1. Introduction
Non-native grasses cover a large extent of southern California that was once covered by extensive stands of native coastal sage scrub (CSS). CSS is a threatened habitat, where restoration efforts are often hindered by the dominance of exotic grasses. Native plant communities are slow to recover once exotic grasses have established [
1,
2]. CSS is a vegetation community type of high priority for conservation because of its high degree of biological diversity and marked reduction in areal extent. Management of CSS habitats is critical for the conservation of native vegetation, which supports several rare and threatened plant and animal species [
3,
4]. Mapping large regions of CSS habitat at the growth form level and tracking changes in shrub cover over time will create a baseline of CSS conditions and develop a trajectory for future recovery and restoration [
5].
Two main approaches tend to be utilized for monitoring CSS communities, vegetation-community-type mapping and field sampling of individual sample plots. Vegetation community maps provide information about the distribution of community types over broad regions, while field sampling allows for species-specific information for small sample plots [
6,
7]. An alternative to these methods is mapping vegetation at the growth-form level using high-spatial-resolution remotely sensed imagery and a semi-automated image classification. This allows for more detailed mapping of vegetation composition and structure and their changes than community type mapping, but still enables coverage of large extents. Spatially exhaustive vegetation maps are therefore more suitable for regional planning than plot-based information [
6]. However, such maps are not independent of, nor will they replace the need for field-based approaches.
CSS vegetation on San Clemente Island (SCI), (located approximately 100 km off the coast of southern California), has been heavily impacted due to prolonged overgrazing and military-related disturbances. Since the removal of feral herbivores in the early 1990s, CSS has begun to recover, providing an optimal study site for observing restoration due to current limited disturbances [
8]. Shrub cover provides critical habitat for threatened and endangered bird species, and biologists and resource managers are interested in mapping shrub cover in order to understand if current management actions are facilitating shrub recovery.
Vegetation growth forms refer to general classes for plant species that are grouped based on similarities in structure, function and growth dynamics. Growth forms composing shrubland landscapes of southern California include scattered tree, true shrub, sub-shrub, herbs (grasses and forbs), and bare cover types [
7]. Previous studies on shrublands have indicated that cover estimates of vegetation growth forms are an effective gauge of habitat health and integrity [
9,
10]. Changes in shrub and herbaceous cover can be indicative of the internal conditions within vegetation communities within Mediterranean-type ecosystems.
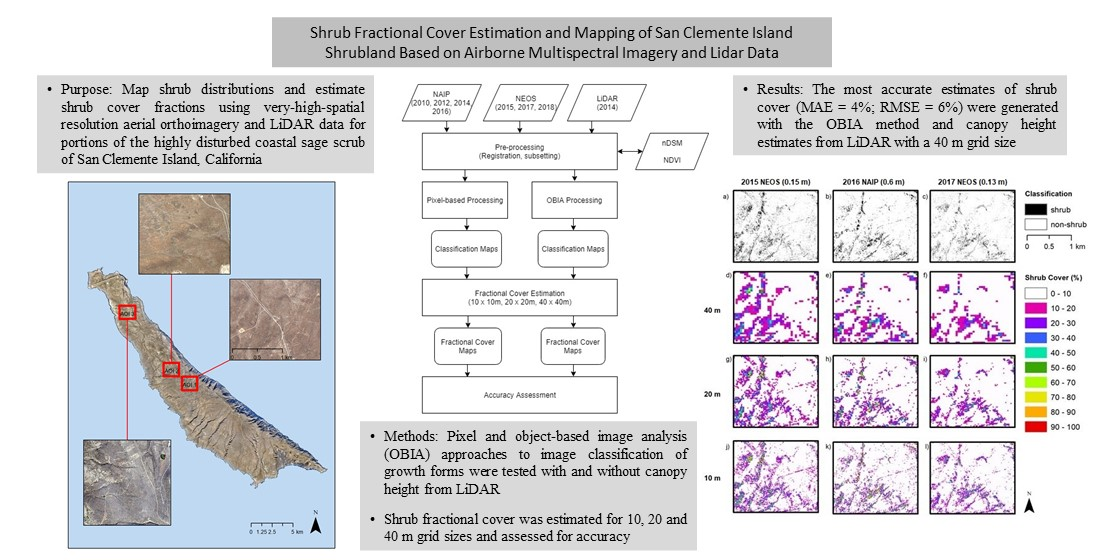
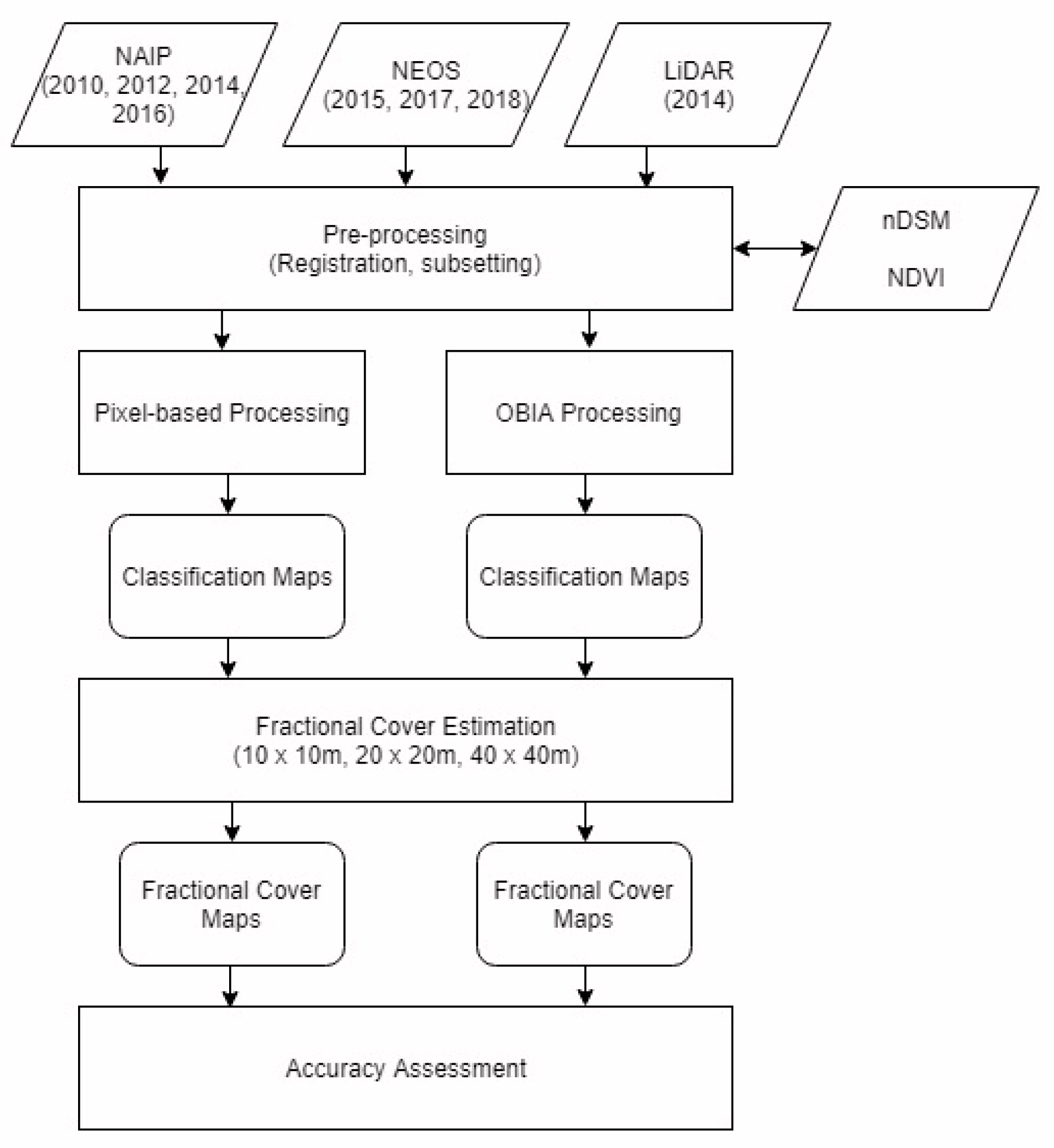
The primary objective of this study is to test the accuracy of estimating and mapping shrub cover fractions across SCI based on the classification of several very-high-spatial-resolution multispectral orthoimagery types (mosaics of orthorectified imagery) and light detection and ranging (LiDAR) data. The aerial orthoimagery data sets were obtained from the United States National Agricultural Imagery Program (NAIP) and from a commercial provider, Near Earth Observation System, Ltd. (NEOS). An emphasis was placed on object-based image analysis approaches. An analysis of multi-temporal aerial orthoimagery data sets captured cover over a nine-year period (2010-2018) was conducted in the context of mapping shrub cover.
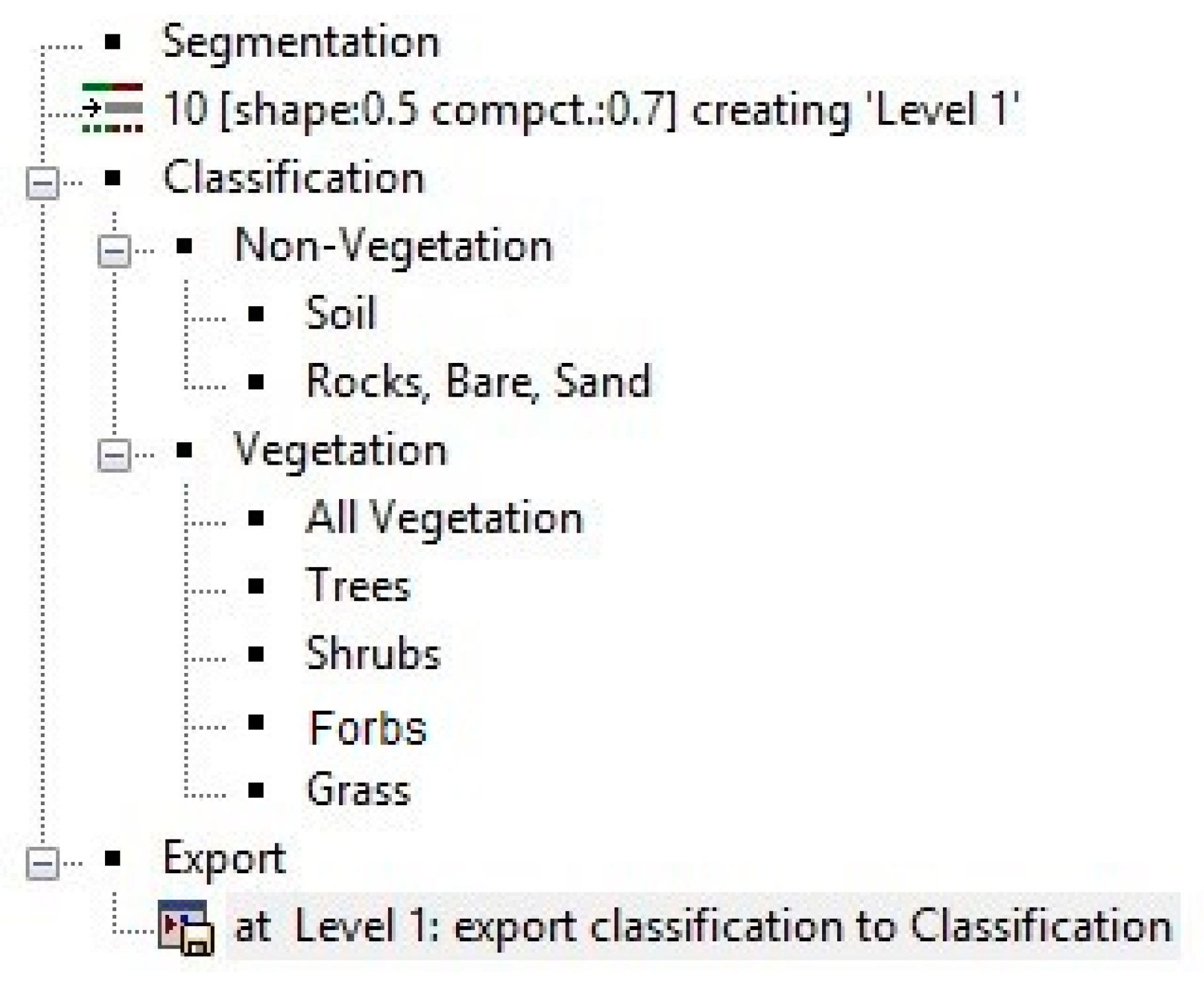
Object-based image analysis (OBIA) approaches have proven to be useful for the classification and mapping of shrubs [
9,
11,
12]. Hamada et al. [
9] and Laliberte et al. [
11] found that OBIA routines tend to yield higher accuracy map products than other processing methods, including per-pixel classifiers and spectral mixture models. Hamada et al. [
9] assessed both high- and low-resolution processing methods to generate growth form maps in a CSS community. The results showed that OBIA yielded lower error than pixel-based classification approaches based on maximum likelihood and artificial neural network routines, and spectral mixture analysis applied to high and moderate spatial resolution satellite data. Laliberte et al. [
11] utilized a similar OBIA process for mapping growth forms in rangelands and further broke down the classification to the species level for shrubs and grasses. Species-level classifications were attained for the dominant shrub species, while grasses were especially difficult to separate by species [
9,
11].
Another advantage of OBIA is the ability for cost-effective long-term ecological monitoring of CSS communities. Stow et al. [
13] demonstrated that OBIA methods are more appropriate for monitoring shrub cover changes than pixel-based methods, when applied to high-spatial-resolution and multi-temporal image datasets. Through multi-temporal analyses, life form cover changes can be tracked over time. Laliberte et al. [
11] also highlights the advantage of more efficient remote sensing methods over ground-based measurements. However, there is a trade-off between the accuracy and computational cost of OBIA methods [
14]. OBIA is semi-automated; therefore, it is often highly dependent on expert knowledge and user input for the development of the segmentation and classification scheme. However, machine learning is also a method that shows a lot of promise and researchers are increasingly using machine learning techniques for image classification. Machine learning may allow for more accurate feature extraction and classification of National Agricultural Imagery Program (NAIP) imagery. However, machine learning algorithms require a large amount of very accurately labeled training inputs [
15], which typically involves some form of semi-automatic classification and subsequent manual digitizing for accurate label generation. Extensive field sampling is challenging to conduct for places such as San Clemente Island that has restricted access.
Hamada et al. [
5,
7] utilized 25 × 25 m and 50 × 50 m grids for fractional cover estimation. Spatial sampling units 25 × 25 m in size proved to be more appropriate for estimating fractional cover within CSS communities for comparative reasons, while a 50 × 50 m sampling unit is appropriate for estimating growth form cover from a management perspective. These plot sizes are the most common sampling units used for studying CSS communities and the maximum area used by resource managers for restoration efforts such as eradicating invasive plants and planting native species [
5]. However, different size sampling units may be more appropriate for the heavily disturbed and recovering state of shrublands on SCI.
Based on the above, we address the following research questions in this study:
Which data inputs and image processing approaches generate the most accurate shrub fractional cover maps?
How does the incorporation of nDSM data, that enables quantification of shrub and tree canopy heights, affect the accuracy of the map products?
Does an OBIA approach yield more reliable shrub cover estimates than a pixel-based approach when applied to NAIP (0.6 to 1 m spatial resolution) orthoimage datasets that are readily available for SCI since at least 2010?
What is the most appropriate grid size for quantifying shrub cover fractions in the context of potential future monitoring, given the relatively disturbed nature of shrublands on SCI?
2. Study Area
SCI is the southernmost of the Channel Islands off the southern California coast. The island trends 34 km northwest to southeast and ranges from 2 to 7 km wide, and has the highest level of endemism of the Channel Islands with 47 species endemic to the island group and 17 species endemic to SCI. Six of these species are classified as endangered or threatened and have faced difficulties re-establishing due to anthropogenic disturbances [
8].
Vegetation on SCI is composed of two major community types: grasslands and CSS. There are 382 plant species present, 272 of which are native [
8]. SCI has the highest plant endemism of the Channel Islands with 47 species endemic to the island group and 17 to SCI [
16]. The upland terrain on the central and eastern portions of the island are dominated by non-native annual grasses such as
Avena ssp. (wild oats) and
Bromus ssp. (bromes) [
17]. The coastal sage scrub found on the low-elevation areas is categorized as maritime succulent scrub, including
Opuntia littoralis (prickly pear cactus),
Cylindropuntia prolifera (cholla),
Lycium californicum (box thorn), and other drought-resistant species. Although they once had a wider distribution, CSS habitats and trees are now confined to the moist environments of the canyons due to historic overgrazing [
17,
18].
CSS at the lower elevations of the island tends to be found on isolated coastal bluffs and marine terraces. Vegetation is characterized by shrub and succulent species, including
L. californicum,
Rhus integrifolia,
O. littoralis and
C. prolifera (
Figure 1).
L. californicum is a low growing shrub found primarily on the lower terraces of the western shore.
O. littoralis is prevalent on the terrace faces, which has served as shelter for shrub seedlings from herbivores.
C. prolifera is found on the slopes of the southern end of the island [
8].
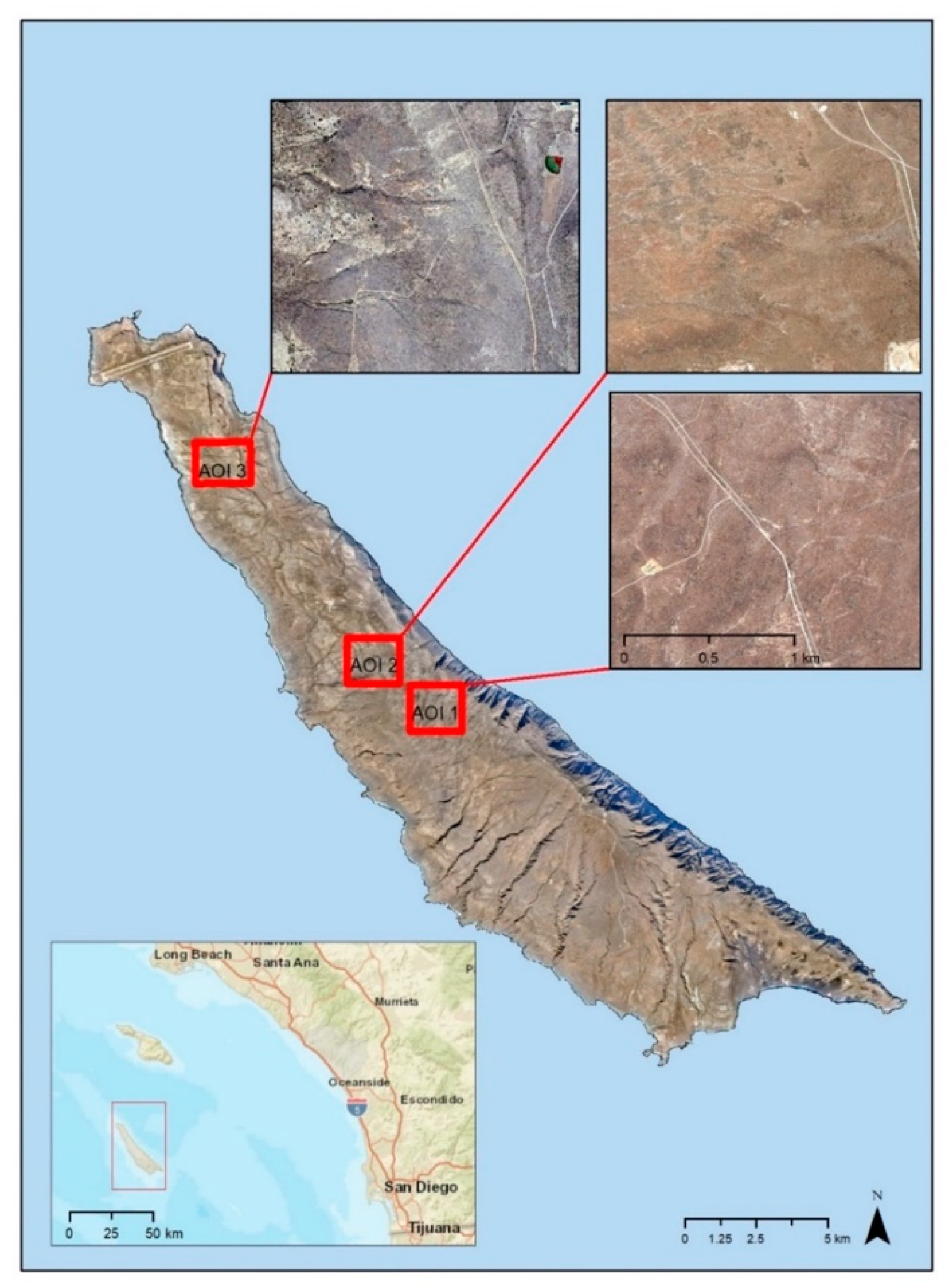
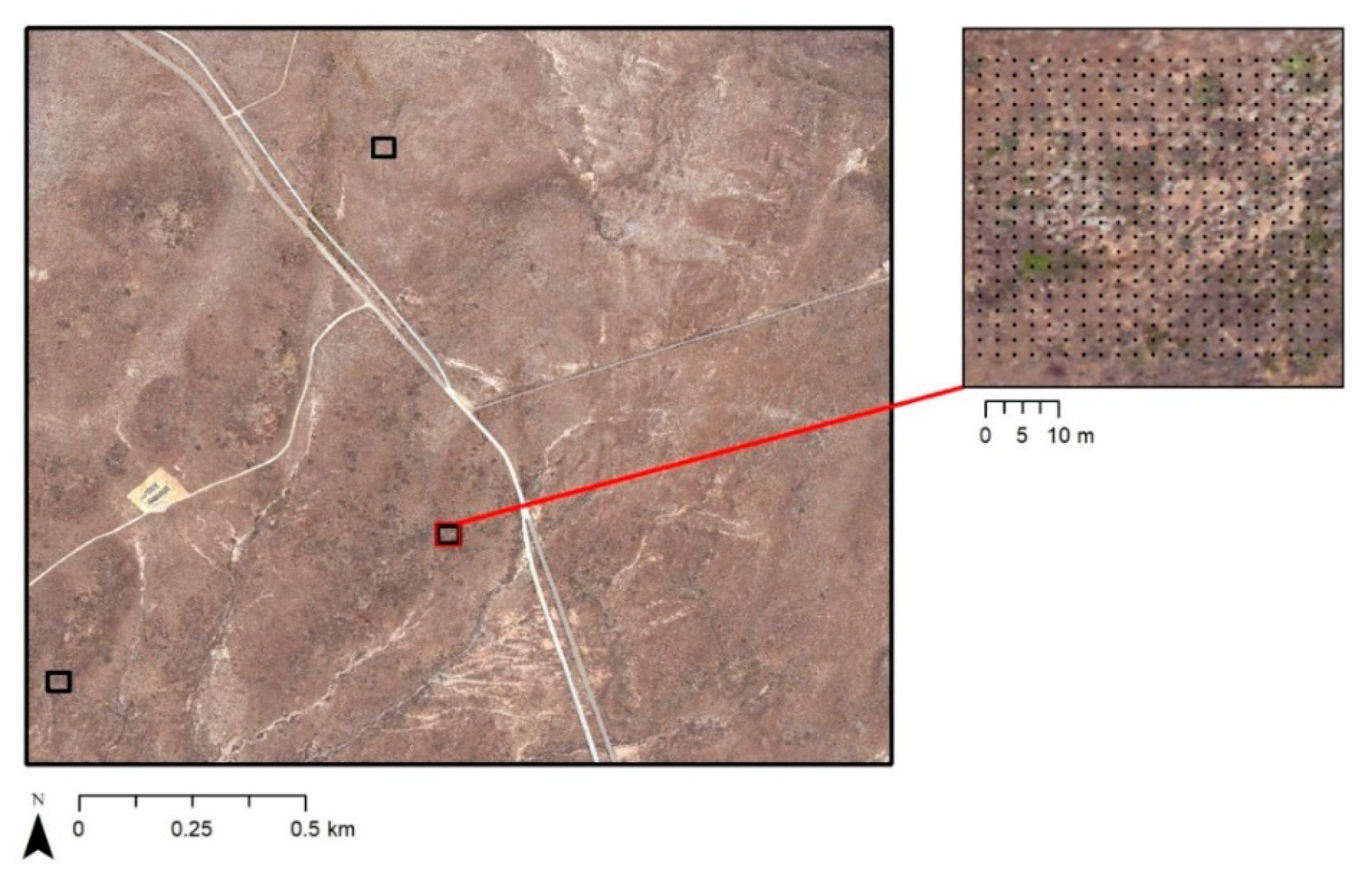
This shrub mapping research is focused on two general study areas on the island called Stone and Flasher. Two areas of interest (AOIs) were selected within the Stone study area on the eastern side of the island (
Figure 1). These AOIs were selected based on the relative diversity of vegetation types, as well as the accessibility for performing ground-based accuracy assessments. A third AOI was selected within the Flasher study area located on the northwestern part of SCI to represent a range of shrub cover and diversity of shrub species not represented within the Stone study area.
AOIs 1 and 2 within the Stone study area are dominated by grasslands and a high cover of
O. littoralis. The soil on the eastern slopes is composed of loam. AOI 3 in the Flasher study area is located the sand dunes on the northwestern coast. This area contains higher cover of
L. californicum and
R. intergrifolia. Soil in the Flasher study area is composed of a sandy loam with sand dunes on the coastline [
8].
The island vegetation experienced severe degradation due to overgrazing and mechanical disturbances by feral goats, sheep and cattle. Sheep and cattle ranching ended in 1934, though feral herbivores were not completely eradicated until the early 1990s. The U.S. Navy has owned and operated on SCI since 1934. While disturbances to the island have been widespread, most of recent military-related disturbances have occurred in the southern third of the island. Since 1996, half of the fires occurring on the island have occurred within the Shore Bombardment Area (SHOBA). However, these fires account for approximately 90% of the total acreage burned [
8].
SCI has a Mediterranean-type climate which is characterized by cool, wet winters and hot, dry summers. On average, 95% of rainfall occurs between November and April, with the most occurring in January and February. June, July and August are the driest months with only 1% of the annual mean rainfall. Regional rainfall patterns are highly variable and unpredictable. Multi-year droughts punctuated by wet years are not uncommon [
19].
Both precipitation and the time of year have an impact on the spectral and physical properties of vegetation. Seasonal changes in leaf color and leaf area index can show inconsistent vegetation cover when comparing imagery captured during different seasons. Sims et al. [
20] found that a greater decline in normalized difference vegetation index (NDVI) than canopy chlorophyll index (CCI) during extreme drought, suggesting that canopy structure is affected more than leaf greenness.
Table 1 shows the total precipitation for one, three and six months prior to the collection of aerial imagery used in this study [
8,
21,
22]. The amount of precipitation leading into the growing season influences the greenness of different vegetation types and therefore the ability to discriminate and accurately classify them.
5. Discussion
5.1. Seasonality
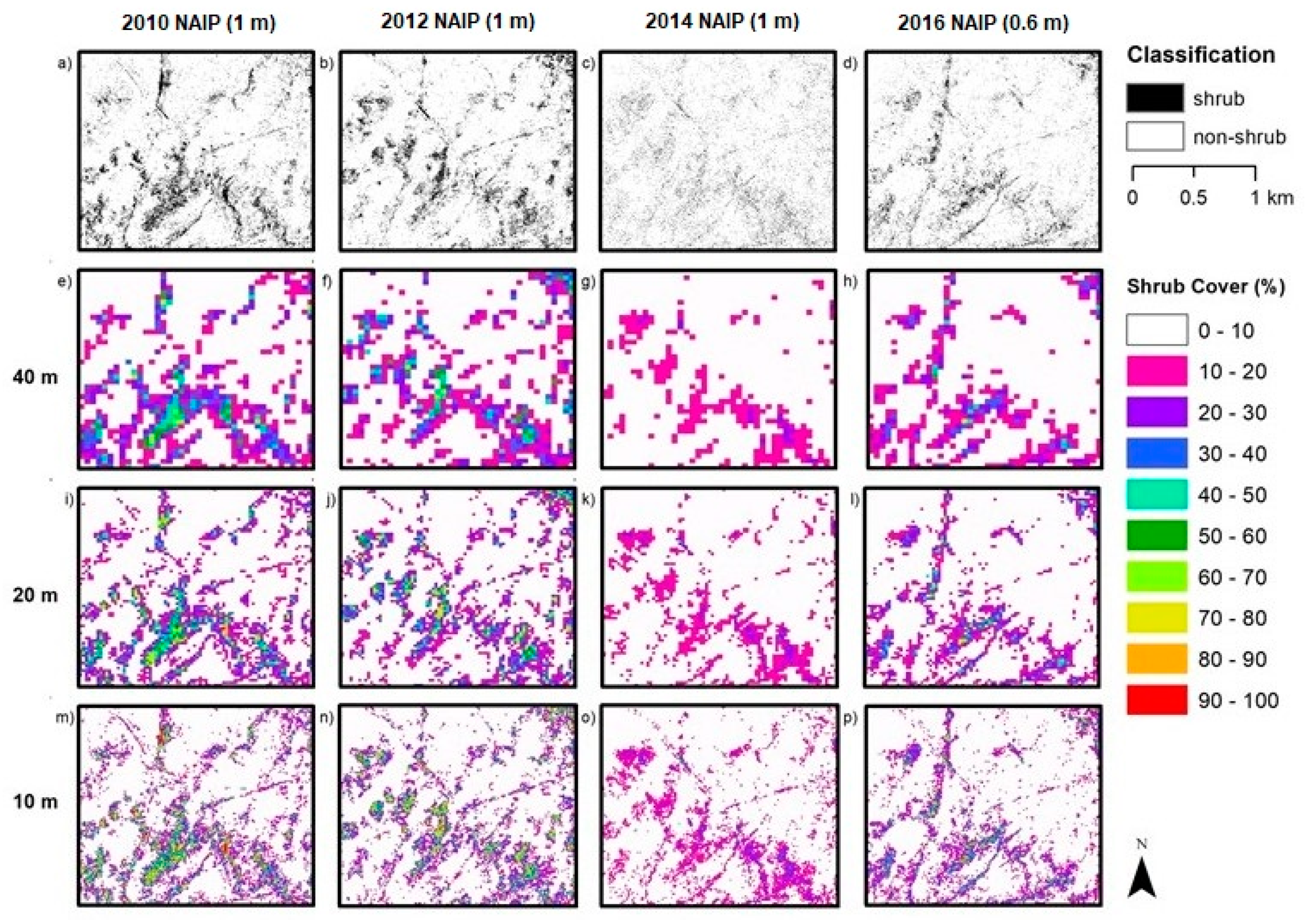
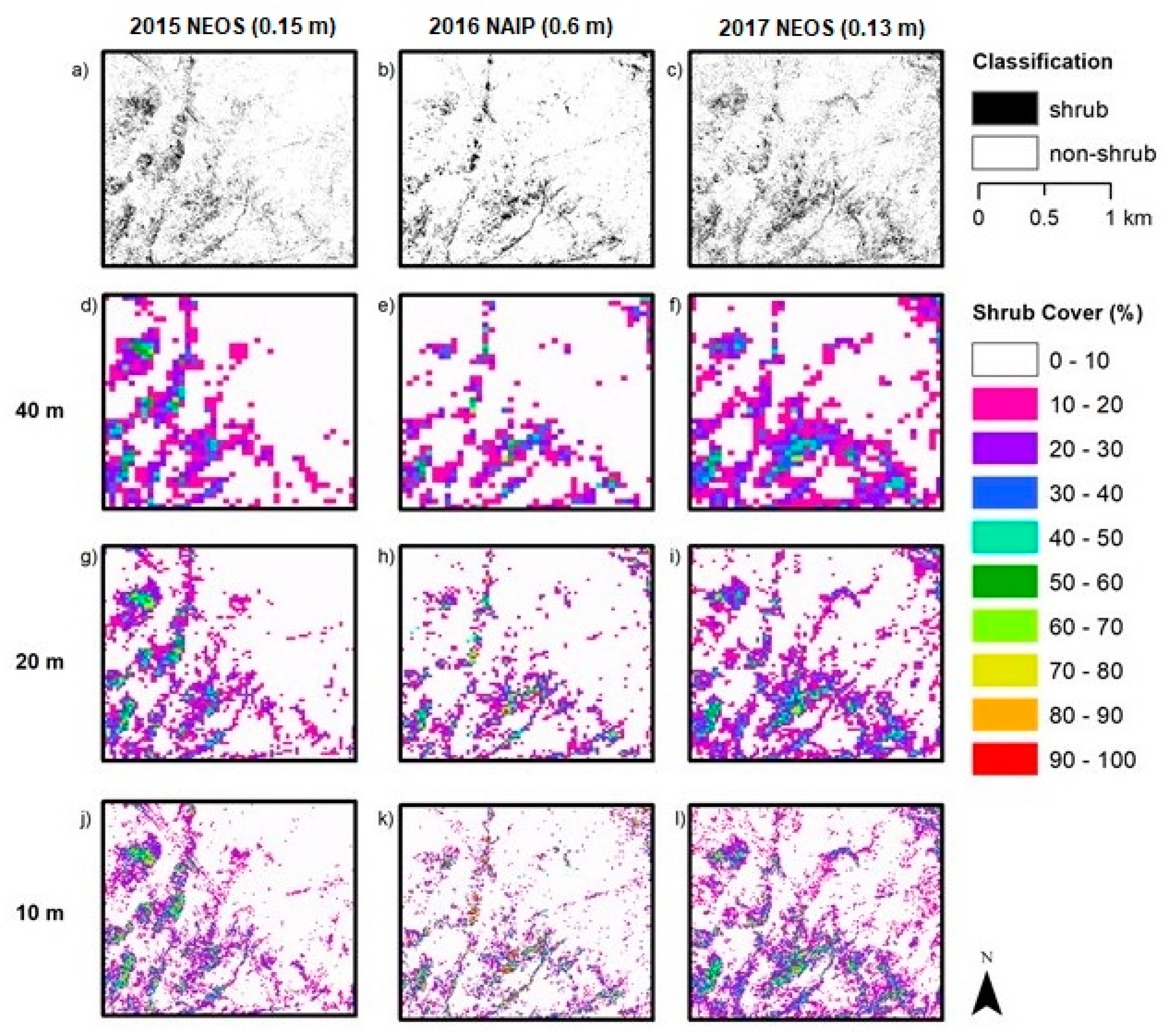
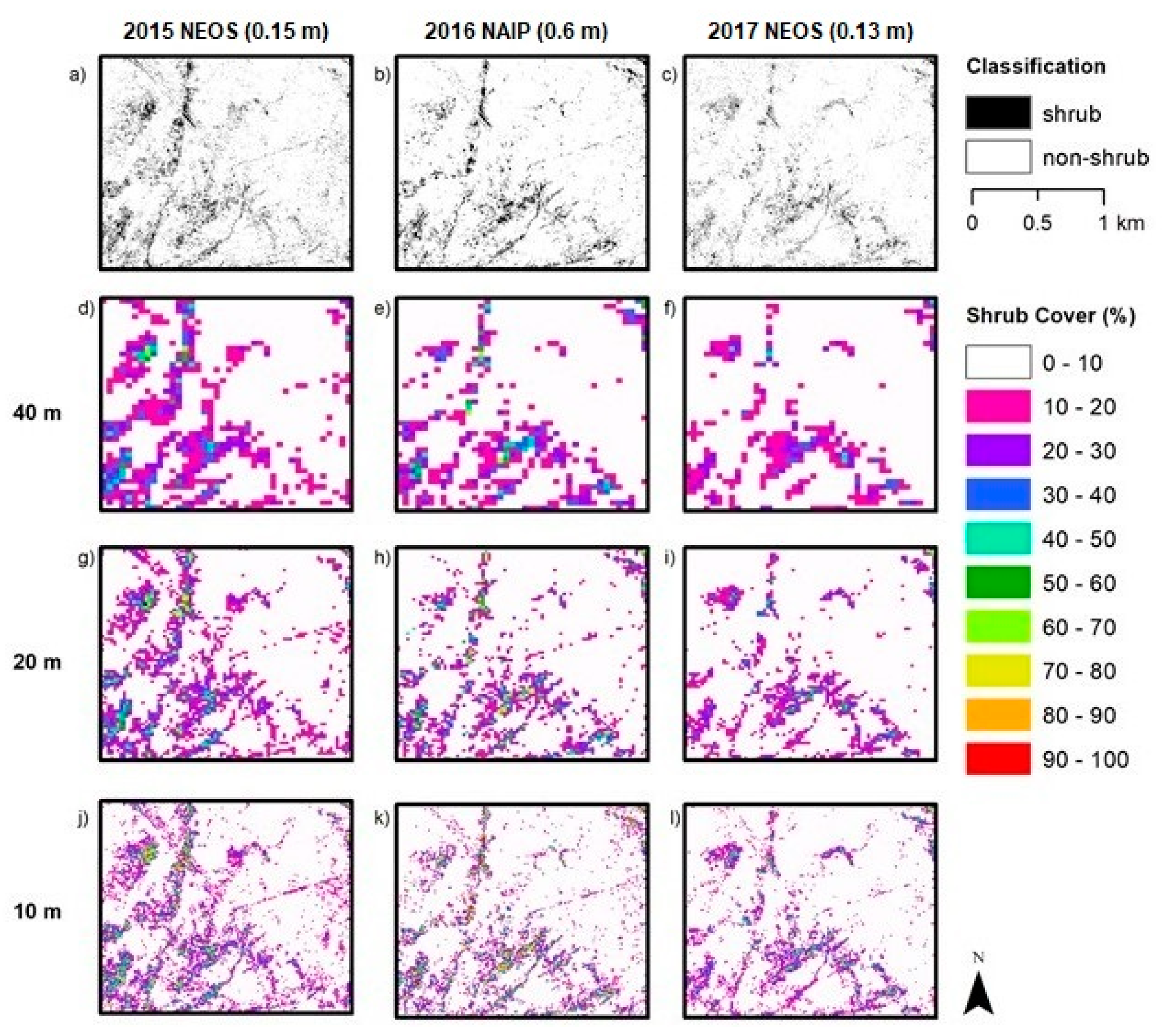
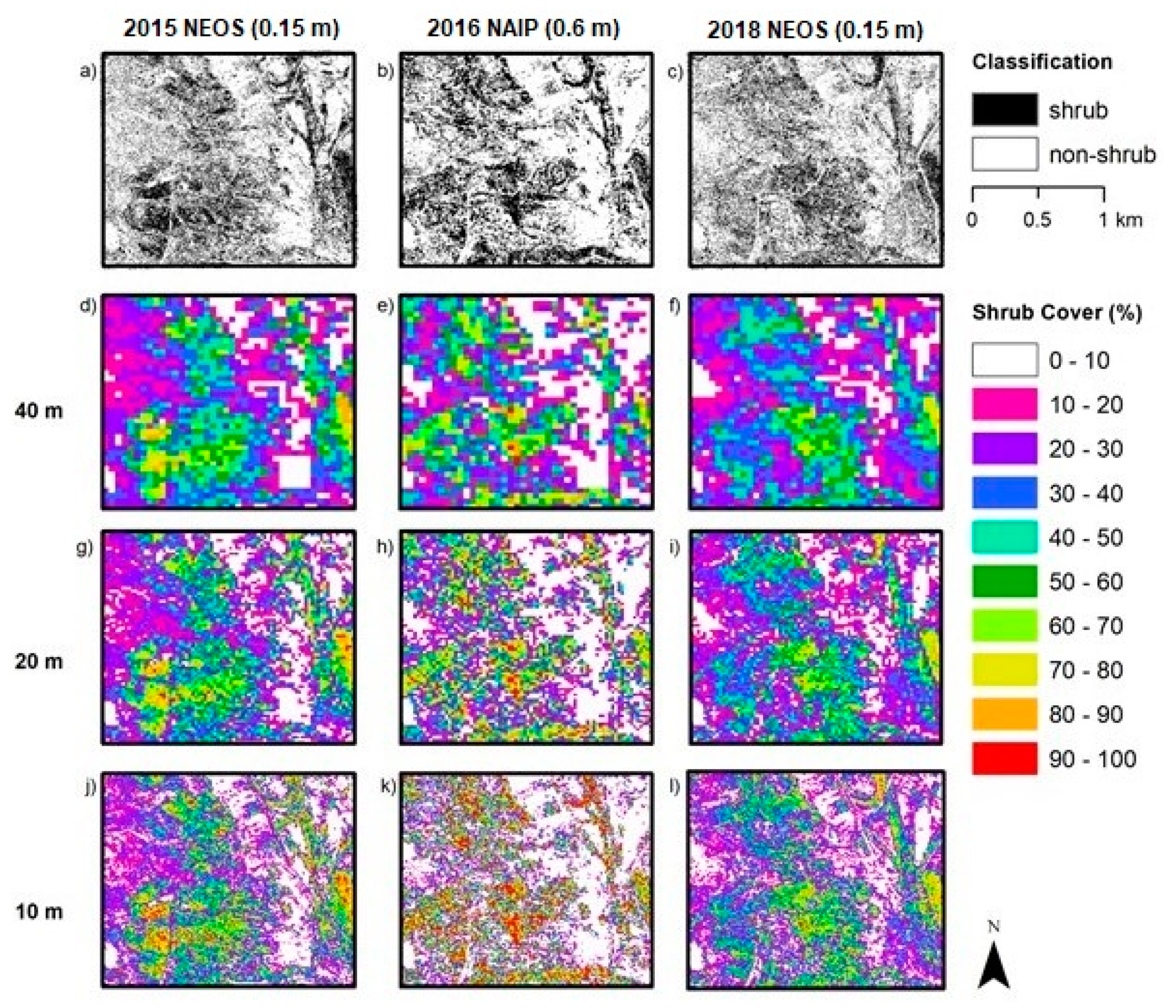
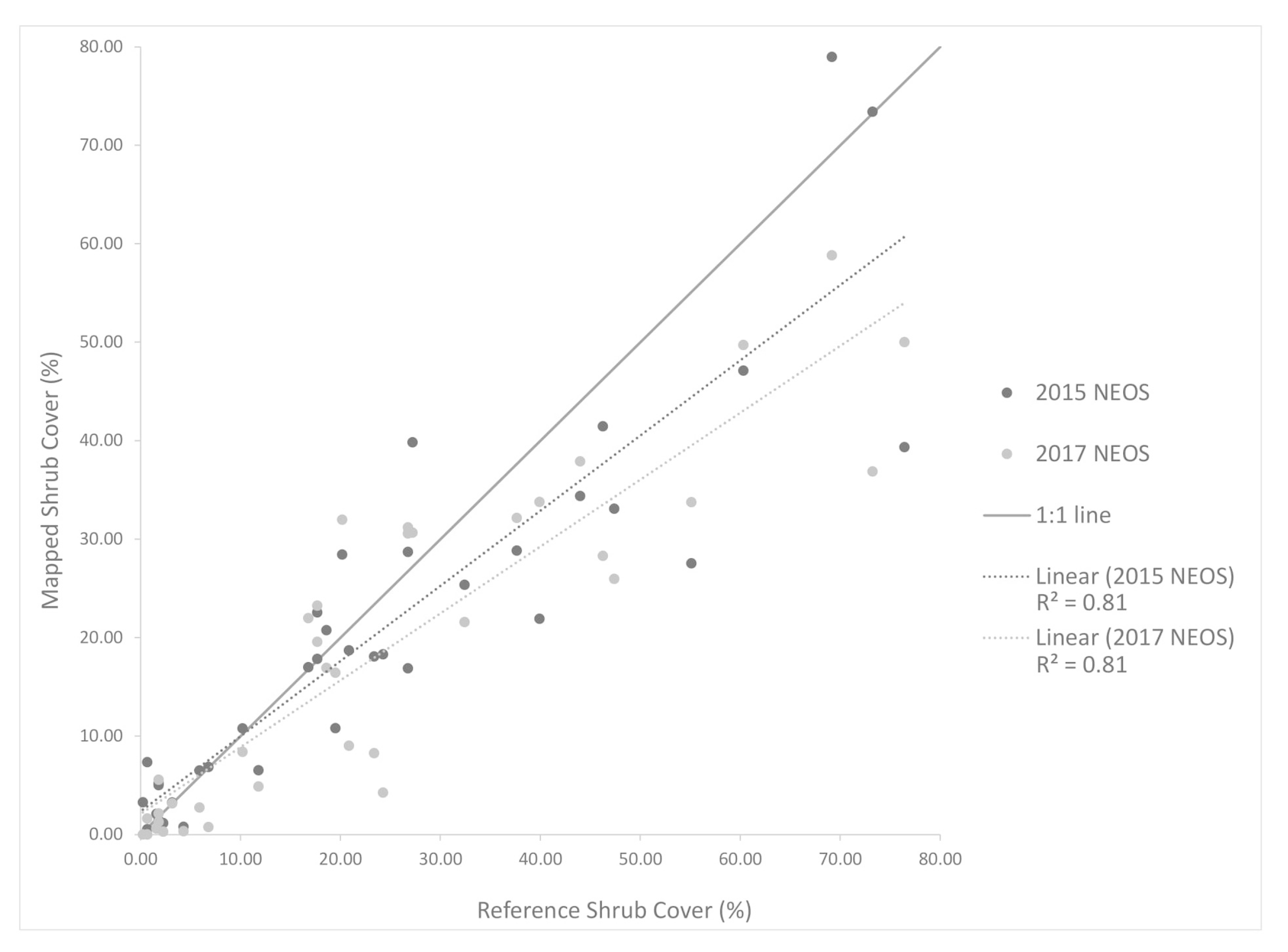
Aerial orthoimagery analyzed in this study was captured during different years and seasons, resulting in different vegetation conditions associated with vegetation phenology and available plant moisture. NAIP imagery for 2010, 2012, 2014 and 2016 were acquired in the spring and early summer, while the NEOS imagery for 2015, 2017 and 2018 were acquired in the late summer and fall. Seasonal variation in plant phenology causes differences in leaf and canopy properties between growth form types over the year and, therefore, differences in spectral reflectance signatures. During the spring growing season shrubs and herbs “green up” meaning increases in vigor, leaf area and canopy cover. As the season progresses into summer and fall, herbaceous plants senesce, while the shrubs maintain more green leaf cover. These differences in timing in image capture impact the procedures and rulesets for classifying vegetation.
Precipitation amounts for 2010 and 2012 were relatively high. Moreover, the study area receives greater precipitation during the December through March period (i.e., wet season) (
Table 1). Since 2010 and 2012 experienced greater precipitation and the imagery was captured in spring, herbs were green longer (along with shrubs) and not senescent until later than normal, which likely led to the over-classification of shrub cover. The year 2014 was an especially dry (drought) year, which may have caused some die off of shrubs or reduction in vigor and green leaf cover. In addition, the 2014 NAIP imagery was captured in the summer, whereas the 2010 and 2012 images were captured in spring. Shrub cover maps for 2014 exhibited the lowest estimated shrub cover. The 2015 and 2017 NEOS imagery was collected in fall and late summer, respectively, when more of the herbaceous cover had senesced, such that shrubs and herbs exhibited substantial differences in spectral signatures.
5.2. Image Spatial Resolution
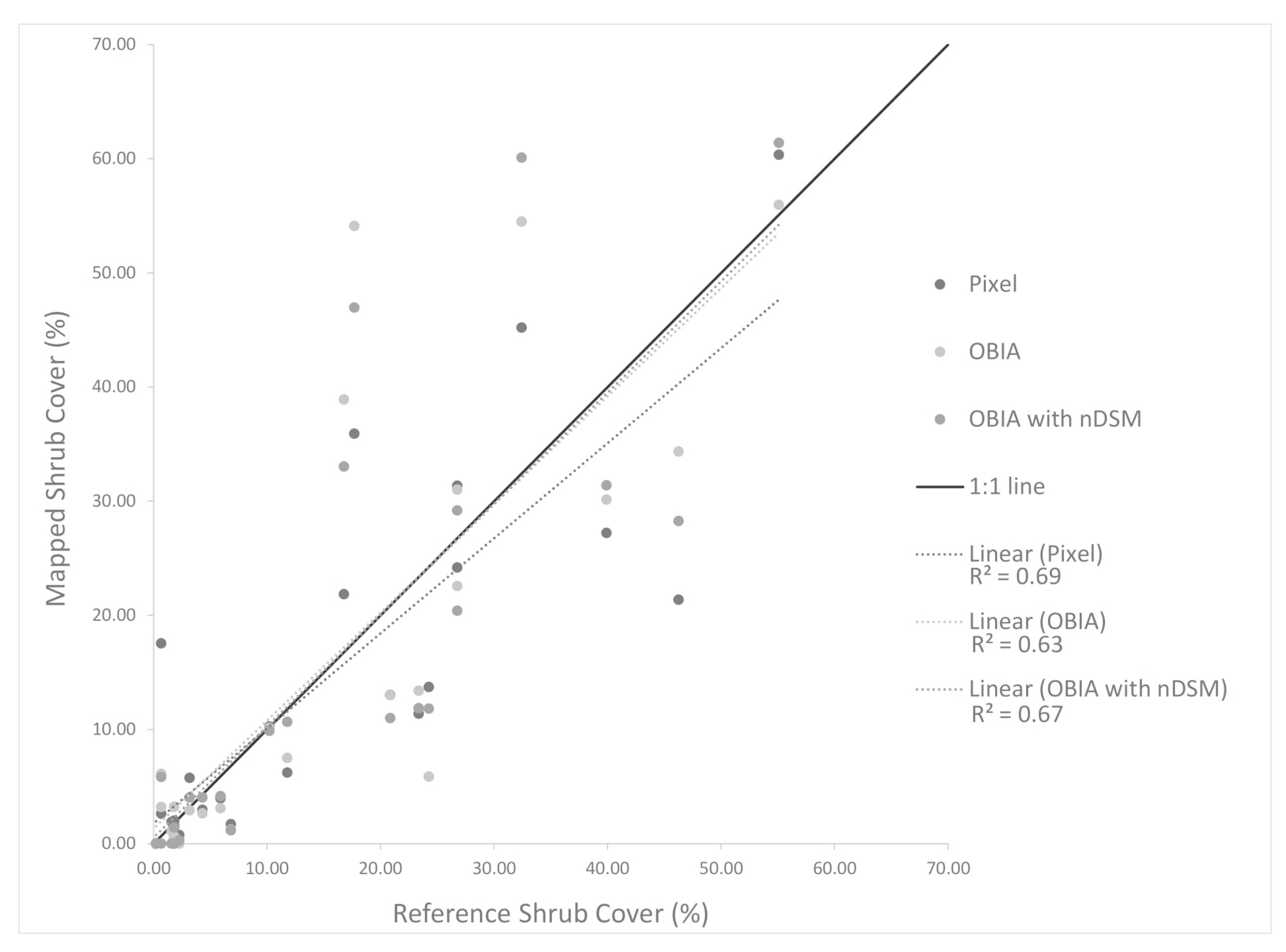
Image spatial resolution can also influence the choice of classification method chosen for mapping shrubs and the resultant accuracy of shrub cover estimates. The earlier NAIP imagery dates (2010, 2012 and 2014) have the lowest (coarsest) resolution of 1 m while that of the 2016 NAIP imagery is 0.6 m. The higher spatial resolution 2016 imagery yielded the highest accuracy shrub cover estimates of the NAIP images tested. The pixel-based approach applied to the 2016 NAIP imagery yielded more accurate estimates than for the OBIA approach for two of the three AOI, suggesting that a pixel-based approach may be a viable option for mapping shrub cover over time based on NAIP imagery. This may be useful for future applications of shrub tracking as NAIP now has set more consistent collection standards for summer image acquisitions with a 0.6 m spatial resolution.
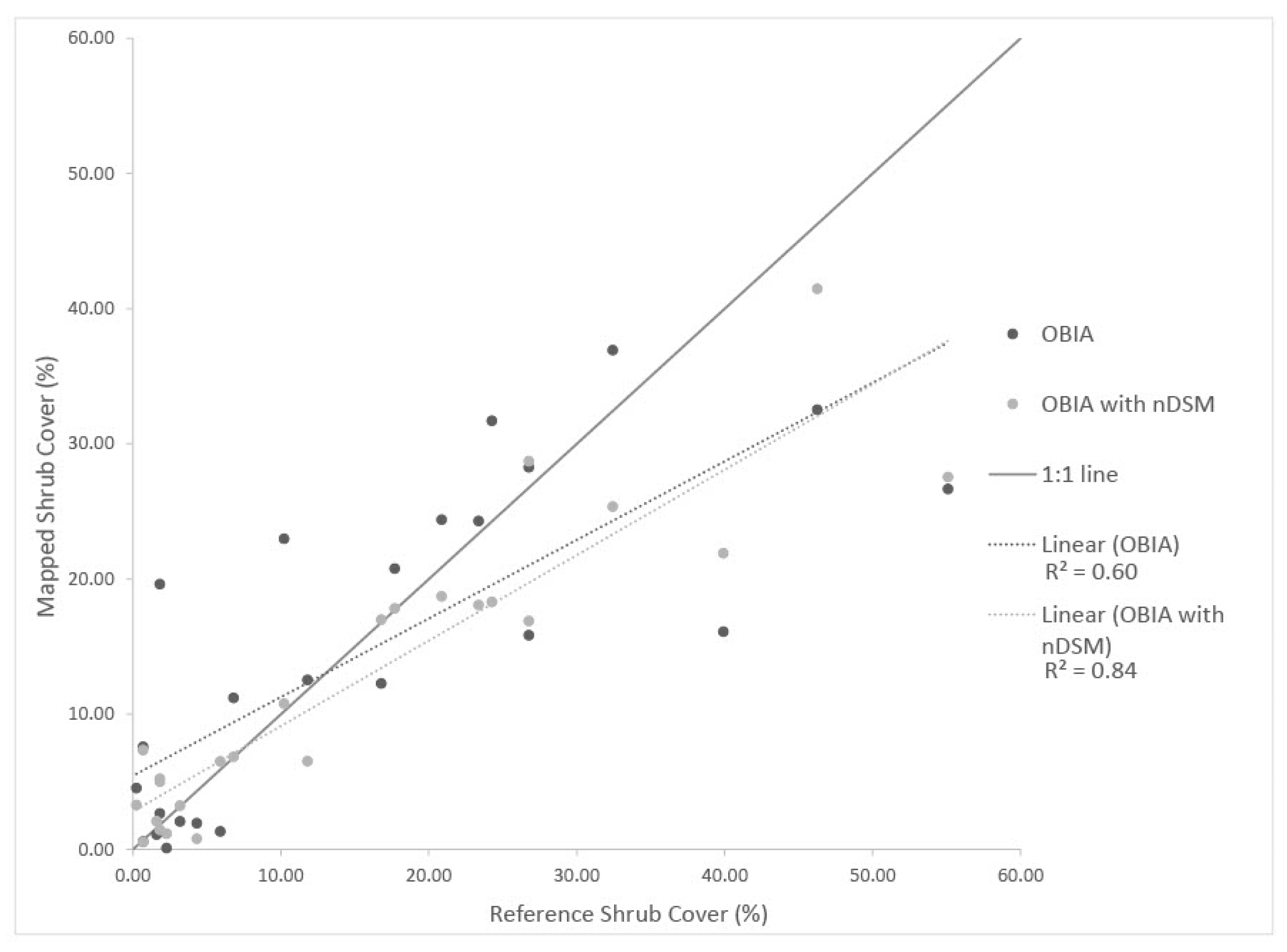
5.3. nDSM
The OBIA approach with the incorporation of nDSM allowed shrubs to be classified uniquely from the predominant non-native grasslands of San Clemente Island, based on shrub height information. With the addition of nDSM data, OBIA applied to NEOS imagery yielded the highest accuracy for mapping shrub cover. However, for AOI 3, the nDSM data were not as helpful for successfully classifying shrubs due to the lack of height signal from the predominant, low stature and densely packed shrub species, L. californicum. With minimal height information, classification was based primarily on multispectral brightness and NDVI feature inputs.
A single nDSM product, based on LiDAR data collected in 2014, was available for the study period. The single-time LiDAR-derived nDSM proved to be useful for the classification of shrubs. DSMs were generated from the highly overlapping NEOS multispectral imagery, as part of the Structure-from-Motion (SfM) processing in support of orthorectification of image frames. However, based on our cursory analyses, nDSM generated from SfM-derived DSM data were not sufficiently accurate or detailed to reliably classify shrubs vs. non-shrubs. Including a reliable nDSM derived from the same aerial image set as the orthoimage in the growth form classification process should allow changes in shrub height and expansion to be quantified, likely increasing the accuracy of shrub cover/change maps and estimates.
5.4. Fractional Cover Grid Size
Estimation of fractional vegetation cover is necessarily based on arbitrary sized and shaped spatial sampling units; for this study, square grids of three varying areal extents were tested. One criterion for determining the appropriate grid size for shrub monitoring is the sensitivity in capturing changes in shrub cover, which would tend to justify a smaller grid. For example, detection of a change in shrub cover within the 40 m grid would tend to require a greater amount of shrub cover change to yield a detectable signal, compared to the 10 m grid. However, the empirically based evidence from this study suggests that error and uncertainty in shrub cover estimates were least for the 40 m grid and greatest for the 10 m grid. A 20 m grid would seem to be a good compromise, as the error estimates for this level were only slightly greater than for the 40 m grid. Moreover, the 20 m grid is four times smaller for sensitivity purposes, and similar in size to the 25 m grid sampling scale that is advocated for monitoring growth form cover within CSS communities [
1,
7].
5.5. Challenges and Limitations
We found no sources of reference data on locations of shrubs for the earlier years (prior to 2015), and the reference data used in this study were only collected within a short period of time at the end of the study period. The reference data were generated by biologists conducting field sampling in September 2019 and through interpretation of aerial multispectral imagery captured in 2017 and 2018, having a spatial resolution twice as fine as that used for automated image classification. The 2015 and 2017 NEOS imagery resulted in more accurate classification products than the 2010 – 2016 NAIP products (
Table 3,
Table 4 and
Table 5). These results are likely due to the higher resolution of the NEOS imagery but may also be related to the fact that the field- and image-based reference data were collected near the end of the study period. However, based on the assumption that little change has occurred in the eight-year period of this study and the lack of reference data from previous years, the reference data collected in 2019 provided the basis for accuracy assessment of all shrub cover products (i.e., for all years in the study period).
The coarser spatial resolution (1 m rather than 0.15 or 0.6 m) and lack of time-specific nDSM for the earlier years made the accurate classification and cover estimation of shrubs challenging. The 1 m resolution was too coarse to accurately resolve and identify some shrub, resulting in errors and uncertainty in representing shrub cover over space and time.
Shrub cover products generated with nDSM are clearly more accurate and reliable. However, changes in the cover and height of shrubs could render a single reliable nDSM as sub-optimal for tracking changes over time (as discussed above).
5.6. Recommendations for Future Research
While the goal of this study is to create a reliable shrub cover monitoring approach, the archived image data sets that are available are not sufficient to achieve this. However, this study allows us to make recommendations for follow-on research and future shrub cover monitoring programs. As mentioned in the previous section, consistent timing for imagery capture is important to image vegetation at a similar growth state each year. Imagery collected in the late summer or early fall is optimal for distinguishing green shrubs against a background of herbaceous cover that has senesced. The incorporation of canopy height information is also crucial for the classification of the relatively sparse, low-stature shrubs on San Clemente Island and other heavily disturbed shrublands. For this study, nDSMs representing vegetation heights were derived from airborne LiDAR data, which can be expensive. Another option is using DSM created through SfM. This method is cheaper; however, generating a high-quality nDSM from SfM can be challenging from slight motions of vegetation between capture of adjacent, overlapping image frames. Very accurate and precise ground control points and image capture during relatively still wind conditions are requirements for generating reliable DSMs from SfM procedures.
OBIA classification of very-high-spatial-resolution multispectral imagery appears to be the most appropriate image analysis approach for mapping shrub growth forms and estimating their fractional cover at SCI. This study has shown that an OBIA approach that incorporates high-spatial-resolution imagery and nDSM data yields higher accuracy shrub cover maps than pixel-based mapping. Using increasingly available DSM products in combination with temporally consistent, systematically acquired high-spatial-resolution imagery, mapping changes in shrub cover should be achievable in a reliable manner. This is the emphasis of our follow-on research.
6. Conclusions
Fractional cover analysis of shrubs can provide ecologically meaningful information for the conservation and restoration of CSS communities [
7,
10]. Fractional cover of shrubs is an indication of the integrity of shrubland ecosystems. Tracking changes in shrub cover over time from a baseline may provide a monitoring tool of CSS conditions and develop a trajectory for future recovery, as long as shrub cover maps are accurate and reliable.
The major factors that influenced the reliability of shrub mapping and fractional cover estimation include the date (time of year) the imagery was collected, the spatial resolution of the imagery, the type of classifier used, the inputs to the classifier, and the grid scale used for cover estimation. We tested both pixel-based and OBIA approaches for the classification of remotely sensed imagery. Upon comparing the shrub cover between 2010 and 2018, an apparent decrease in shrub cover is evident. However, it is likely that this apparent decrease is due to over-classification of shrubs in the earlier NAIP imagery.
This study was based on both readily available and newly acquired orthoimagery with varying spatial resolutions. We found that the higher spatial resolution datasets resulted in the most accurate shrub classification and cover estimation products. We also analyzed the utility of nDSM data for classifying shrubs with OBIA. The canopy height information contained within nDSM data yielded more accurate identification of shrub objects during the classification process than when based solely on the visible/NIR image brightness values. The object-based approach with nDSM input was found to be the most accurate for identifying shrubs in all but one case (2016 NAIP). The pixel-based classifications using the 2016 NAIP imagery performed nearly as well as the OBIA methods using nDSM for AOIs 1 and 2, which include shrub landscapes with 20–60% shrub cover. These landscapes should be the focus of future ecological monitoring as they are recovering from disturbance and may show varying degrees of shrub development or transition to herbaceous cover.

 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}