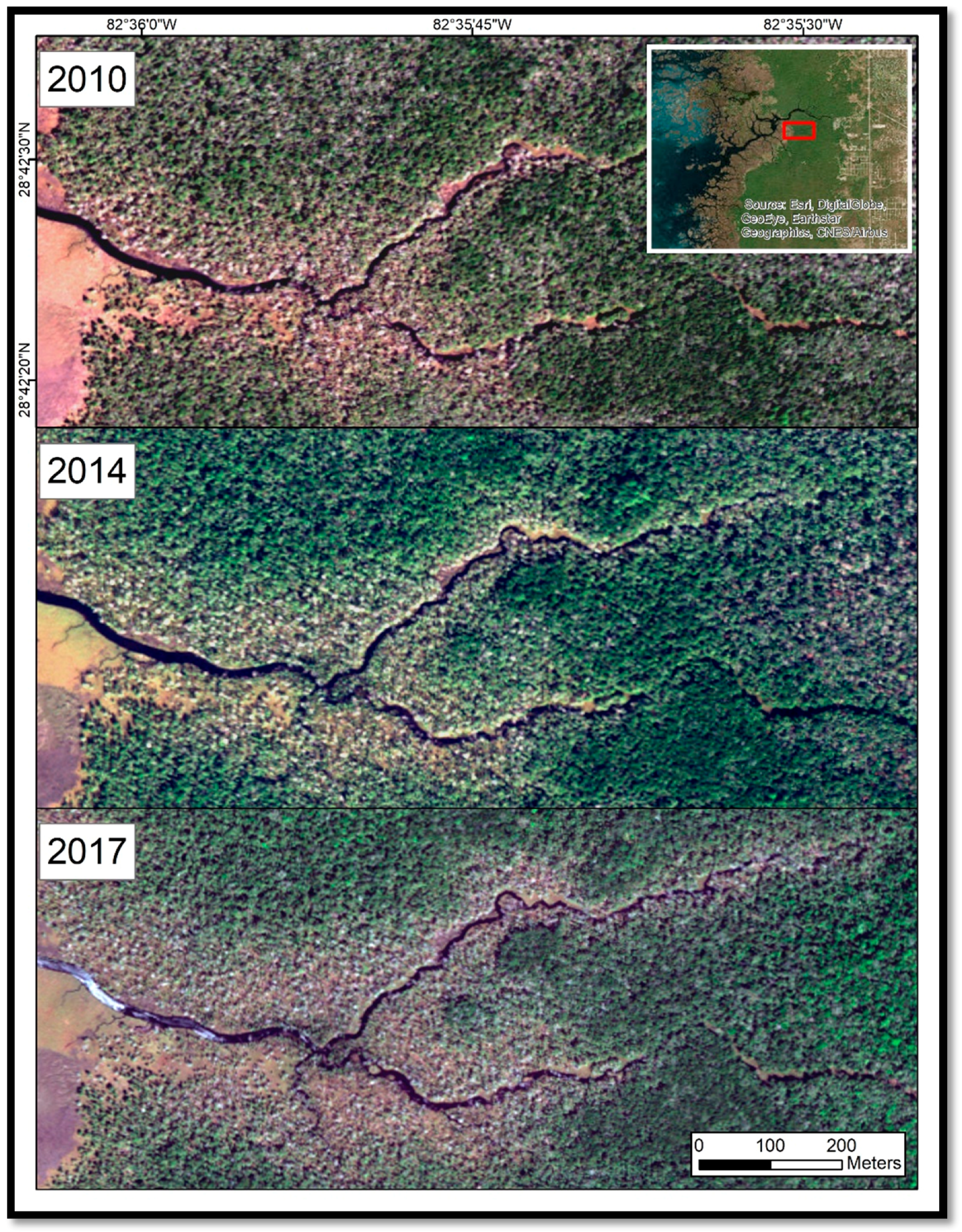
Our study found that coastal forest decline at the marsh–forest boundary and along waterways in the Big Bend region accelerated in the 2010–2017 timeframe. Sea-level records showed an acceleration in sea-level rise since 2000. Aerial photographs from 2006 to 2014 suggested that forest degradation did not noticeably occur until 2010. Aerial photographs and satellite imagery indicated that forest degradation continued through 2014 and 2017 (
Figure 10).
Time series of digital satellite-images showed only a small decline (0.36 km
2) from 1982 to 2003, and a decline of 0.60 km
2 from 2003 to 2015 with 30 m Landsat imagery, but a loss of 4.37 km
2 from 2010 to 2017 with 2 m WorldView-2 imagery. The discrepancy of the observed decline in the latter two imagery pairs may be explained by the additional two years of study and the greater spatial resolution of the WorldView-2 imagery. The coarser Landsat data may contain mixed signals within each 900 m
2 pixel that are better resolved by the 4 m
2 WorldView-2 data. Discrepancies in preprocessing steps (i.e., top-of-atmosphere Landsat reflectance vs. atmospherically corrected WorldView-2 reflectance) may also have contributed to the discrepancies. However, Zhang et al. [
19] found only marginal differences in agreement (r
2 > 0.89 vs. r
2 > 0.87, respectively) between NDVI results, from multi-sensor analyses, from when imagery was atmospherically corrected compared to when only converted to top-of-atmosphere reflectance. Further, the use of −0.05 as a threshold for signal vs. noise was chosen arbitrarily as no standard could be identified in the literature. Nonetheless, the patterns of decline location and extent agreed (
Figure 6). The visual agreement of apparent degradation, as observed by the four independent data sources (i.e., in-situ photographs, imagery from WorldView-2, Landsat, and aerial surveys), was used to identify −0.05 as an appropriate threshold. Given that appropriate ground-truth data does not exist for this region during the time period studied, we relied on the in-situ photographs and agreement between independent data sources to substantiate the observed patterns of loss.
Langston et al. [
11] studied mortality and recovery of 14 coastal tree species relative to tidal flooding, at a site located approximately 47 km northwest of this study area, during field campaigns in 1992–1998, 2000, 2005, and 2014–2016. Flooding rates were observed in 1992 and 1993, and modeled thereafter. The species-specific data shows that
Quercus laurafolia, Morus rubra, Pinus taeda, and others were lost or found relict at all sites in 2014, regardless of whether flooding occurred or not. The USDA describes these three species as having no tolerance to salinity, but indicates that they are relatively tolerant to cold air temperatures (−19 to −36 °C;
plants.usda.gov). Given that the coldest temperature recorded at Brooksville Airport was −18.2 °C, the loss of these species may be attributable to saltwater-intrusion-induced salt stress. However,
S. palmetto, which is characterized as having “medium” salinity tolerance, but a minimum temperature tolerance of only −10 °C (
plants.usda.gov), saw new growth in all five of the plots with minimal flooding as of 2014, while the flooded
S. palmetto plots contained only lost or relict stands. Their data, however, were unable to resolve the observed 2010 event, or determine when die-off occurred. Further, they reported that
Q. virginiana was regenerating in one plot in 2005, but relict there by 2014, relict in two plots, and regenerating in two plots, all of which reportedly experienced zero or one flooding week during this time. The USDA reports that
Q. virginiana has a “low” salinity tolerance, and minimum temperature tolerance of −13.8 °C (
plants.usda.gov). The Brooksville Airport’s lowest temperature of −18.2 °C occurred in January of 2011, which may help explain the mixed response of this species that has some salinity tolerance and did not experience flooding. Langston et al. [
11] did evaluate temperature as a potential driver of decline, but only used annual temperatures, and therefore could not resolve the local single-day record-low temperature events in 2010 and 2011.
The cold snaps beginning in 2009 that exceeded the S. palmetto tolerance threshold of −10 °C were the first in 20 years, which itself set a record for the longest such hiatus over the period of record (1940–2017). The advent of high-spatial-resolution aerial and satellite imagery, combined with interannual or better repeat imagery acquisition, have allowed us to document the timing and extent of coastal forest decline, and correlate it with concomitant environmental phenomena. Further research, including the collection of robust ground validation data, is needed to monitor and assess future patterns of loss.
While Langston et al. [
11] report regeneration of some species in some low-flooding plots in 2014,
Figure 10 shows continued decline in 2014 and 2017 for our nearby study area. Doyle [
20] predicted an estimated loss of 13,000 ha of Big Bend coastal forest under a one meter sea-level rise scenario by 2100, representing a rate of approximately 0.25% loss per year of the area studied here. Geselbracht et al. [
10] predicted a loss of 0.5% per year. Our estimates suggest the loss from 2010 to 2017 is approximately 1% per year.
5. Conclusions
There is accelerated die-off of coastal forest along the Big Bend of Florida. The decline is documented by in-situ photographs, and in synoptic aerial and satellite imagery, although formal validation with ground-truth surveys was not feasible. Time series of the latter show that the decline accelerated in the last decade with no apparent regeneration.
We conclude that a combination of chronic sea-level rise stress and acute cold snap events have caused rapid, sustained, and likely irreversible damage to coastal forest along the largest stretch of undeveloped coastal land in the continental United States. Further decline is expected with an accelerating sea-level rise and an increase in extreme weather events.
High-resolution satellite images should be used to monitor seasonal to annual changes for the Big Bend region. This area serves as a canary in the coalmine for the compounded impacts on coastal habitats of sea-level rise and extremes in meteorological events.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}