Toward Mapping Dietary Fibers in Northern Ecosystems Using Hyperspectral and Multispectral Data


Abstract
:1. Introduction
2. Materials and Methods
2.1. Greenhouse Procedures
2.2. Hyperspectral, Leaf Area, and Destructive Vegetation Collection
2.3. Statistical Analyses
2.4. Band Equivalent Reflectance
3. Results
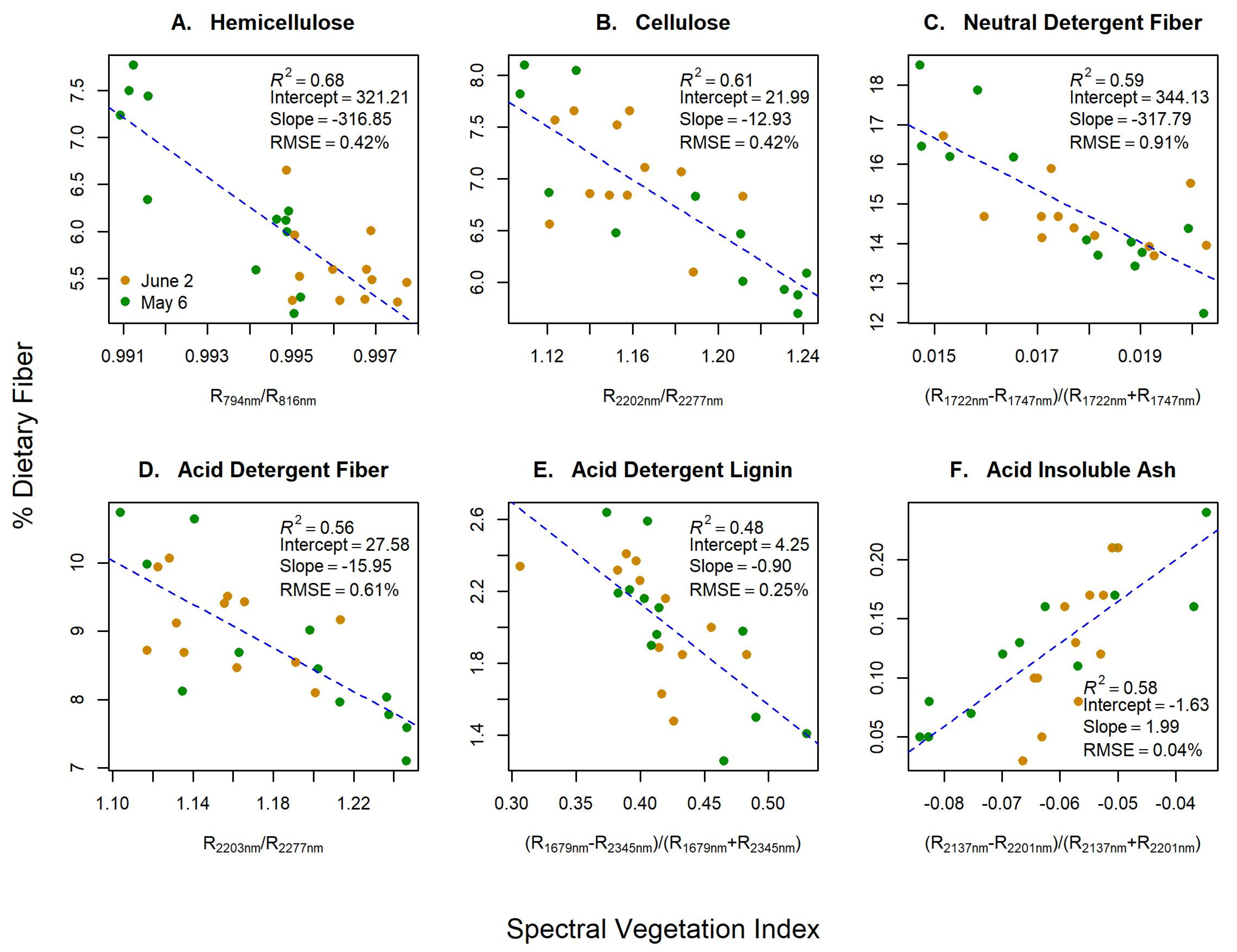
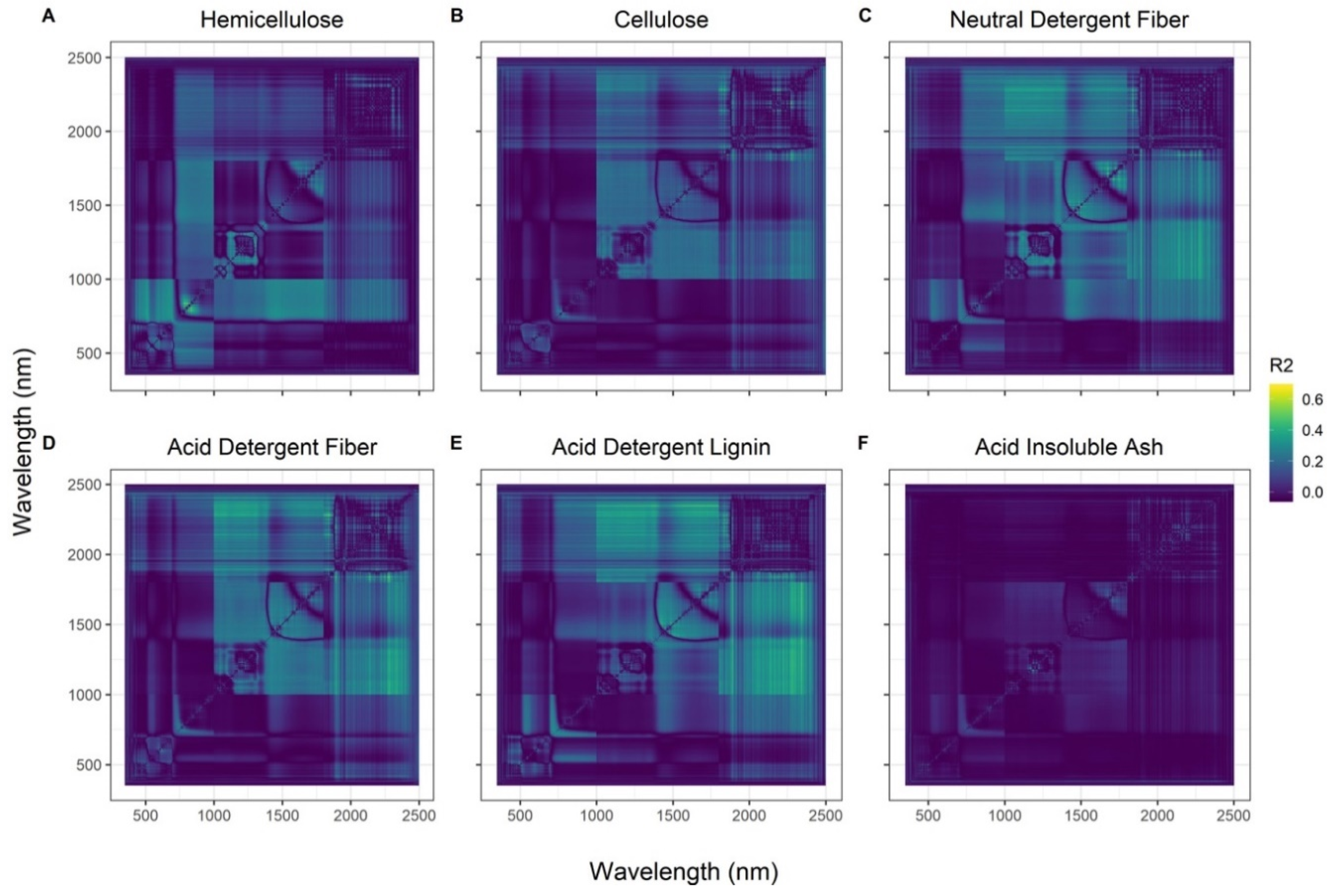
3.1. Hyperspectral Vegetation Indices
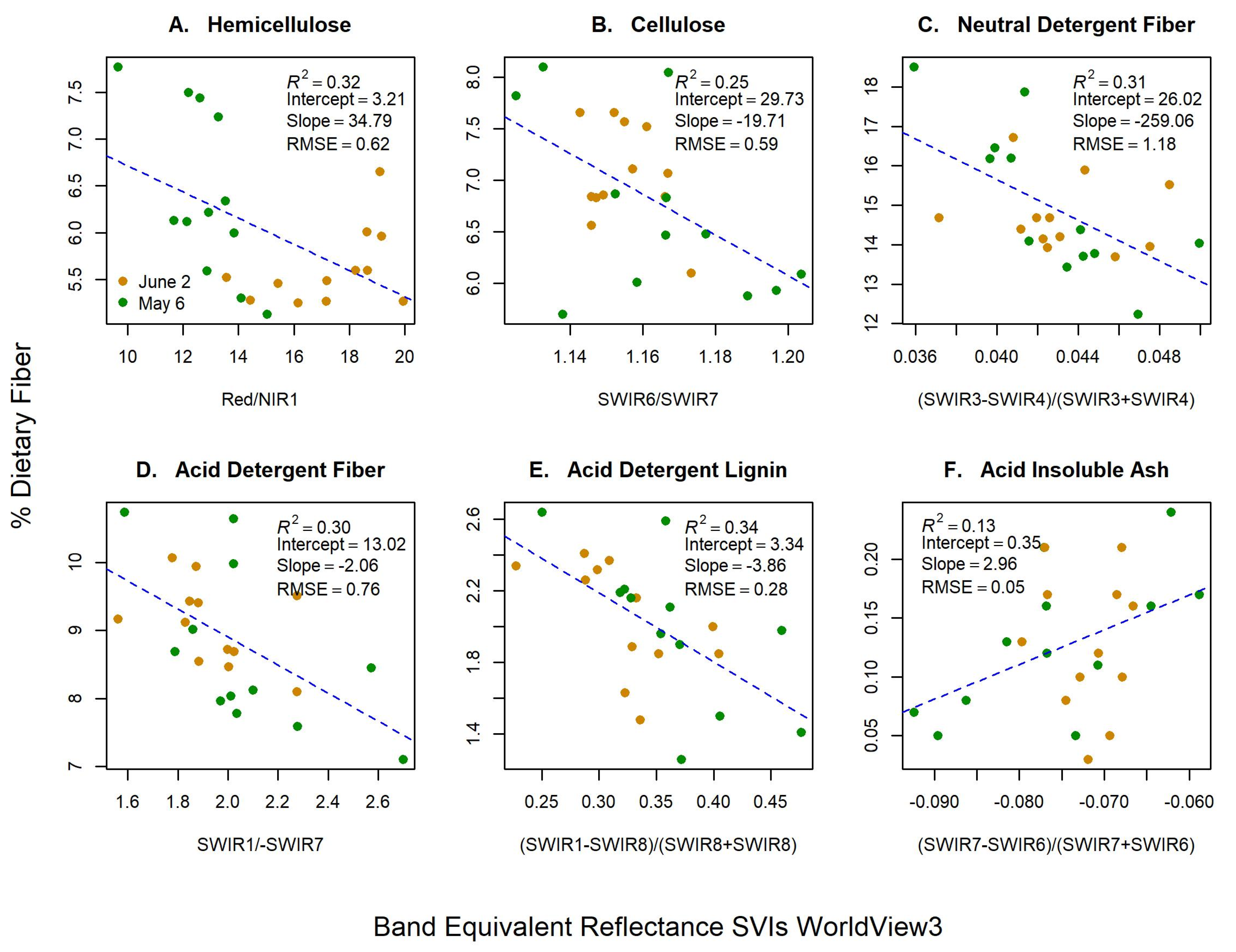
3.2. Band Equivalent Reflectance of WV-3
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Details on Nitrogen (N) Fertilizer Treatment Estimation
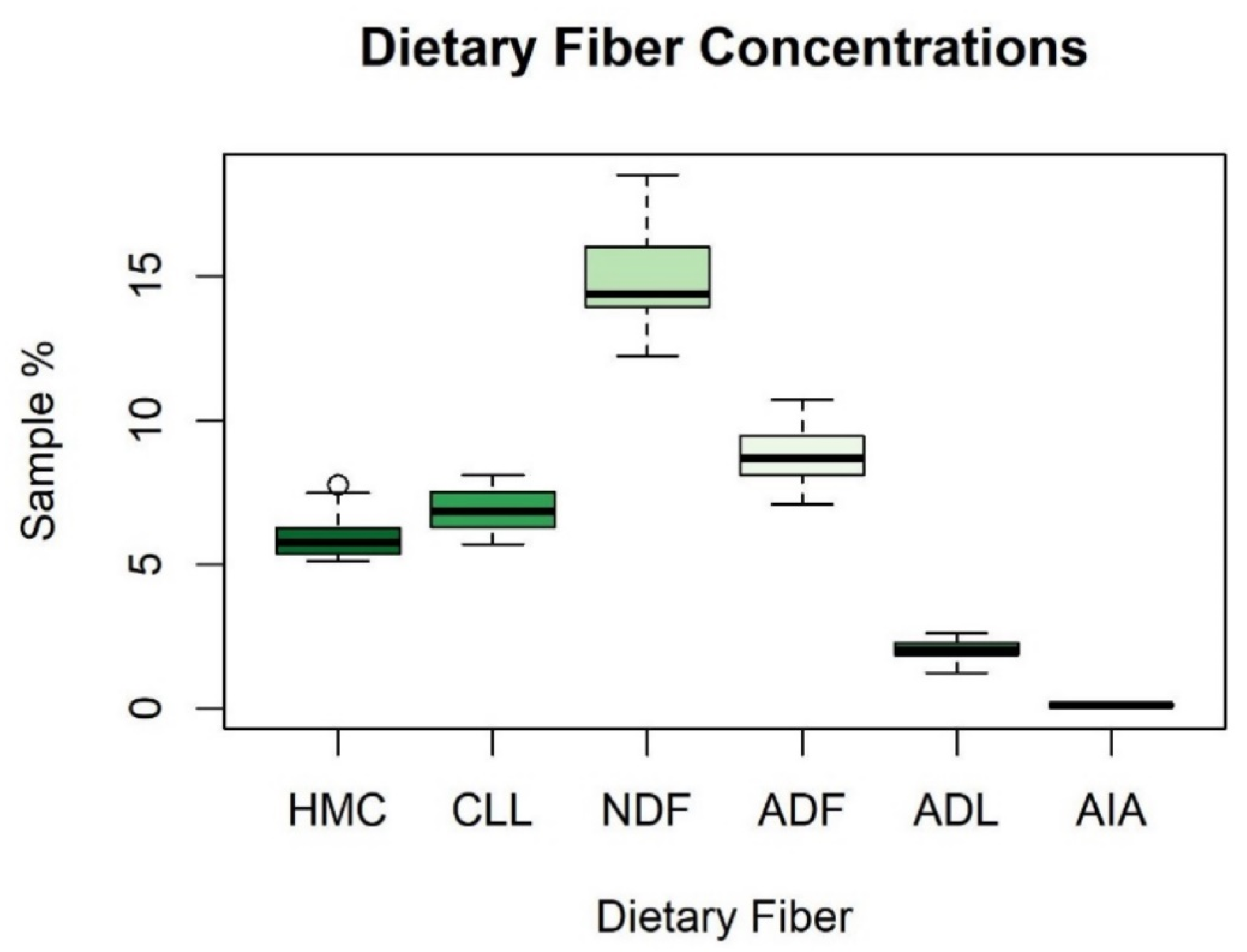
Appendix B. Summary Statistics for Dietary Fibers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fiber | Range | Mean | Standard Deviation |
|---|---|---|---|
| HMC | 5.13–7.77% | 6.01% | 0.79% |
| CLL | 5.70–8.10% | 6.87% | 0.71% |
| NDF | 12.24–18.51% | 14.89% | 1.48% |
| ADF | 7.11–10.74% | 8.89% | 0.95% |
| ADL | 1.26–2.64% | 2.02% | 0.37% |
| AIA | 0.03–0.24% | 0.12% | 0.06% |

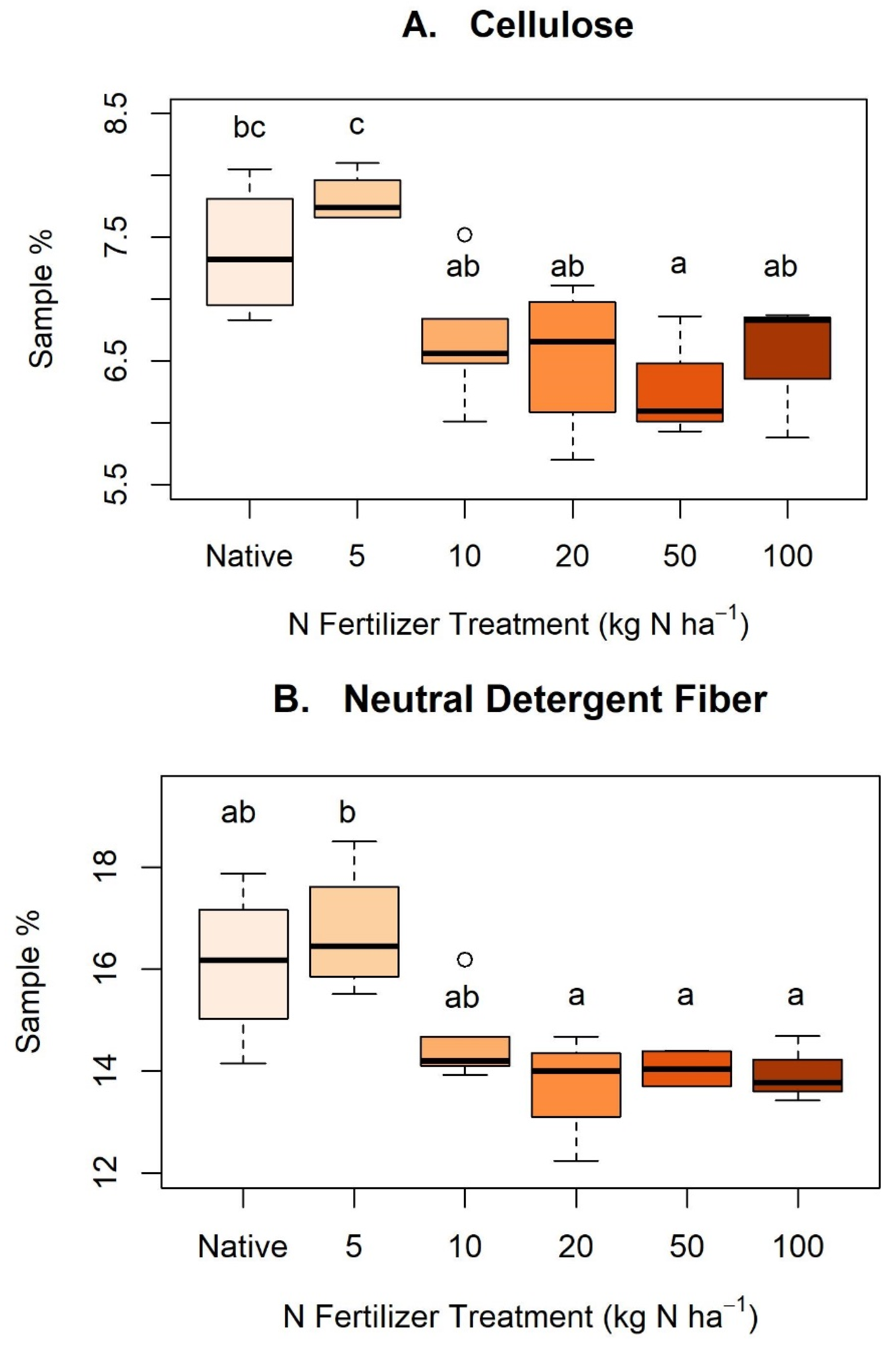
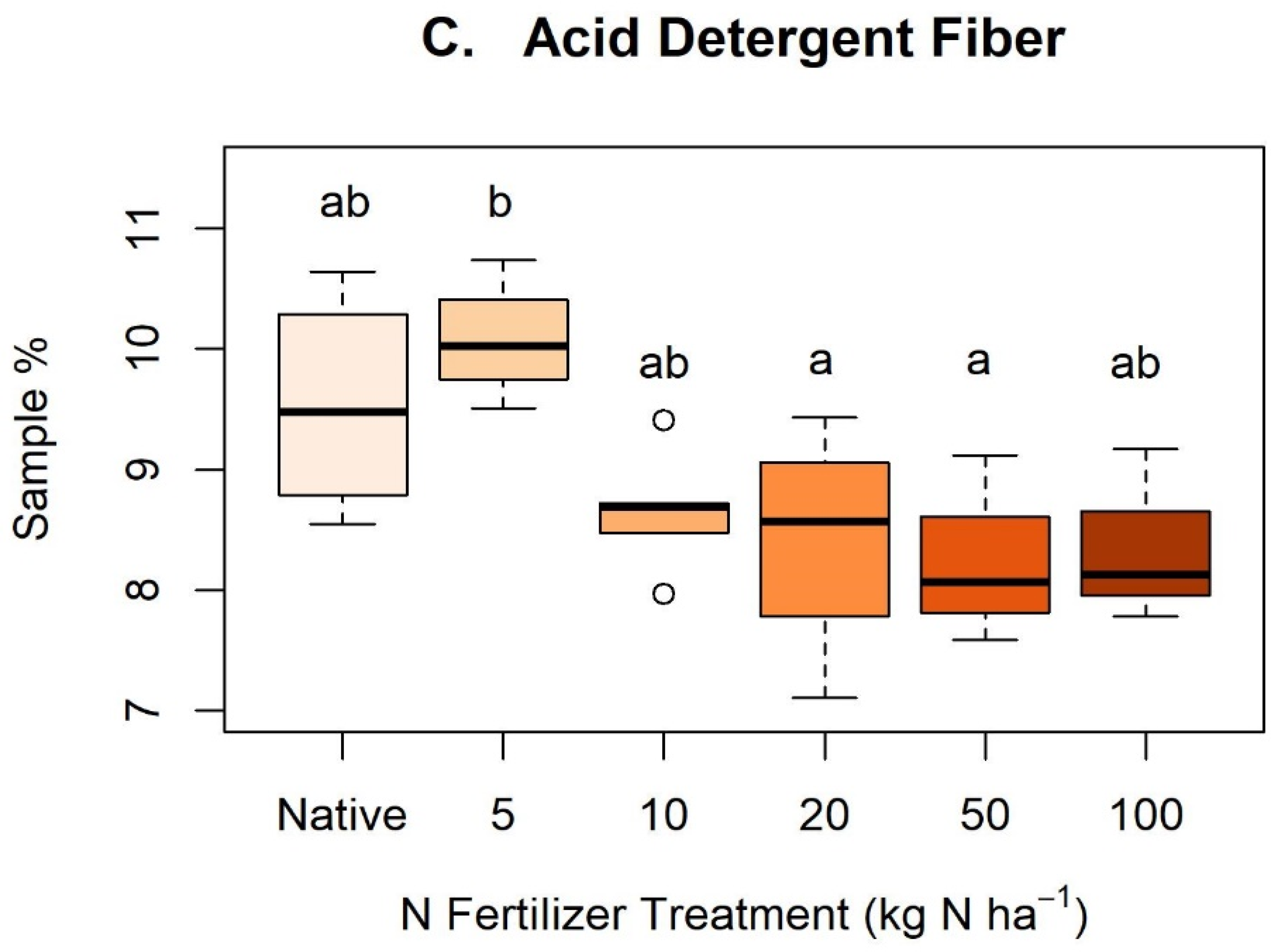
Appendix C. Nitrogen Treatments and Cellulose, Neutral Detergent Fiber, and Acid Detergent Fiber


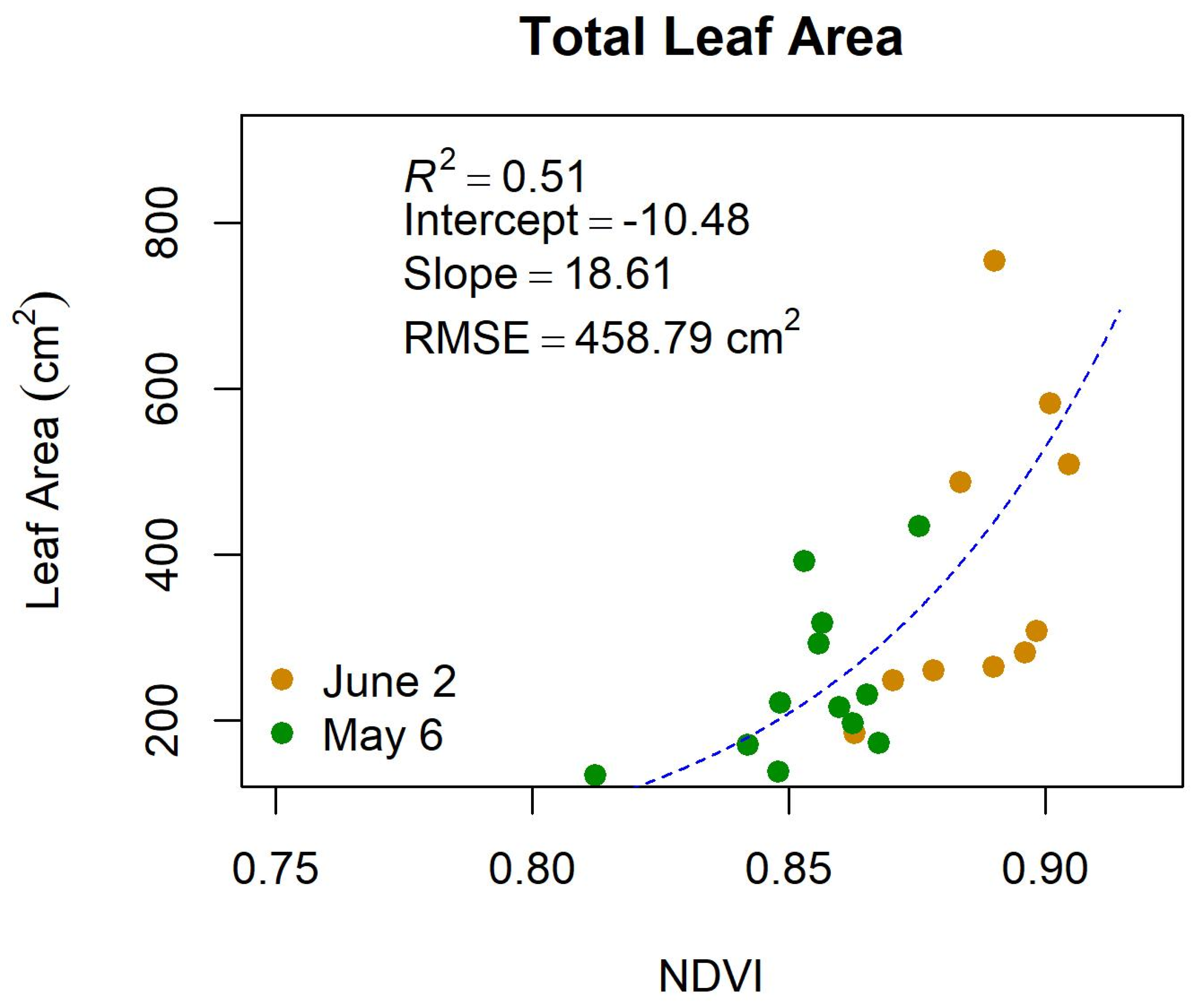
Appendix D. Results of Swapping Leaf Area for the Normalized Difference Vegetation Index
| Models | R2 | RMSE | AICc | ΔAICc | LOOCV Slope | LOOCV | ΔLOOCV |
|---|---|---|---|---|---|---|---|
| Hemicellulose (HMC) | |||||||
| SVI | 0.32 | 0.62% | 52.65 | - | 0.27 | 0.52 | - |
| SVI+ NDVI | 0.37 | 4.28% | 52.41 | −0.24 | 0.67 | 0.53 | +1% |
| Cellulose (CLL) | |||||||
| SVI | 0.25 | 0.59% | 50.10 | - | 0.22 | 0.45 | - |
| SVI+ NDVI | 0.21 | 0.59% | 53.00 | +2.90 | 0.14 | 0.47 | +5% |
| Neutral detergent fiber (NDF) | |||||||
| SVI | 0.31 | 1.18% | 83.33 | - | 0.26 | 0.67 | - |
| SVI+ NDVI | 0.28 | 1.18% | 86.09 | +2.76 | 0.22 | 0.62 | −5% |
| Acid detergent fiber (ADF) | |||||||
| SVI | 0.30 | 0.76% | 62.10 | - | 0.28 | 0.57 | - |
| SVI+ NDVI | 0.27 | 0.76% | 64.92 | +2.82 | 0.21 | 0.58 | +9% |
| Acid detergent lignin (ADL) | |||||||
| SVI | 0.34 | 0.28% | 15.05 | - | 0.33 | 0.70 | - |
| SVI+ NDVI | 0.34 | 0.28% | 16.99 | +1.94 | 0.33 | 0.71 | +1% |
| Acid detergent ash (AIA) | |||||||
| SVI | 0.13 | 0.05% | −66.35 | - | 0.11 | 0.31 | - |
| SVI+ NDVI | 0.13 | 0.05% | −64.44 | +1.91 | 0.07 | 0.33 | +2% |

References
- Serreze, M.C.; Walsh, J.E.; Chapin, F.S.I.; Osterkamp, T.; Dyurgerov, M.; Romanovsky, V.; Oechel, W.C.; Morison, J.; Zhang, T.; Barry, R.G. Observational evidence of recent change in the northern high- latitude environment. Clim. Chang. 2000, 46, 159–207. [Google Scholar] [CrossRef]
- Wolken, J.M.; Hollingsworth, T.N.; Rupp, T.S.; Chapin, F.S.; Trainor, S.F.; Barrett, T.M.; Sullivan, P.F.; McGuire, A.D.; Euskirchen, E.S.; Hennon, P.E.; et al. Evidence and implications of recent and projected climate change in Alaska’s forest ecosystems. Ecosphere 2011, 2, 1–35. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Forbes, B.C.; Wilmking, M.; Hallinger, M.; Lantz, T.; Blok, D.; Tape, K.D.; Macias-fauria, M.; Sass-klaassen, U.; Esther, L. Shrub expansion in tundra ecosystems: Dynamics, impacts and research priorities. Environ. Res. Lett. 2011, 6, 045509. [Google Scholar] [CrossRef] [Green Version]
- Sturm, M.; Racine, C.; Tape, K.; Cronin, T.W.; Caldwell, R.L.; Marshall, J. Increasing shrub abundance in the Arctic. Nature 2001, 411, 546–547. [Google Scholar] [CrossRef]
- Tape, K.D.; Gustine, D.D.; Ruess, R.W.; Adams, L.G.; Clark, J.A. Range expansion of moose in Arctic Alaska linked to warming and increased shrub habitat. PLoS ONE 2016, 11, e0152636. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Tape, K.D.; Prugh, L.; Kofinas, G.; Carroll, G.; Kielland, K. Enhanced shrub growth in the Arctic increases habitat connectivity for browsing herbivores. Glob. Chang. Biol. 2020, 26, 3809–3820. [Google Scholar] [CrossRef]
- Bryant, J.P. Feltleaf willow-snowshoe hare interactions: Plant carbon/nutrient balance and floodplain succession. Ecology 1987, 68, 1319–1327. [Google Scholar] [CrossRef]
- Christie, K.S.; Bryant, J.P.; Gough, L.; Ravolainen, V.T.; Ruess, R.W.; Tape, K.D. The role of vertebrate herbivores in regulating shrub expansion in the Arctic: A synthesis. Bioscience 2015, 65, 1123–1133. [Google Scholar] [CrossRef] [Green Version]
- Butler, L.G.; Kielland, K. Acceleration of vegetation turnover and element cycling by mammalian herbivory in riparian ecosystems. J. Ecol. 2008, 96, 136–144. [Google Scholar] [CrossRef]
- Kielland, K.; Bryant, J.P. Moose herbivory in Taiga: Effects on biogeochemistry and vegetation dynamics in primary succession. OIKOS 1998, 82, 377–383. [Google Scholar] [CrossRef]
- Speed, J.D.M.; Austrheim, G.; Hester, A.J.; Mysterud, A.; Speed, J.D.M.; Austrheim, G.; Hester, A.J.; Mysterud, A. Experimental evidence for herbivore limitation of the treeline. Ecology 2010, 91, 3414–3420. [Google Scholar] [CrossRef] [PubMed]
- Doiron, M.; Gauthier, G.; Lévesque, E. Effects of experimental warming on nitrogen concentration and biomass of forage plants for an arctic herbivore. J. Ecol. 2014, 102, 508–517. [Google Scholar] [CrossRef]
- Zamin, T.J.; Côté, S.D.; Tremblay, J.P.; Grogan, P. Experimental warming alters migratory caribou forage quality. Ecol. Appl. 2017, 27, 2061–2073. [Google Scholar] [CrossRef] [PubMed]
- Felton, A.M.; Wam, H.K.; Stolter, C.; Mathisen, K.M.; Wallgren, M. The complexity of interacting nutritional drivers behind food selection, a review of northern cervids. Ecosphere 2018, 9, e02230. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and non- starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Shipley, L.A.; Spalinger, D.E. Mechanics of browsing in dense food patches: Effects of plant and animal morphology on intake rate. Can. J. Zool. 1992, 70, 1743–1752. [Google Scholar] [CrossRef]
- Barboza, P.S.; Parker, K.L.; Hume, I.D. Integrative Wildlife Nutrition; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Xue, J.; Su, B. Significant remote sensing vegetation indices: A review of developments and applications. J. Sens. 2017. [Google Scholar] [CrossRef] [Green Version]
- Rouse, J.W., Jr.; Haas, R.H.; Schell, J.A.; Deering, D.W. Paper A 20. In Third Earth Resources Technology Satellite-1 Symposium: The Proceedings of A SYMPOSIUM Held by Goddard Space Flight Center at Washington, DC on 10–14 December 1973: Prepared at Goddard Space Flight Center; Scientific and Technical Information Office, National Aeronautics and Space Administration: Washington, DC, USA, 1974; Volume 351, p. 309. [Google Scholar]
- Doiron, M.; Legagneux, P.; Gauthier, G.; Lévesque, E. Broad-scale satellite Normalized Difference Vegetation Index data predict plant biomass and peak date of nitrogen concentration in Arctic tundra vegetation. Appl. Veg. Sci. 2013, 16, 343–351. [Google Scholar] [CrossRef]
- Johnson, H.E.; Gustine, D.D.; Golden, T.S.; Adams, L.G.; Parrett, L.S.; Lenart, E.A.; Barboza, P.S. NDVI exhibits mixed success in predicting spatiotemporal variation in caribou summer forage quality and quantity. Ecosphere 2018, 9, e02461. [Google Scholar] [CrossRef] [Green Version]
- Kokaly, R.F.; Asner, G.P.; Ollinger, S.V.; Martin, M.E.; Wessman, C.A. Characterizing canopy biochemistry from imaging spectroscopy and its application to ecosystem studies. Remote Sens. Environ. 2009, 113, 78–91. [Google Scholar] [CrossRef]
- Elvidge, C.D. Visible and near-infrared reflectance characteristics of dry plant materials. Int. J. Remote Sens. 1990, 11, 1775–1795. [Google Scholar] [CrossRef]
- Curran, P.J. Remote sensing of foliar chemistry. Remote Sens. Environ. 1989, 30, 271–278. [Google Scholar] [CrossRef]
- Wang, Z.; Townsend, P.A.; Schweiger, A.K.; Couture, J.J.; Singh, A.; Hobbie, S.E.; Cavender-bares, J. Mapping foliar functional traits and their uncertainties across three years in a grassland experiment. Remote Sens. Environ. 2019, 221, 405–416. [Google Scholar] [CrossRef]
- Knox, N.M.; Skidmore, A.K.; Prins, H.H.; Heitkönig, I.M.; Slotow, R.; van der Waal, C.; de Boer, W.F. Remote sensing of forage nutrients: Combining ecological and spectral absorption feature data. Isprs J. Photogramm. Remote Sens. 2012, 72, 27–35. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Carranza-jim, L.; Sinca, F.; Tupayachi, R.; Anderson, C.B.; Martinez, P. Functional and biological diversity of foliar spectra in tree canopies throughout the Andes to Amazon region. New Phytol. 2014, 204, 127–139. [Google Scholar] [CrossRef]
- Mirik, M.; Norland, J.E.; Crabtree, R.L.; Biondini, M.E. Hyperspectral one-meter-resolution remote sensing in Yellowstone National Park, Wyoming: I. Forage Nutritional Values. Soc. Range Manag. 2005, 58, 452–458. [Google Scholar] [CrossRef]
- Starks, P.J.; Coleman, S.W.; Phillips, W.A. Determination of forage chemical composition using remote sensing. J. Range Manag. 2004, 57, 635–640. [Google Scholar] [CrossRef]
- Chen, J.M.; Cihlar, J. Retrieving leaf area index of boreal conifer forests using landsat TM images. Remote Sens. Environ. 1996, 55, 153–162. [Google Scholar] [CrossRef]
- Turner, D.P.; Cohen, W.B.; Kennedy, R.E.; Fassnacht, K.S.; Briggs, J.M. Relationships between leaf area index and Landsat TM spectral vegetation indices across three temperate zone sites. Remote Sens. Environ. 1999, 70, 52–68. [Google Scholar] [CrossRef]
- Asner, G.P. Biophysical and biochemical sources of variability in canopy reflectance. Remote Sens. Environ. 1998, 64, 234–253. [Google Scholar] [CrossRef]
- Knyazikhin, Y.; Schull, M.A.; Stenberg, P.; Mõttus, M.; Rautiainen, M.; Yang, Y.; Marshak, A.; Carmona, P.L.; Kaufmann, R.K.; Lewis, P.; et al. Hyperspectral remote sensing of foliar nitrogen content. Proc. Natl. Acad. Sci. USA 2013, 110, E185–E192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vierling, L.A.; Deering, D.W.; Eck, T.F. Differences in arctic tundra vegetation type and phenology as seen using bidirectional radiometry in the early growing season. Remote Sens. Environ. 1997, 60, 71–82. [Google Scholar] [CrossRef]
- Molvar, E.M.; Bowyer, R.T.; Van Ballenberghe, V. Moose herbivory, browse quality, and nutrient cycling in an Alaskan treeline community. Oecologia 1993, 94, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Blair, R.M.; Alcaniz, R.; Harrell, A. Shade intensity influences the nutrient quality and digestibility of southern deer browse leaves. J. Range Manag. 1983, 36, 257–264. [Google Scholar] [CrossRef]
- Zengeya, F.M.; Mutanga, O.; Murwira, A. Linking remotely sensed forage quality estimates from WorldView-2 multispectral data with cattle distribution in a savanna landscape. Int. J. Appl. Earth Obs. Geoinf. 2012, 21, 513–524. [Google Scholar] [CrossRef]
- Ramoelo, A.; Cho, M.A.; Mathieu, R.; Madonsela, S.; Van De Kerchove, R.; Kaszta, Z.; Wolff, E. Monitoring grass nutrients and biomass as indicators of rangeland quality and quantity using random forest modelling and WorldView-2 data. Int. J. Appl. Earth Obs. Geoinf. 2015, 43, 43–54. [Google Scholar] [CrossRef]
- Hively, W.D.; Lamb, B.T.; Daughtry, C.S.T.; Shermeyer, J.; Mccarty, G.W.; Quemada, M. Mapping crop residue and tillage intensity using WorldView-3 Satellite shortwave infrared residue indices. Remote Sens. 2018, 10, 1657. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Levin, N.; Seabrook, L.; Moore, B.; Mcalpine, C. Mapping foliar nutrition using WorldView-3 and WorldView-2 to assess Koala habitat suitability. Remote Sens. 2019, 11, 215. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Budkewitsch, P.; Treitz, P. Examining spectral reflectance features related to Arctic percent vegetation cover: Implications for hyperspectral remote sensing of Arctic tundra. Remote Sens. Environ. 2017, 192, 58–72. [Google Scholar] [CrossRef]
- Langford, Z.; Kumar, J.; Hoffman, F.M.; Norby, R.J.; Wullschleger, S.D.; Sloan, V.L.; Iversen, C.M. Mapping Arctic plant functional type distributions in the Barrow Environmental Observatory using WorldView-2 and LiDAR datasets. Remote Sens. 2016, 8, 733. [Google Scholar] [CrossRef] [Green Version]
- Dumroese, R.K.; Pinto, J.R.; Montville, M.E. Using container weights to determine irrigation needs: A simple method. Nativ. Plants J. 2015, 16, 67–71. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Glozer, K. Protocol for Leaf Image Analysis—Surface Area. 2008. Available online: http//ucanr.edu/sites/fruittree/files/49325.pdf (accessed on 10 March 2020).
- R-Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.r-project.org/ (accessed on 10 January 2019).
- Welch, B.L. The generalization of students’ problem when several different population variances are involved. Biometrika 1947, 34, 28–35. [Google Scholar] [PubMed]
- Tukey, J.W. Comparing individual means in the analysis of variance. Biometrics 1949, 5, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Cavanaugh, J.E. Unifying the derivations for the Akaike and corrected Akaike information criteria. Stat. Probab. Lett. 1997, 33, 201–208. [Google Scholar] [CrossRef]
- Barton, K.; Barton, M.K. Package ‘MuMIn’. 2015. Available online: https://cran.r-project.org/web/packages/MuMIn/MuMIn.pdf (accessed on 24 March 2020).
- Smith, A.M.S.; Wooster, M.J.; Drake, N.A.; Dipotso, F.M.; Falkowski, M.J.; Hudak, A.T. Testing the potential of multi-spectral remote sensing for retrospectively estimating fire severity in African Savannahs. Remote Sens. Environ. 2005, 97, 92–115. [Google Scholar] [CrossRef] [Green Version]
- Trigg, S.; Flasse, S. Characterizing the spectral-temporal response of burned savannah using in situ spectroradiometry and infrared thermometry. Int. J. Remote Sens. 2000, 21, 3161–3168. [Google Scholar] [CrossRef]
- Heiskanen, J. Estimating aboveground tree biomass and leaf area index in a mountain birch forest using ASTER satellite data. Int. J. Remote Sens. 2006, 27, 1135–1158. [Google Scholar] [CrossRef]
- Lin, C.H.; McGraw, M.L.; George, M.F.; Garrett, H.E. Nutritive quality and morphological development under partial shade of some forage species with agroforestry potential. Agrofor. Syst. 2001, 53, 269–281. [Google Scholar] [CrossRef]
- Juutinen, S.; Virtanen, T.; Kondratyev, V.; Laurila, T.; Linkosalmi, M.; Mikola, J.; Nyman, J.; Räsänen, A.; Tuovinen, J.P.; Aurela, M. Spatial variation and seasonal dynamics of leaf-Area index in the arctic tundra-implications for linking ground observations and satellite images. Environ. Res. Lett. 2017, 12, 095002. [Google Scholar] [CrossRef]
- Hanley, T.A. Physical and chemical response of understory vegetation to deer use in southeastern Alaska. Can. J. Res. 1987, 17, 195–199. [Google Scholar] [CrossRef]
- Lenart, E.A.; Bowyer, R.T.; Hoef, J.V.; Ruess, R.W. Climate change and caribou: Effects of summer weather on forage. Can. J. Zool. 2002, 80, 664–678. [Google Scholar] [CrossRef]
- Moore, K.J.; Jung, H.J.G. Lignin and fiber digestion. Rangel. Ecol. Manag. Range Manag. Arch. 2001, 54, 420–430. [Google Scholar] [CrossRef]
- Osier, T.L.; Lindroth, R.L. Effects of light and nutrient availability on Aspen: Growth, phytochemistry, and insect performance. J. Chem. Ecol. 1999, 25, 1687–1714. [Google Scholar]
- Weladji, R.B.; Klein, D.R.; Holand, Ø. Mysterud, a Comparative response of Rangifer tarandus and other northern ungulates to climatic variability. Rangifer 2002, 22, 33–50. [Google Scholar] [CrossRef] [Green Version]
- Elmendorf, S.C.; Henry, G.H.R.; Hollister, R.D.; Björk, R.G.; Bjorkman, A.D.; Callaghan, T.V.; Collier, L.S.; Cooper, E.J.; Cornelissen, J.H.C.; Day, T.A.; et al. Global assessment of experimental climate warming on tundra vegetation: Heterogeneity over space and time. Ecol. Lett. 2012, 15, 164–175. [Google Scholar] [CrossRef]
- Guanter, L.; Kaufmann, H.; Segl, K.; Foerster, S.; Rogass, C.; Chabrillat, S.; Kuester, T.; Hollstein, A.; Rossner, G.; Chlebek, C.; et al. The EnMAP spaceborne imaging spectroscopy mission for earth observation. Remote Sens. 2015, 7, 8830–8857. [Google Scholar] [CrossRef] [Green Version]
- Jensen, J.L.R.; Humes, K.S.; Vierling, L.A.; Hudak, A.T. Discrete return lidar-based prediction of leaf area index in two conifer forests. Remote Sens. Environ. 2008, 112, 3947–3957. [Google Scholar] [CrossRef] [Green Version]
- Pope, G.; Treitz, P. Leaf Area Index (LAI) estimation in boreal mixedwood forest of Ontario, Canada using Light detection and ranging (LiDAR) and worldview-2 imagery. Remote Sens. 2013, 5, 5040–5063. [Google Scholar] [CrossRef] [Green Version]
- Somers, B.; Asner, G.P.; Tits, L.; Coppin, P. Endmember variability in Spectral Mixture Analysis: A review. Remote Sens. Environ. 2011, 115, 1603–1616. [Google Scholar] [CrossRef]
- Adams, J.B.; Smith, M.O. Spectral mixture modeling: A new analysis of rock and soil types at the Viking Lander 1 site. J. Geophys. Res. 1986, 91, 8098–8112. [Google Scholar] [CrossRef]
- Eitel, J.U.H.; Long, D.S.; Gessler, P.E.; Hunt, E.R.; Brown, D.J. Sensitivity of ground-based remote sensing estimates of wheat chlorophyll content to variation in soil reflectance. Soil Sci. Soc. Am. 2009, 73, 1715–1723. [Google Scholar] [CrossRef] [Green Version]
- Darvishzadeh, R.; Skidmore, A.; Atzberger, C.; van Wieren, S. Estimation of vegetation LAI from hyperspectral reflectance data: Effects of soil type and plant architecture. Int. J. Appl. Earth Obs. Geoinf. 2008, 10, 358–373. [Google Scholar] [CrossRef]




| Models | R2 | RMSE | AICc | ΔAICc | LOOCV Slope | LOOCV | ΔLOOCV |
|---|---|---|---|---|---|---|---|
| Hemicellulose (HMC) | |||||||
| SVI | 0.68 | 0.42 | 34.62 | - | 0.66 | 0.73 | - |
| SVI+ LA Sun | 0.74 | 0.37 | 31.45 | −3.17 | 0.75 | 0.69 | −4% |
| SVI+ LA Shade | 0.69 | 0.41 | 35.48 | +0.86 | 0.68 | 0.77 | +4% |
| SVI+ LA Total | 0.72 | 0.39 | 32.63 | −1.99 | 0.71 | 0.73 | 0 |
| Cellulose (CLL) | |||||||
| SVI | 0.61 | 0.42 | 34.48 | - | 0.59 | 0.78 | - |
| SVI+ LA Sun | 0.59 | 0.43 | 37.14 | +2.69 | 0.56 | 0.80 | +2% |
| SVI+ LA Shade | 0.60 | 0.42 | 36.90 | +2.42 | 0.55 | 0.81 | +3% |
| SVI+ LA Total | 0.60 | 0.42 | 36.97 | +2.49 | 0.56 | 0.81 | +3% |
| Neutral detergent fiber (NDF) | |||||||
| SVI | 0.51 | 0.99 | 75.30 | - | 0.57 | 0.79 | - |
| SVI+ LA Sun | 0.55 | 0.93 | 74.82 | −0.40 | 0.59 | 0.81 | +2% |
| SVI+ LA Shade | 0.67 | 0.80 | 67.65 | −7.65 | 0.62 | 0.87 | +8% |
| SVI+ LA Total | 0.62 | 0.86 | 71.02 | −4.28 | 0.60 | 0.85 | +6% |
| Acid detergent fiber (ADF) | |||||||
| SVI | 0.56 | 0.61 | 51.31 | - | 0.53 | 0.75 | - |
| SVI+ LA Sun | 0.53 | 0.61 | 54.15 | +2.84 | 0.48 | 0.76 | +1% |
| SVI+ LA Shade | 0.53 | 0.61 | 54.20 | +2.89 | 0.50 | 0.75 | 0 |
| SVI+ LA Total | 0.53 | 0.61 | 54.17 | +2.86 | 0.49 | 0.76 | +1% |
| Acid detergent lignin (ADL) | |||||||
| SVI | 0.48 | 0.25 | 9.46 | - | 0.49 | 0.83 | - |
| SVI+ LA Sun | 0.49 | 0.25 | 10.86 | +1.46 | 0.51 | 0.82 | −1% |
| SVI+ LA Shade | 0.51 | 0.24 | 9.87 | +0.41 | 0.52 | 0.82 | −1% |
| SVI+ LA Total | 0.51 | 0.24 | 10.07 | +0.61 | 0.52 | 0.82 | −1% |
| Acid detergent ash (AIA) | |||||||
| SVI | 0.58 | 0.04 | −83.54 | - | 0.57 | 0.81 | - |
| SVI+ LA Sun | 0.56 | 0.04 | −82.77 | +0.84 | 0.55 | 0.81 | 0 |
| SVI+ LA Shade | 0.58 | 0.04 | −81.94 | +1.60 | 0.58 | 0.81 | 0 |
| SVI+ LA Total | 0.56 | 0.04 | −80.73 | +2.81 | 0.55 | 0.81 | 0 |
| Models | R2 | RMSE | AICc | ΔAICc | LOOCV Slope | LOOCV | ΔLOOCV |
|---|---|---|---|---|---|---|---|
| Hemicellulose (HMC) | |||||||
| SVI | 0.32 | 0.62 | 52.65 | - | 0.27 | 0.52 | - |
| SVI+ LA Sun | 0.41 | 0.56 | 50.74 | −1.91 | 0.39 | 0.72 | +20% |
| SVI+ LA Shade | 0.40 | 0.57 | 51.38 | −1.27 | 0.38 | 0.72 | +20% |
| SVI+ LA Total | 0.45 | 0.55 | 49.29 | −3.36 | 0.43 | 0.72 | +20% |
| Cellulose (CLL) | |||||||
| SVI | 0.25 | 0.59 | 50.10 | - | 0.22 | 0.45 | - |
| SVI+ LA Sun | 0.23 | 0.59 | 52.44 | +2.34 | 0.20 | 0.50 | +5% |
| SVI+ LA Shade | 0.22 | 0.59 | 52.66 | +2.56 | 0.18 | 0.49 | +4% |
| SVI+ LA Total | 0.23 | 0.58 | 52.44 | +2.34 | 0.19 | 0.49 | +4% |
| Neutral detergent fiber (NDF) | |||||||
| SVI | 0.31 | 1.18 | 83.33 | - | 0.26 | 0.67 | - |
| SVI+ LA Sun | 0.33 | 1.14 | 84.34 | +1.01 | 0.31 | 0.71 | +4% |
| SVI+ LA Shade | 0.45 | 1.03 | 79.82 | −3.51 | 0.43 | 0.78 | +11% |
| SVI+ LA Total | 0.39 | 1.08 | 82.09 | −1.24 | 0.37 | 0.78 | +11% |
| Acid detergent fiber (ADF) | |||||||
| SVI | 0.30 | 0.76 | 62.10 | - | 0.28 | 0.57 | - |
| SVI+ LA Sun | 0.34 | 0.72 | 62.40 | +0.30 | 0.35 | 0.66 | +9% |
| SVI+ LA Shade | 0.40 | 0.69 | 60.43 | −2.33 | 0.38 | 0.76 | +19% |
| SVI+ LA Total | 0.38 | 0.70 | 60.92 | −2.82 | 0.38 | 0.73 | +16% |
| Acid detergent lignin (ADL) | |||||||
| SVI | 0.34 | 0.28 | 15.05 | - | 0.33 | 0.70 | - |
| SVI+ LA Sun | 0.32 | 0.28 | 17.51 | +2.46 | 0.32 | 0.71 | +1% |
| SVI+ LA Shade | 0.34 | 0.28 | 17.00 | +1.95 | 0.33 | 0.71 | +1% |
| SVI+ LA Total | 0.33 | 0.28 | 17.19 | +2.14 | 0.33 | 0.69 | −1% |
| Acid detergent ash (AIA) | |||||||
| SVI | 0.13 | 0.05 | −66.35 | - | 0.11 | 0.31 | - |
| SVI+ LA Sun | 0.09 | 0.05 | −63.45 | +2.90 | 0.08 | 0.31 | 0 |
| SVI+ LA Shade | 0.10 | 0.05 | −63.78 | +2.57 | 0.09 | 0.31 | 0 |
| SVI+ LA Total | 0.09 | 0.05 | −63.54 | +2.81 | 0.08 | 0.28 | −3% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jennewein, J.S.; Eitel, J.U.H.; Pinto, J.R.; Vierling, L.A. Toward Mapping Dietary Fibers in Northern Ecosystems Using Hyperspectral and Multispectral Data. Remote Sens. 2020, 12, 2579. https://doi.org/10.3390/rs12162579
Jennewein JS, Eitel JUH, Pinto JR, Vierling LA. Toward Mapping Dietary Fibers in Northern Ecosystems Using Hyperspectral and Multispectral Data. Remote Sensing. 2020; 12(16):2579. https://doi.org/10.3390/rs12162579
Chicago/Turabian StyleJennewein, Jyoti S., Jan U.H. Eitel, Jeremiah R. Pinto, and Lee A. Vierling. 2020. "Toward Mapping Dietary Fibers in Northern Ecosystems Using Hyperspectral and Multispectral Data" Remote Sensing 12, no. 16: 2579. https://doi.org/10.3390/rs12162579
APA StyleJennewein, J. S., Eitel, J. U. H., Pinto, J. R., & Vierling, L. A. (2020). Toward Mapping Dietary Fibers in Northern Ecosystems Using Hyperspectral and Multispectral Data. Remote Sensing, 12(16), 2579. https://doi.org/10.3390/rs12162579