
Complex Analysis of the Efficiency of Difference Reflectance Indices on the Basis of 400–700 nm Wavelengths for Revealing the Influences of Water Shortage and Heating on Plant Seedlings

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
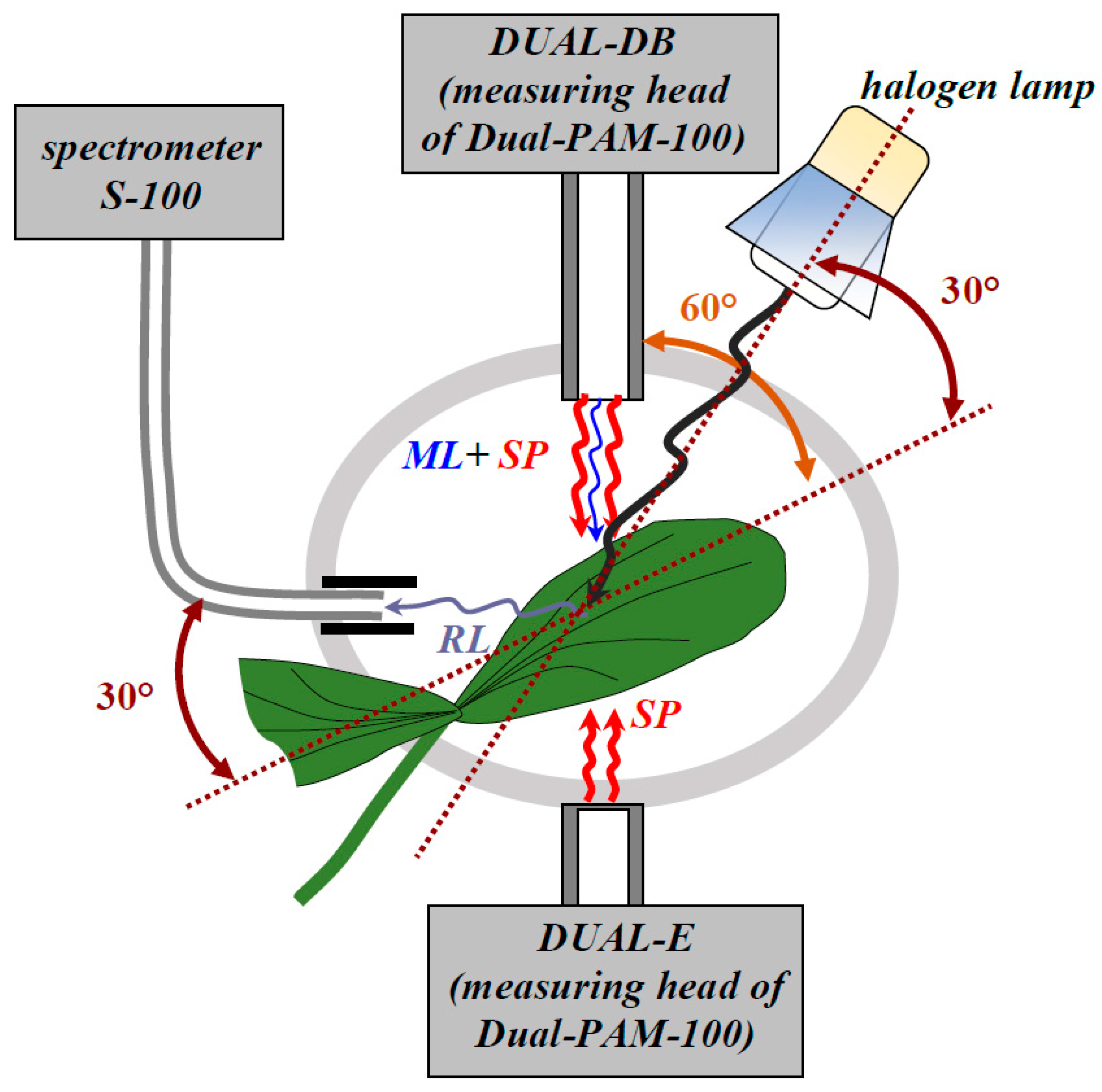
2.2. Measurement of Reflected Light and Maximal Quantum Yield of Photosystem II
2.3. Calculation of Difference Reflectance Indices and Data Analysis
- (i)
- Calculation of all possible difference RIs according to Equation (2):where I(R1) and I(R2) are intensities of reflected light at R1 and R2 wavelengths (in the spectral range from 400 to 700 nm). The intensities were averaged in a 3 nm range for increasing accuracy of the analysis. If R2 ≥ R1 then RI was not calculated. Additionally, light-induced changes in RIs (ΔRIs) were calculated according to Equation (3):where RI1 was calculated based on I(R1) and I(R2), which were measured immediately after turning on the halogen lamp (first time point), and RI2 was calculated based on I(R1) and I(R2), which were measured 7 min after turning on the halogen lamp (second time point).
- (ii)
- Calculation of significance (p) of differences between control and experimental plants for all calculated RIs (or ΔRIs); the nonparametric Mann–Whitney U test was used for estimation of p. Directions of changes were also estimated. Two-dimensional data arrays (significances and directions of changes for each RI as function of R1 and R2) were results of this stage of the analysis. The arrays were used for direct revealing the effective RIs (or ΔRIs) or for following construction of heatmaps.
- (iii)
- Construction of two-dimensional heatmaps based on the arrays. Each pixel of the heatmaps could show three variants (different colors): significant increase of the experimental RI or ΔRI (p ≤ 0.05, increase), significant decrease of the experimental RI or ΔRI (p ≤ 0.05, decrease), and absence of significant difference between control and experimental RI or ΔRI (p > 0.05).
2.4. Statistics
3. Results
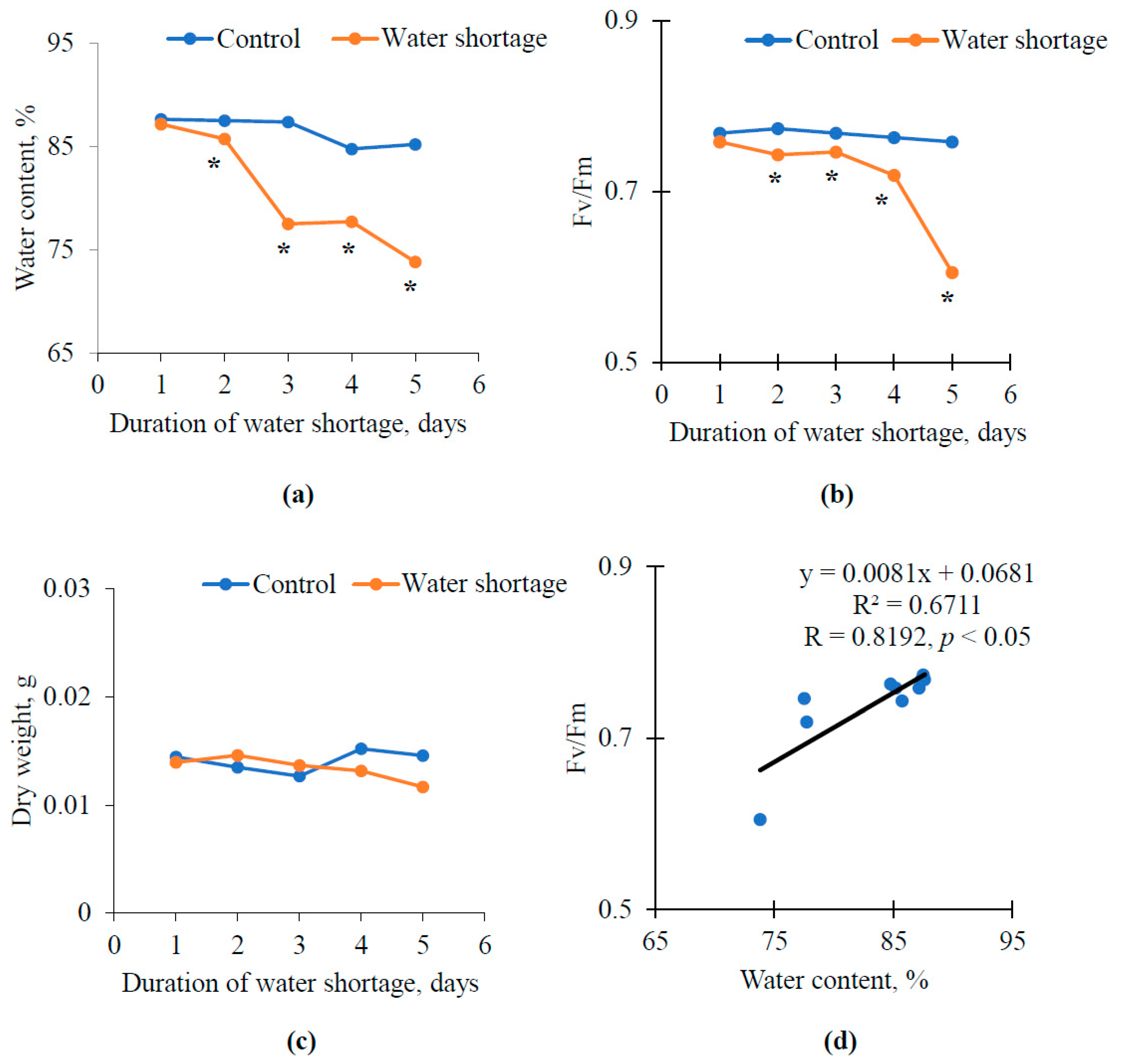
3.1. Influence of Water Shortage on Relative Water Content and Maximal Quantum Yield of Photosystem II
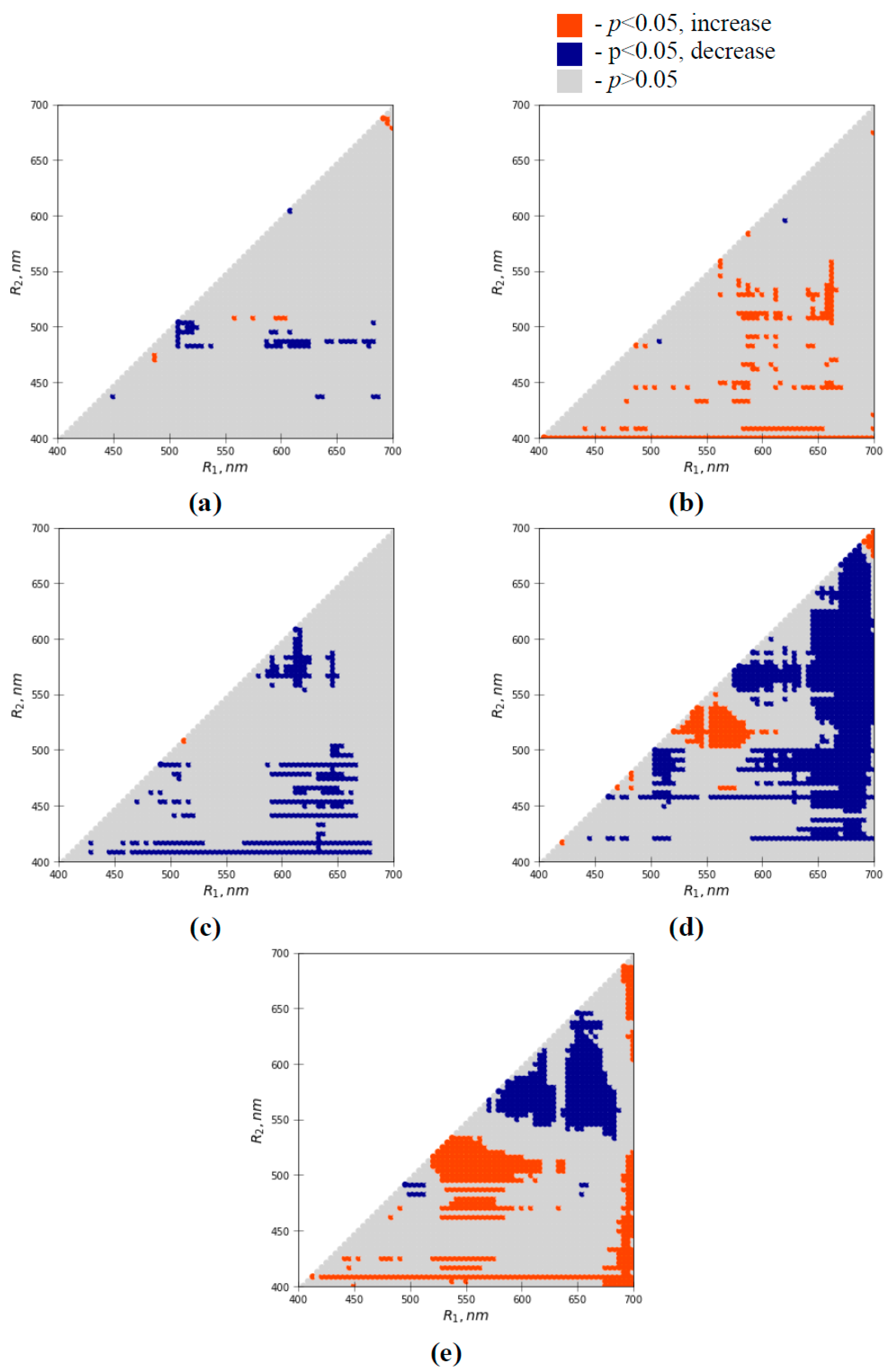
3.2. Influence of Water Shortage on Difference Reflectance Indices
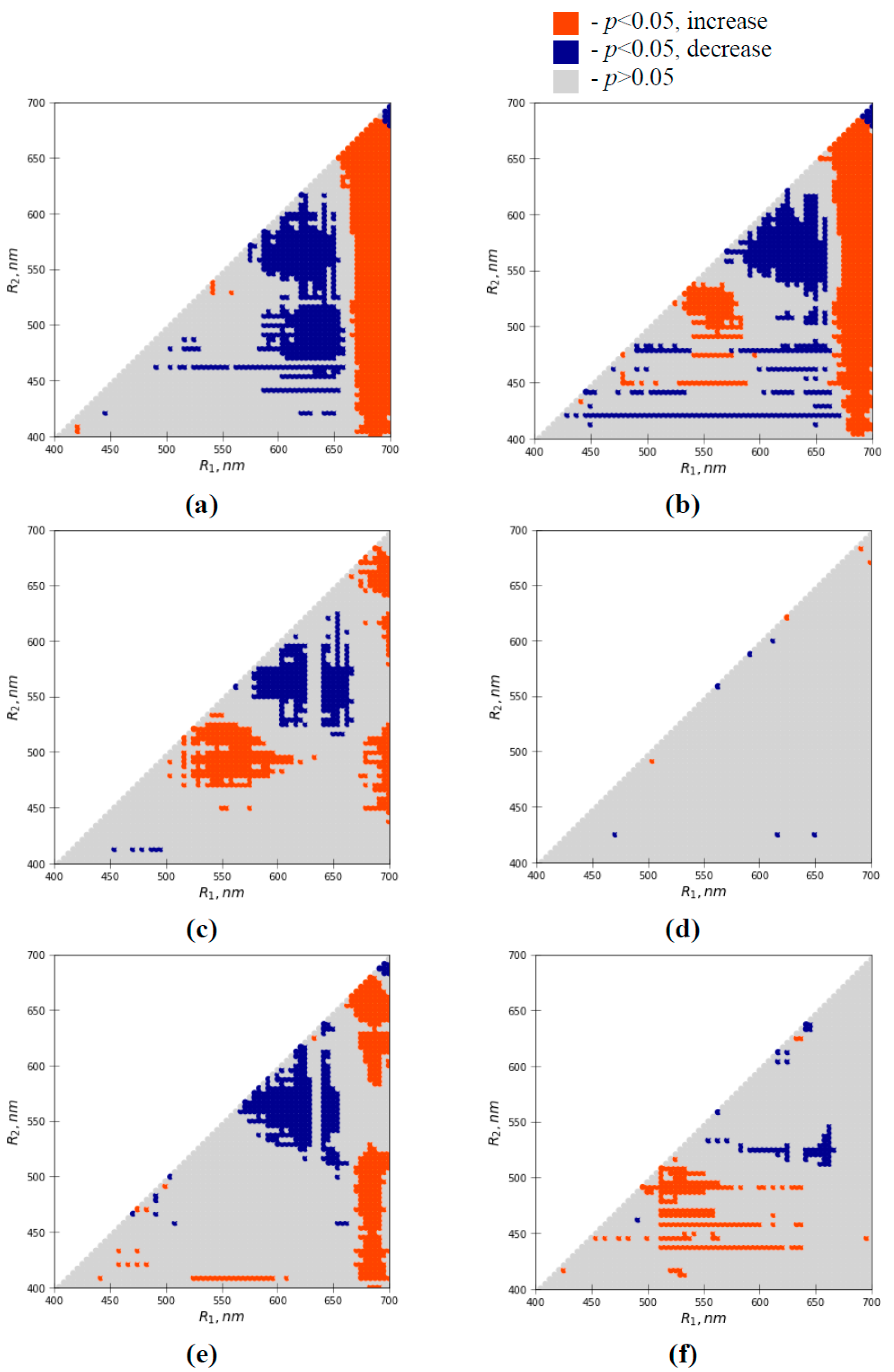
3.3. Analysis of Efficiencies of Difference Reflectance Indices for Revealing Water Shortage-Induced Changes in Seedlings
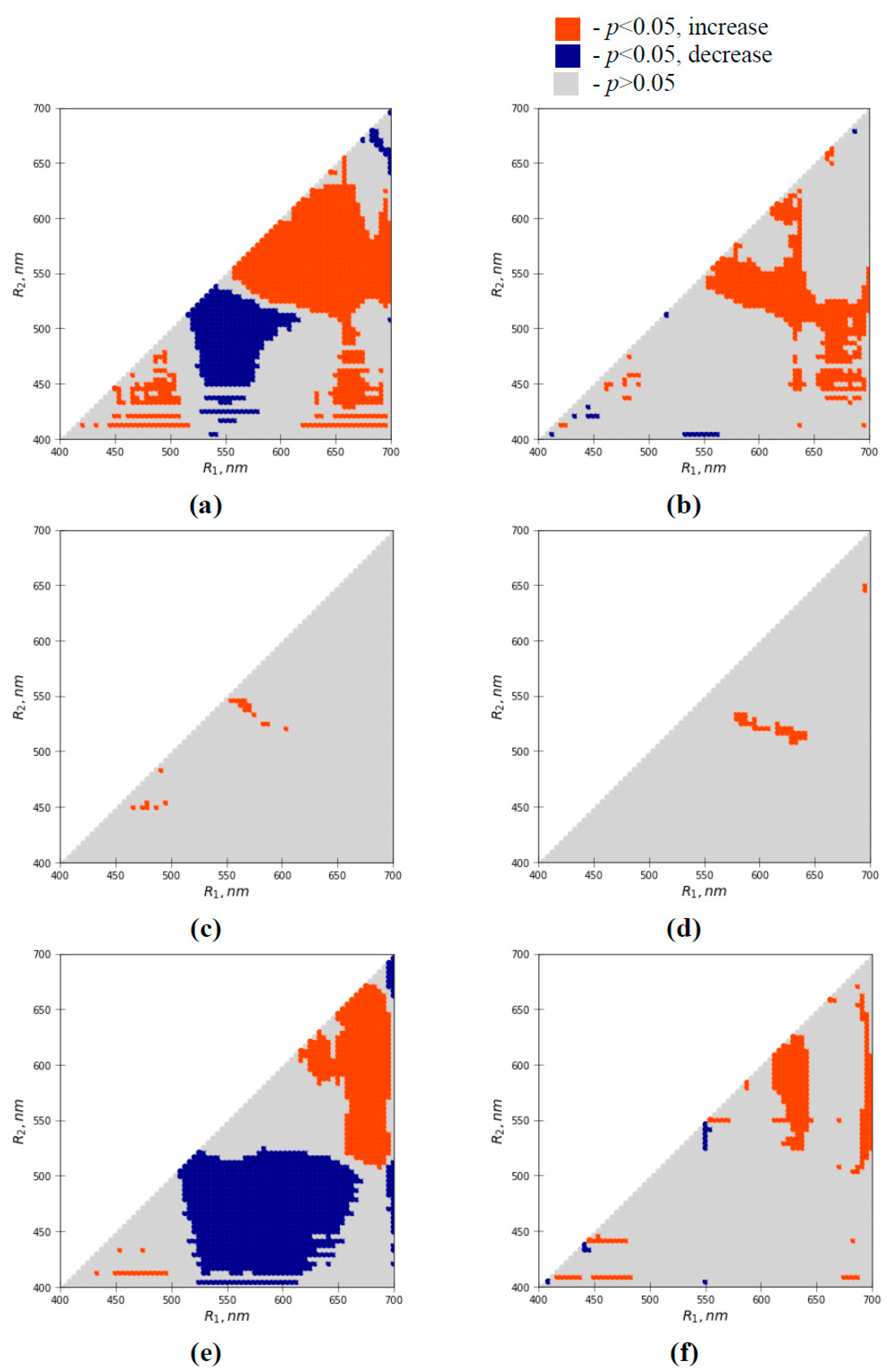
3.4. Influence of Short-Term Heating on Maximal Quantum Yield of Photosystem II and Difference Reflectance Indices
3.5. Analysis of Efficiencies of the Water Shortage-Sensitive Reflectance Indices for Revealing Heating-Induced Changes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Battisti, D.S.; Naylor, R.L. Historical warnings of future food insecurity with unprecedented seasonal heat. Science 2009, 323, 240–244. [Google Scholar] [CrossRef] [Green Version]
- Gornall, J.; Betts, R.; Burke, E.; Clark, R.; Camp, J.; Willett, K.; Wiltshire, A. Implications of climate change for agricultural productivity in the early twenty-first century. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2973–2989. [Google Scholar] [CrossRef]
- Matsubara, S. Growing plants in fluctuating environments: Why bother? J. Exp. Bot. 2018, 69, 4651–4654. [Google Scholar] [CrossRef] [PubMed]
- Zeppel, M.J.B.; Wilks, J.V.; Lewis, J.D. Impacts of extreme precipitation and seasonal changes in precipitation on plants. Biogeosciences 2014, 11, 3083–3093. [Google Scholar] [CrossRef] [Green Version]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Quiles, M.J.; López, N.I. Photoinhibition of photosystems I and II induced by exposure to high light intensity during oat plant growth. Effects on the chloroplast NADH dehydrogenase complex. Plant Sci. 2004, 166, 815–823. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Carpentier, R.; Mohanty, P. Heat stress: An overview of molecular responses in photosynthesis. Photosynth. Res. 2008, 98, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2008, 103, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Zivcak, M.; Brestic, M.; Balatova, Z.; Drevenakova, P.; Olsovska, K.; Kalaji, H.M.; Yang, X.; Allakhverdiev, S.I. Photosynthetic electron transport and specific photoprotective responses in wheat leaves under drought stress. Photosynth. Res. 2013, 117, 529–546. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Mittler, R.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Plant adaptations to the combination of drought and high temperatures. Physiol. Plant. 2018, 162, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Fromm, J.; Fei, H. Electrical signaling and gas exchange in maize plants of drying soil. Plant Sci. 1998, 132, 203–213. [Google Scholar] [CrossRef]
- Du, C.; Chai, L.; Wang, Z.; Fan, H. Response of proteome and morphological structure to short-term drought and subsequent recovery in Cucumis sativus leaves. Physiol. Plant. 2019, 167, 676–689. [Google Scholar] [CrossRef] [PubMed]
- Altuntaş, C.; Demiralay, M.; Sezgin Muslu, A.; Terzi, R. Proline-stimulated signaling primarily targets the chlorophyll degra-dation pathway and photosynthesis associated processes to cope with short-term water deficit in maize. Photosynth. Res. 2020, 144, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Pinter, P.J.J.; Hatfield, J.L.; Schepers, J.S.; Barnes, E.M.; Moran, M.S.; Daughtry, C.S.T.; Upchurch, D.R. Remote Sensing for Crop Management. Photogramm. Eng. Remote Sens. 2003, 69, 647–664. [Google Scholar] [CrossRef] [Green Version]
- Yang, C. Remote Sensing and Precision Agriculture Technologies for Crop Disease Detection and Management with a Practical Application Example. Engineering 2020, 6, 528–532. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Berjón, A.; Miller, J.R. Stress detection in crops with hyperspectral remote sensing and physical simulation models. In Proceedings of the 2004 Airborne Imaging Spectroscopy Workshop, Bruges, Belgium, 8 October 2004; pp. 1–5. [Google Scholar]
- Behmann, J.; Steinrücken, J.; Plümer, L. Detection of early plant stress responses in hyperspectral images. ISPRS J. Photogramm. Remote. Sens. 2014, 93, 98–111. [Google Scholar] [CrossRef]
- Gerhards, M.; Schlerf, M.; Mallick, K.; Udelhoven, T. Challenges and future perspectives of multi-/hyperspectral thermal in-frared remote sensing for crop water-stress detection: A review. Remote Sens. 2019, 11, 1240. [Google Scholar] [CrossRef] [Green Version]
- Mahlein, A.-K. Plant Disease Detection by Imaging Sensors—Parallels and Specific Demands for Precision Agriculture and Plant Phenotyping. Plant Dis. 2016, 100, 241–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahlein, A.K.; Kuska, M.T.; Behmann, J.; Polder, G.; Walter, A. Hyperspectral sensors and imaging technologies in phytopathology: State of the art. Annu. Rev. Phytopathol. 2018, 56, 535–558. [Google Scholar] [CrossRef]
- Prabhakar, M.; Prasad, Y.G.; Rao, M.N. Remote sensing of biotic stress in crop plants and its applications for pest management. In Crop Stress and Its Management: Perspectives and Strategies; Venkateswarlu, B., Shanker, A., Shanker, C., Maheswari, M., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 517–545. [Google Scholar]
- Gorbe, E.; Calatayud, A. Applications of chlorophyll fluorescence imaging technique in horticultural research: A review. Sci. Hortic. 2012, 138, 24–35. [Google Scholar] [CrossRef]
- Porcar-Castell, A.; Tyystjärvi, E.; Atherton, J.; Van Der Tol, C.; Flexas, J.; Pfündel, E.E.; Moreno, J.; Frankenberg, C.; Berry, J.A. Linking chlorophyll a fluorescence to photosynthesis for remote sensing applications: Mechanisms and challenges. J. Exp. Bot. 2014, 65, 4065–4095. [Google Scholar] [CrossRef]
- Zhang, C.; Atherton, J.; Peñuelas, J.; Filella, I.; Kolari, P.; Aalto, J.; Ruhanen, H.; Bäck, J.; Porcar-Castell, A. Do all chlorophyll fluorescence emission wavelengths capture the spring recovery of photosynthesis in boreal evergreen foliage? Plant Cell Environ. 2019, 42, 3264–3279. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Fu, R.; Dickinson, R.; Joiner, J.; Frankenberg, C.; Gu, L.; Xia, Y.; Fernando, N. Drought onset mechanisms revealed by satellite solar-induced chlorophyll fluorescence: Insights from two contrasting extreme events. J. Geophys. Res. Biogeosci. 2015, 120, 2427–2440. [Google Scholar] [CrossRef]
- Liu, L.; Yang, X.; Zhou, H.; Liu, S.; Zhou, L.; Li, X.; Yang, J.; Han, X.; Wu, J. Evaluating the utility of solar-induced chloro-phyll fluorescence for drought monitoring by comparison with NDVI derived from wheat canopy. Sci. Total Environ. 2018, 625, 1208–1217. [Google Scholar] [CrossRef]
- Wang, C.; Beringer, J.; Hutley, L.B.; Cleverly, J.; Li, J.; Liu, Q.; Sun, Y. Phenology Dynamics of Dryland Ecosystems Along the North Australian Tropical Transect Revealed by Satellite Solar-Induced Chlorophyll Fluorescence. Geophys. Res. Lett. 2019, 46, 5294–5302. [Google Scholar] [CrossRef]
- Jones, H.G.; Stoll, M.; Santos, T.; de Sousa, C.; Chaves, M.M.; Grant, O.M. Use of infrared thermography for monitoring sto-matal closure in the field: Application to grapevine. J. Exp. Bot. 2002, 53, 2249–2260. [Google Scholar] [CrossRef]
- Jones, H.G.; Serraj, R.; Loveys, B.R.; Xiong, L.; Wheaton, A.; Price, A.H. Thermal infrared imaging of crop canopies for the remote diagnosis and quantification of plant responses to water stress in the field. Funct. Plant Biol. 2009, 36, 978–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.-J.; Lee, B.-W. Estimation of rice growth and nitrogen nutrition status using color digital camera image analysis. Eur. J. Agron. 2013, 48, 57–65. [Google Scholar] [CrossRef]
- Gamon, J.A.; Huemmrich, K.F.; Wong, C.Y.S.; Ensminger, I.; Garrity, S.; Hollinger, D.Y.; Noormets, A.; Peñuelas, J. A re-motely sensed pigment index reveals photosynthetic phenology in evergreen conifers. Proc. Natl. Acad. Sci. USA 2016, 113, 13087–13092. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Garbulsky, M.F.; Filella, I. Photochemical reflectance index (PRI) and remote sensing of plant CO2 uptake. New Phytol. 2011, 191, 596–599. [Google Scholar] [CrossRef] [PubMed]
- Garbulsky, M.F.; Peñuelas, J.; Gamon, J.; Inoue, Y.; Filella, I. The photochemical reflectance index (PRI) and the remote sensing of leaf, canopy and ecosystem radiation use efficiencies. A review and meta-analysis. Remote Sens. Environ. 2011, 115, 281–297. [Google Scholar] [CrossRef]
- Zhang, C.; Filella, I.; Garbulsky, M.F.; Peñuelas, J. Affecting factors and recent improvements of the photochemical reflectance index (PRI) for remotely sensing foliar, canopy and ecosystemic radiation-use efficiencies. Remote Sens. 2016, 8, 677. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Filella, I.; Biel, C.; Serrano, L.; Savé, R. The reflectance at the 950–970 nm region as an indicator of plant water status. Int. J. Remote Sens. 1993, 14, 1887–1905. [Google Scholar] [CrossRef]
- Sarlikioti, V.; Driever, S.; Marcelis, L. Photochemical reflectance index as a mean of monitoring early water stress. Ann. Appl. Biol. 2010, 157, 81–89. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; González-Dugo, V.; Berni, J.A.J. Fluorescence, temperature and narrow-band indices acquired from a UAV platform for water stress detection using a micro-hyperspectral imager and a thermal camera. Remote Sens. Environ. 2012, 117, 322–337. [Google Scholar] [CrossRef]
- Hmimina, G.; Dufrêne, E.; Soudani, K. Relationship between photochemical reflectance index and leaf ecophysiological and biochemical parameters under two different water statuses: Towards a rapid and efficient correction method using real-time measurements. Plant Cell Environ. 2014, 37, 473–487. [Google Scholar] [CrossRef]
- Gerhards, M.; Schlerf, M.; Rascher, U.; Udelhoven, T.; Juszczak, R.; Alberti, G.; Miglietta, F.; Inoue, Y. Analysis of airborne optical and thermal imagery for detection ofwater stress symptoms. Remote Sens. 2018, 10, 1139. [Google Scholar] [CrossRef] [Green Version]
- Jordan, C.F. Derivation of Leaf-Area Index from Quality of Light on the Forest Floor. Ecology 1969, 50, 663–666. [Google Scholar] [CrossRef]
- Gamon, J.; Peñuelas, J.; Field, C. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote. Sens. Environ. 1992, 41, 35–44. [Google Scholar] [CrossRef]
- Penuelas, J.; Baret, F.; Filella, I. Semiempirical indices to assess carotenoids/chlorophyll a ratio from leaf spectral reflectance. Photosynthetica 1995, 31, 221–230. [Google Scholar]
- Gamon, J.A.; Surfus, J.S. Assessing leaf pigment content and activity with a refectometer. New Phytol. 1999, 143, 105–117. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Zur, Y.; Chivkunova, O.B.; Merzlyak, M.N. Assessing Carotenoid Content in Plant Leaves with Reflectance Spectroscopy. Photochem. Photobiol. 2007, 75, 272–281. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Chivkunova, O.B.; Merzlyak, M.N. Nondestructive estimation of anthocyanins and chlorophylls in anthocyanic leaves. Am. J. Bot. 2009, 96, 1861–1868. [Google Scholar] [CrossRef]
- Peñuelas, J.; Gamon, J.; Fredeen, A.; Merino, J.; Field, C. Reflectance indices associated with physiological changes in nitrogen- and water-limited sunflower leaves. Remote Sens. Environ. 1994, 48, 135–146. [Google Scholar] [CrossRef]
- Gamon, J.A.; Serrano, L.; Surfus, J.S. The photochemical reflectance index: An optical indicator of photosynthetic radiation use efficiency across species, functional types, and nutrient levels. Oecologia 1997, 112, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Brueck, H.; Asch, F. Chlorophyll index, photochemical reflectance index and chlorophyll fluorescence measurements of rice leaves supplied with different N levels. J. Photochem. Photobiol. B Biol. 2012, 113, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Marino, G.; Llusia, J.; Morfopoulos, C.; Farré-Armengol, G.; Filella, I. Photochemical reflectance index as an indirect estimator of foliar isoprenoid emissions at the ecosystem level. Nat. Commun. 2013, 4, 2604. [Google Scholar] [CrossRef] [Green Version]
- Balzarolo, M.; Peñuelas, J.; Filella, I.; Portillo-Estrada, M.; Ceulemans, R. Assessing ecosystem isoprene emissions by hyperspectral remote sensing. Remote Sens. 2018, 10, 1086. [Google Scholar] [CrossRef] [Green Version]
- Sukhov, V.; Sukhova, E.; Gromova, E.; Surova, L.; Nerush, V.; Vodeneev, V. The electrical signal-induced systemic photosynthetic response is accompanied by changes in the photochemical reflectance index in pea. Funct. Plant Biol. 2019, 46, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Sukhova, E.; Yudina, L.; Akinchits, E.; Vodeneev, V.; Sukhov, V. Influence of electrical signals on pea leaf reflectance in the 400–800-nm range. Plant Signal. Behav. 2019, 14, 1610301. [Google Scholar] [CrossRef]
- Sukhova, E.; Yudina, L.; Gromova, E.; Nerush, V.; Vodeneev, V.; Sukhov, V. Burning-induced electrical signals influence broadband reflectance indices and water index in pea leaves. Plant Signal. Behav. 2020, 15, 1737786. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, M.; Norton, R.; Thorp, K.R.; Wang, G. Association of Spectral Reflectance Indices with Plant Growth and Lint Yield in Upland Cotton. Crop. Sci. 2012, 52, 849–857. [Google Scholar] [CrossRef]
- Brandão, Z.N.; Sofiatti, V.; Bezerra, J.R.C.; Ferreira, G.B.; Medeiros, J.C. Spectral reflectance for growth and yield assessment of irrigated cotton. AJCS 2015, 9, 75–84. [Google Scholar]
- Evain, S.; Flexas, J.; Moya, I. A new instrument for passive remote sensing: 2. Measurement of leaf and canopy reflectance changes at 531 nm and their relationship with photosynthesis and chlorophyll fluorescence. Remote Sens. Environ. 2004, 91, 175–185. [Google Scholar] [CrossRef]
- Filella, I.; Porcar-Castell, A.; Munné-Bosch, S.; Bäck, J.; Garbulsky, M.F.; Peñuelas, J. PRI assessment of long-term changes in carotenoids/chlorophyll ratio and short-term changes in de-epoxidation state of the xanthophyll cycle. Int. J. Remote Sens. 2009, 30, 4443–4455. [Google Scholar] [CrossRef]
- Sukhova, E.; Sukhov, V. Relation of Photochemical Reflectance Indices Based on Different Wavelengths to the Parameters of Light Reactions in Photosystems I and II in Pea Plants. Remote Sens. 2020, 12, 1312. [Google Scholar] [CrossRef] [Green Version]
- Mahlein, A.K.; Steiner, U.; Dehne, H.W.; Oerke, E.C. Spectral signatures of sugar beet leaves for the detection and differentiation of diseases. Precis. Agric. 2010, 11, 413–431. [Google Scholar] [CrossRef]
- Zhu, H.; Chu, B.; Zhang, C.; Liu, F.; Jiang, L.; He, Y. Hyperspectral Imaging for Presymptomatic Detection of Tobacco Disease with Successive Projections Algorithm and Machine-learning Classifiers. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Fu, P.; Meacham-Hensold, K.; Guan, K.; Bernacchi, C.J. Hyperspectral Leaf Reflectance as Proxy for Photosynthetic Capacities: An Ensemble Approach Based on Multiple Machine Learning Algorithms. Front. Plant Sci. 2019, 10, 730. [Google Scholar] [CrossRef]
- Gold, K.M.; Townsend, P.A.; Herrmann, I.; Gevens, A.J. Investigating potato late blight physiological differences across potato cultivars with spectroscopy and machine learning. Plant Sci. 2020, 295, 110316. [Google Scholar] [CrossRef] [PubMed]
- Sonobe, R.; Hirono, Y.; Oi, A. Non-Destructive Detection of Tea Leaf Chlorophyll Content Using Hyperspectral Reflectance and Machine Learning Algorithms. Plants 2020, 9, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.K.; Ganapathysubramanian, B.; Sarkar, S.; Singh, A. Deep learning for plant stress phenotyping: Trends and future perspectives. Trends Plant Sci. 2018, 23, 883–898. [Google Scholar] [CrossRef] [Green Version]
- Paulus, S.; Mahlein, A.-K. Technical workflows for hyperspectral plant image assessment and processing on the greenhouse and laboratory scale. GigaScience 2020, 9, 90. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Baret, F. PROSPECT: A model of leaf optical properties spectra. Remote Sens. Environ. 1990, 34, 75–91. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, J.; Wang, F.; Blackburn, G.A.; Zhang, H.K.; Wang, X.; Wei, C.; Zhang, K.; Wei, C. An extended PRO-SPECT: Advance in the leaf optical properties model separating total chlorophylls into chlorophyll a and b. Sci. Rep. 2017, 7, 6429. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Comar, A.; Burger, P.; Bancal, P.; Weiss, M.; Baret, F. Estimation of leaf traits from reflectance measurements: Comparison between methods based on vegetation indices and several versions of the PROSPECT model. Plant Methods 2018, 14, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ustin, S.L.; Jacquemoud, S.; Govaerts, Y. Simulation of photon transport in a three-dimensional leaf: Implications for photo-synthesis. Plant Cell Environ. 2001, 24, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Hikosaka, K.; Kumagai, T.; Ito, A. Modeling canopy photosynthesis. In Canopy Photosynthesis: From Basics to Applications. Advances in Photosynthesis and Respiration (Including Bioenergy and Related Processes); Hikosaka, K., Niinemets, Ü., Anten, N., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 239–268. [Google Scholar]
- Evers, J.B.; Vos, J.; Yin, X.; Romero, P.; Van Der Putten, P.E.L.; Struik, P.C. Simulation of wheat growth and development based on organ-level photosynthesis and assimilate allocation. J. Exp. Bot. 2010, 61, 2203–2216. [Google Scholar] [CrossRef] [Green Version]
- Hao, D.; Wen, J.; Xiao, Q.; Wu, S.; Lin, X.; You, D.; Tang, Y. Modeling Anisotropic Reflectance Over Composite Sloping Terrain. IEEE Trans. Geosci. Remote Sens. 2018, 56, 3903–3923. [Google Scholar] [CrossRef]
- Rouse, J.W., Jr.; Haas, R.H.; Schell, J.A.; Deering, D.W.; Harlan, J.C. Monitoring the Vernal Advancement and Retrogradation (Green Wave Effect) of Natural Vegetation; Type III Final Rep; The National Aeronautics and Space Administration (NASA)/Goddard Space Flight Center (GSFC): Greenbelt, MD, USA, 1974.
- Eitel, J.U.H.; Long, D.S.; Gessler, P.E.; Hunt, E.R., Jr.; Brown, D.J. Sensitivity of ground-based remote sensing estimates of wheat chlorophyll content to variation in soil reflectance. Soil Sci. Soc. Am. J. 2009, 73, 1715–1723. [Google Scholar] [CrossRef] [Green Version]
- Penuelas, J.P.J.; Pinol, J.; Ogaya, R.; Filella, I. Estimation of plant water concentration by the reflectance Water Index WI (R900/R970). Int. J. Remote Sens. 1997, 18, 2869–2875. [Google Scholar] [CrossRef]
- Gao, B.-C. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Sukhova, E.; Sukhov, V. Connection of the Photochemical Reflectance Index (PRI) with the Photosystem II Quantum Yield and Nonphotochemical Quenching Can Be Dependent on Variations of Photosynthetic Parameters among Investigated Plants: A Meta-Analysis. Remote Sens. 2018, 10, 771. [Google Scholar] [CrossRef] [Green Version]
- Badgley, G.; Field, C.B.; Berry, J.A. Canopy near-infrared reflectance and terrestrial photosynthesis. Sci. Adv. 2017, 3, e1602244. [Google Scholar] [CrossRef] [Green Version]
- Sukhova, E.M.; Yudina, L.M.; Vodeneev, V.A.; Sukhov, V.S. Analysis of Changes in Photochemical Reflectance Index (PRI) in Relation to the Acidification of the Lumen of the Chloroplasts of Pea and Geranium Leaves under a Short-Term Illumination. Biochem. Suppl. Ser. A Membr. Cell Biol. 2019, 13, 243–252. [Google Scholar] [CrossRef]
- Ali, S.; Henchiri, M.; Yao, F.; Zhang, J. Analysis of vegetation dynamics, drought in relation with climate over South Asia from 1990 to 2011. Environ. Sci. Pollut. Res. 2019, 26, 11470–11481. [Google Scholar] [CrossRef]
- Zhang, F.; Zhou, G. Estimation of vegetation water content using hyperspectral vegetation indices: A comparison of crop water indicators in response to water stress treatments for summer maize. BMC Ecol. 2019, 19, 18. [Google Scholar] [CrossRef] [Green Version]
- Caturegli, L.; Matteoli, S.; Gaetani, M.; Grossi, N.; Magni, S.; Minelli, A.; Corsini, G.; Remorini, D.; Volterrani, M. Effects of water stress on spectral reflectance of bermudagrass. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, X.; Goatley, M.; Ervin, E. Heat Shock Proteins in Relation to Heat Stress Tolerance of Creeping Bentgrass at Different N Levels. PLoS ONE 2014, 9, e102914. [Google Scholar] [CrossRef] [Green Version]
- Sytar, O.; Brücková, K.; Kovár, M.; Živčák, M.; Hemmerich, I.; Brestič, M. Nondestructive detection and biochemical quantification of buckwheat leaves using visible (VIS) and near-infrared (NIR) hyperspectral reflectance imaging. J. Centr. Eur. Agric. 2017, 18, 864–878. [Google Scholar] [CrossRef] [Green Version]
- Kovar, M.; Brestic, M.; Sytar, O.; Barek, V.; Hauptvogel, P.; Zivcak, M. Evaluation of Hyperspectral Reflectance Parameters to Assess the Leaf Water Content in Soybean. Water 2019, 11, 443. [Google Scholar] [CrossRef] [Green Version]
- El-Hendawy, S.; Al-Suhaibani, N.; Dewir, Y.H.; Elsayed, S.; Alotaibi, M.; Hassan, W.; Refay, Y.; Tahir, M.U. Ability of modified spectral reflectance indices for estimating growth and photosynthetic efficiency of wheat under saline field conditions. Agronomy 2019, 9, 35. [Google Scholar] [CrossRef] [Green Version]
- El-Hendawy, S.E.; Alotaibi, M.; Al-Suhaibani, N.; Al-Gaadi, K.; Hassan, W.; Dewir, Y.H.; Emam, M.A.E.-G.; Elsayed, S.; Schmidhalter, U. Comparative Performance of Spectral Reflectance Indices and Multivariate Modeling for Assessing Agronomic Parameters in Advanced Spring Wheat Lines Under Two Contrasting Irrigation Regimes. Front. Plant Sci. 2019, 10, 1537. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Feng, M.; Xiao, L.; Yang, W.; Wang, C.; Jia, X.; Zhao, Y.; Zhao, C.; Muhammad, S.K.; Li, D. Assessment of plant water status in winter wheat (Triticum aestivum L.) based on canopy spectral indices. PLoS ONE 2019, 14, e0216890. [Google Scholar] [CrossRef]
- Balegh, S.E.; Biddulph, O. The Photosynthetic Action Spectrum of the Bean Plant. Plant Physiol. 1970, 46, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Inada, K. Action spectra for photosynthesis in higher plants. Plant Cell Physiol. 1976, 17, 355–365. [Google Scholar] [CrossRef]
- Yudina, L.; Sukhova, E.; Gromova, E.; Nerush, V.; Vodeneev, V.; Sukhov, V. A light-induced decrease in the photochemical reflectance index (PRI) can be used to estimate the energy-dependent component of non-photochemical quenching under heat stress and soil drought in pea, wheat, and pumpkin. Photosynth. Res. 2020, 146, 175–187. [Google Scholar] [CrossRef]
- Sukhov, V.S.; Gromova, E.N.; Sukhova, E.M.; Surova, L.M.; Nerush, V.N.; Vodeneev, V.A. Analysis of Correlations between the Indexes of Light-Dependent Reactions of Photosynthesis and the Photochemical Reflectance Index (PRI) in Pea Leaves under Short-Term Illumination. Biochem. Suppl. Ser. A Membr. Cell Biol. 2019, 13, 67–77. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dąbrowski, P.; et al. Frequently asked questions about in vivo chlorophyll fluorescence: Practical issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef] [Green Version]
- Whitmarsh, J. The Photosynthetic Process. In Concepts in Photobiology; Singhal, G.S., Renger, G., Sopory, S.K., Irrgang, K.D., Eds.; Springer: Dordrecht, The Netherlands, 1999; pp. 11–51. [Google Scholar]
- Sukhova, E.; Sukhov, V. Analysis of Light-Induced Changes in the Photochemical Reflectance Index (PRI) in Leaves of Pea, Wheat, and Pumpkin Using Pulses of Green-Yellow Measuring Light. Remote Sens. 2019, 11, 810. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Wahbi, S.; Tsonev, T.; Haworth, M.; Liu, S.; Centritto, M. On the Use of Leaf Spectral Indices to Assess Water Status and Photosynthetic Limitations in Olea europaea L. during Water-Stress and Recovery. PLoS ONE 2014, 9, e105165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayat, B.; Van Der Tol, C.; Verhoef, W. Remote Sensing of Grass Response to Drought Stress Using Spectroscopic Techniques and Canopy Reflectance Model Inversion. Remote Sens. 2016, 8, 557. [Google Scholar] [CrossRef] [Green Version]
- Sukhova, E.; Khlopkov, A.; Vodeneev, V.; Sukhov, V. Simulation of a nonphotochemical quenching in plant leaf under different light intensities. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148138. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Li, X.-P.; Niyogi, K.K. Non-Photochemical Quenching. A Response to Excess Light Energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruban, A.V. Nonphotochemical Chlorophyll Fluorescence Quenching: Mechanism and Effectiveness in Protecting Plants from Photodamage. Plant Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demmig-Adams, B. Carotenoids and photoprotection in plants: A role for the xanthophyll zeaxanthin. Biochim. Biophys. Acta (BBA) Bioenerg. 1990, 1020, 1–24. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Title 1 | 1–5 Days | 2–5 Days | 3–5 Days | 4–5 Days | 5 Days |
|---|---|---|---|---|---|
| RIs | 0.1903% | 1.7504% | 22.8691% | 51.0274% | 71.1568% |
| ΔRIs | 0% | 0% | 1.5221% | 15.6012% | 31.3166% |
| Index | R1, nm | R2, nm | Time Range of RI Efficiency (p < 0.05) | Correlation Coefficient between RI and Fv/Fm | Correlation Coefficient between RI and Water Content | Direction of Changes in RI |
|---|---|---|---|---|---|---|
| NDVI | 764–796 (780) | 664–677 (670) | 3–5 days | 0.9513 * | 0.8931 * | decrease |
| WI | 900 | 970 | 5 day | 0.4052 | 0.6318 | decrease |
| RI 1 | 621 | 442 | 1–5 days | −0.7435 * | −0.8618 * | increase |
| RI 2 | 629 | 442 | 1–5 days | −0.7606 * | −0.8814 * | increase |
| RI 3 | 633 | 442 | 1–5 days | −0.7539 * | −0.8576 * | increase |
| RI 4 | 637 | 442 | 1–5 days | −0.7636 * | −0.8654 * | increase |
| RI 5 | 700 | 442 | 1–5 days | −0.7223 * | −0.8281 * | increase |
| RI 6 | 475 | 450 | 2–5 days | −0.6312 | −0.8849 * | increase |
| RI 7 | 487 | 421 | 2–5 days | −0.8396 * | −0.8985 * | increase |
| RI 8 | 487 | 458 | 2–5 days | −0.7208 * | −0.8039 * | increase |
| RI 9 | 491 | 421 | 2–5 days | −0.8666 * | −0.8983 * | increase |
| RI 10 | 496 | 421 | 2–5 days | −0.8326 * | −0.9099 * | increase |
| RI 11 | 496 | 479 | 2–5 days | −0.6760 * | −0.8831 * | increase |
| RI 12 | 496 | 483 | 2–5 days | −0.6754 * | −0.8997 * | increase |
| RI 13 | 500 | 421 | 2–5 days | −0.8300 * | −0.9272 * | increase |
| RI 14 | 500 | 450 | 2–5 days | −0.7071 * | −0.9244 * | increase |
| RI 15 | 500 | 471 | 2–5 days | −0.8178 * | −0.9721 * | increase |
| RI 16 | 500 | 479 | 2–5 days | −0.6811 * | −0.9096 * | increase |
| RI 17 | 500 | 483 | 2–5 days | −0.7159 * | −0.9491 * | increase |
| RI 18 | 504 | 421 | 2–5 days | −0.8219 * | −0.9346 * | increase |
| RI 19 | 504 | 450 | 2–5 days | −0.7061 * | −0.9328 * | increase |
| RI 20 | 504 | 471 | 2–5 days | −0.7963 * | −0.9404 * | increase |
| RI 21 | 504 | 479 | 2–5 days | −0.7075 * | −0.9309 * | increase |
| RI 22 | 508 | 421 | 2–5 days | −0.7957 * | −0.9308 * | increase |
| RI 23 | 512 | 421 | 2–5 days | −0.7642 * | −0.9077 * | increase |
| RI 24 | 613 | 604 | 2–5 days | −0.9252 * | −0.9184 * | increase |
| RI 25 | 629 | 421 | 2–5 days | −0.7970 * | −0.8835 * | increase |
| RI 26 | 633 | 421 | 2–5 days | −0.7998 * | −0.8851 * | increase |
| RI 27 | 637 | 421 | 2–5 days | −0.7968 * | −0.8817 * | increase |
| RI 28 | 654 | 421 | 2–5 days | −0.8061 * | −0.9084 * | increase |
| RI 29 | 654 | 442 | 2–5 days | −0.8077 * | −0.9210 * | increase |
| RI 30 | 658 | 421 | 2–5 days | −0.8101 * | −0.9074 * | increase |
| RI 31 | 658 | 442 | 2–5 days | −0.8099 * | −0.9202 * | increase |
| RI 32 | 662 | 421 | 2–5 days | −0.8173 * | −0.9121 * | increase |
| RI 33 | 662 | 442 | 2–5 days | −0.8173 * | −0.9213 * | increase |
| RI 34 | 667 | 421 | 2–5 days | −0.8183 * | −0.9063 * | increase |
| RI 35 | 667 | 442 | 2–5 days | −0.8166 * | −0.9135 * | increase |
| RI 36 | 671 | 421 | 2–5 days | −0.8153 * | −0.9033 * | increase |
| RI 37 | 671 | 433 | 2–5 days | −0.8441 * | −0.8884 * | increase |
| RI 38 | 675 | 421 | 2–5 days | −0.8237 * | −0.9037 * | increase |
| RI 39 | 679 | 421 | 2–5 days | −0.8288 * | −0.9027 * | increase |
| RI 40 | 683 | 421 | 2–5 days | −0.8317 * | −0.8973 * | increase |
| RI 41 | 687 | 421 | 2–5 days | −0.8399 * | −0.9041 * | increase |
| RI 42 | 687 | 433 | 2–5 days | −0.8822 * | −0.8702 * | increase |
| RI 43 | 692 | 421 | 2–5 days | −0.8431 * | −0.9067 * | increase |
| RI 44 | 692 | 442 | 2–5 days | −0.8250 * | −0.9145 * | increase |
| RI 45 | 696 | 421 | 2–5 days | −0.8414 * | −0.9190 * | increase |
| RI 46 | 696 | 442 | 2–5 days | −0.8020 * | −0.9117 * | increase |
| Index | 1 h | 1 Day | Index | 1 h | 1 Day |
|---|---|---|---|---|---|
| NDVI | Decrease | - | RI(658; 421) | Increase | - |
| WI | - | - | RI(658; 442) | Increase | - |
| RI(487; 421) | Increase | - | RI(662; 421) | Increase | - |
| RI(491; 421) | Increase | - | RI(662; 442) | Increase | - |
| RI(496; 421) | Increase | - | RI(667; 421) | Increase | - |
| RI(496; 479) | Increase | - | RI(667; 442) | Increase | - |
| RI(500; 421) | Increase | - | RI(671; 421) | Increase | - |
| RI(500; 450) | Increase | - | RI(671; 433) | Increase | - |
| RI(504; 421) | Increase | - | RI(675; 421) | Increase | - |
| RI(504; 450) | Increase | - | RI(679; 421) | Increase | - |
| RI(508; 421) | Increase | - | RI(683; 421) | Increase | - |
| RI(613; 604) | Increase | Increase | RI(687; 421) | Increase | - |
| RI(633; 421) | Increase | - | RI(687; 433) | Increase | - |
| RI(633; 442) | Increase | - | RI(692; 421) | Increase | - |
| RI(637; 421) | Increase | - | RI(692; 442) | Increase | - |
| RI(654; 421) | Increase | - | RI(696; 421) | Increase | - |
| RI(654; 442) | Increase | - | RI(487; 458) | - | Increase |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sukhova, E.; Yudina, L.; Gromova, E.; Ryabkova, A.; Kior, D.; Sukhov, V. Complex Analysis of the Efficiency of Difference Reflectance Indices on the Basis of 400–700 nm Wavelengths for Revealing the Influences of Water Shortage and Heating on Plant Seedlings. Remote Sens. 2021, 13, 962. https://doi.org/10.3390/rs13050962
Sukhova E, Yudina L, Gromova E, Ryabkova A, Kior D, Sukhov V. Complex Analysis of the Efficiency of Difference Reflectance Indices on the Basis of 400–700 nm Wavelengths for Revealing the Influences of Water Shortage and Heating on Plant Seedlings. Remote Sensing. 2021; 13(5):962. https://doi.org/10.3390/rs13050962
Chicago/Turabian StyleSukhova, Ekaterina, Lyubov Yudina, Ekaterina Gromova, Anastasiia Ryabkova, Dmitry Kior, and Vladimir Sukhov. 2021. "Complex Analysis of the Efficiency of Difference Reflectance Indices on the Basis of 400–700 nm Wavelengths for Revealing the Influences of Water Shortage and Heating on Plant Seedlings" Remote Sensing 13, no. 5: 962. https://doi.org/10.3390/rs13050962
APA StyleSukhova, E., Yudina, L., Gromova, E., Ryabkova, A., Kior, D., & Sukhov, V. (2021). Complex Analysis of the Efficiency of Difference Reflectance Indices on the Basis of 400–700 nm Wavelengths for Revealing the Influences of Water Shortage and Heating on Plant Seedlings. Remote Sensing, 13(5), 962. https://doi.org/10.3390/rs13050962