
Phytoplankton Phenology in the Coastal Zone of Cyprus, Based on Remote Sensing and In Situ Observations

 , ,
, , 
 , and
, and
Abstract
:1. Introduction
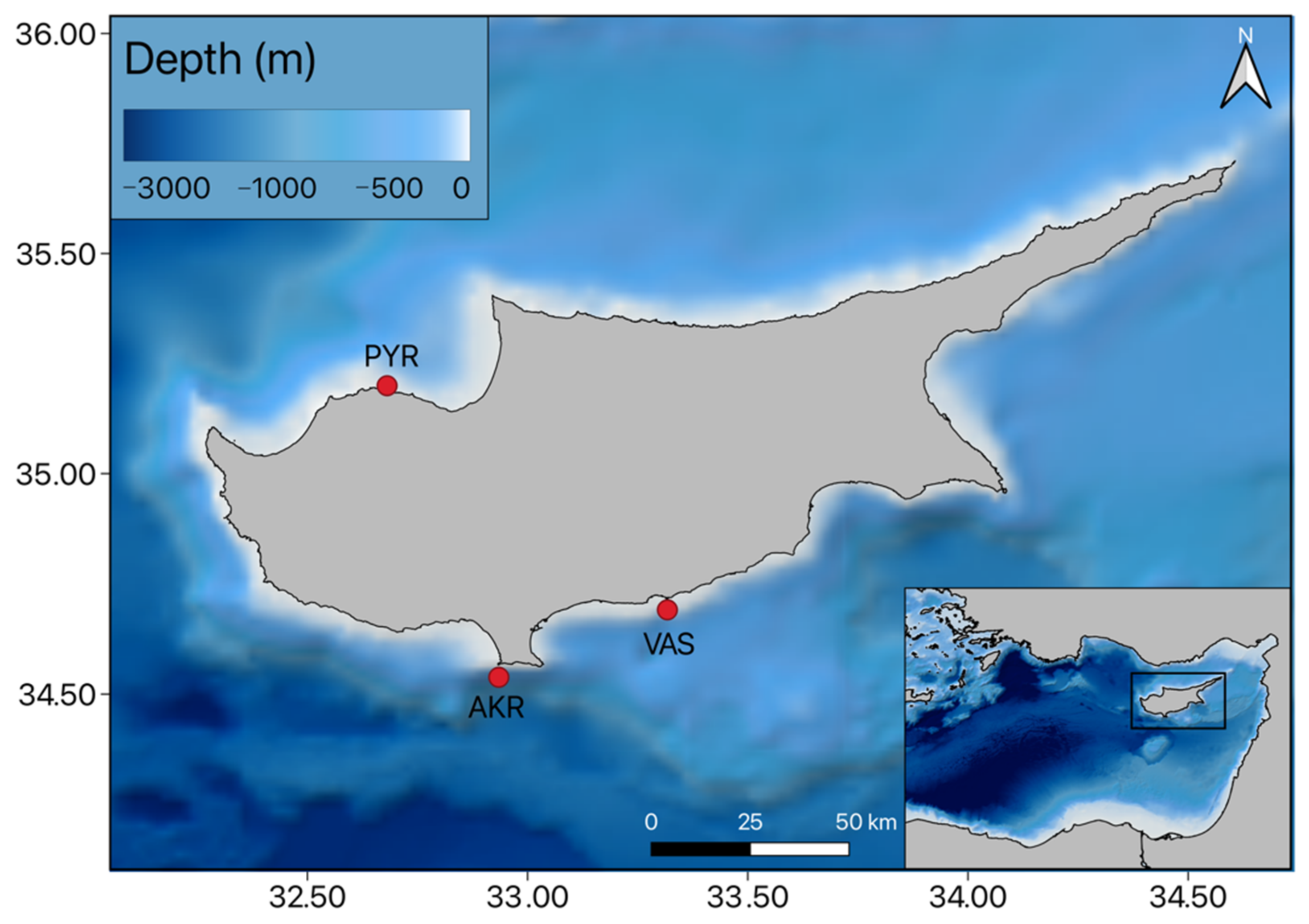
2. Materials and Methods
2.1. Satellite Remote Sensing Data
2.2. in situ Data
2.3. Phytoplankton Pigment-Based Size Classes
2.4. Data Analysis
3. Results
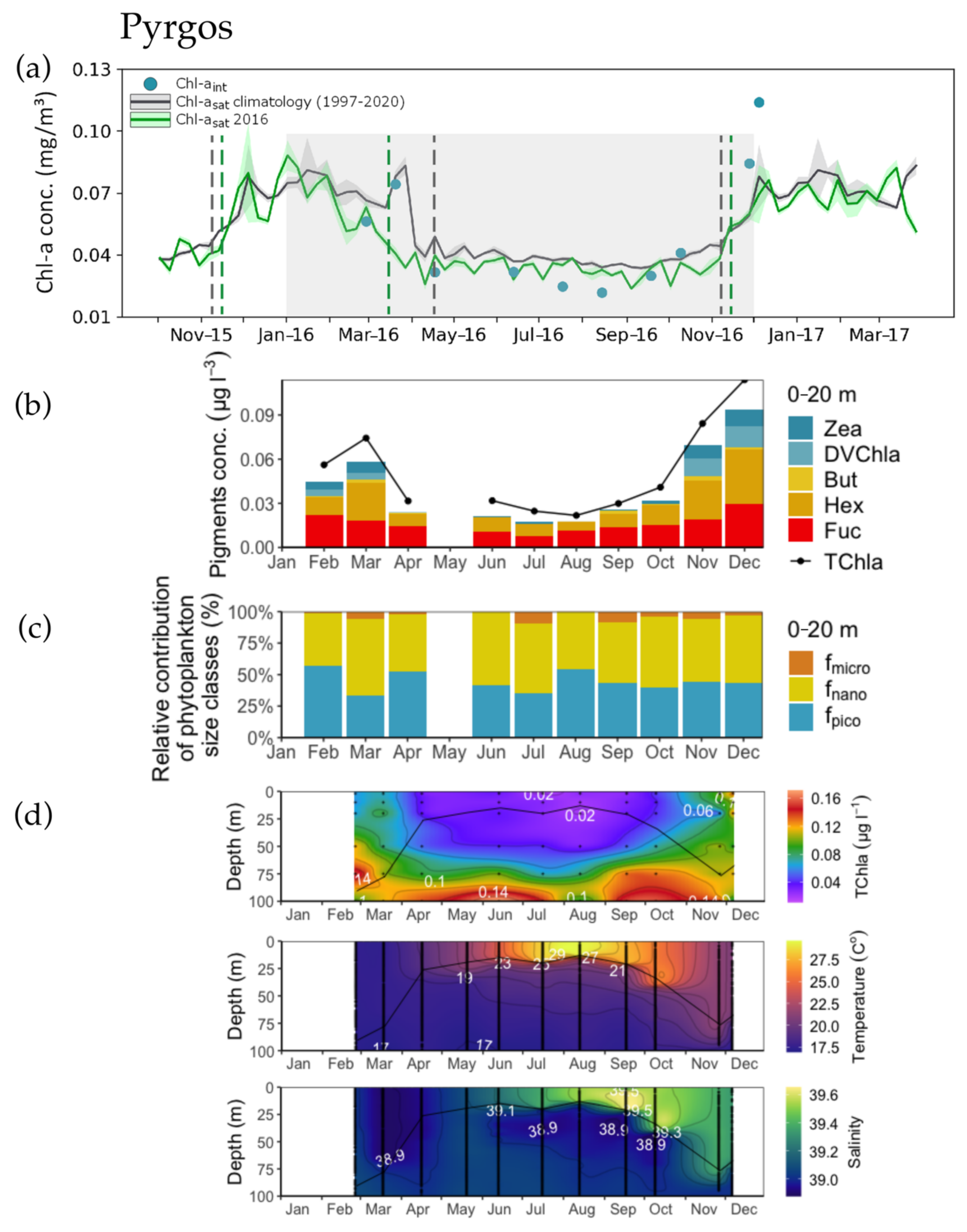
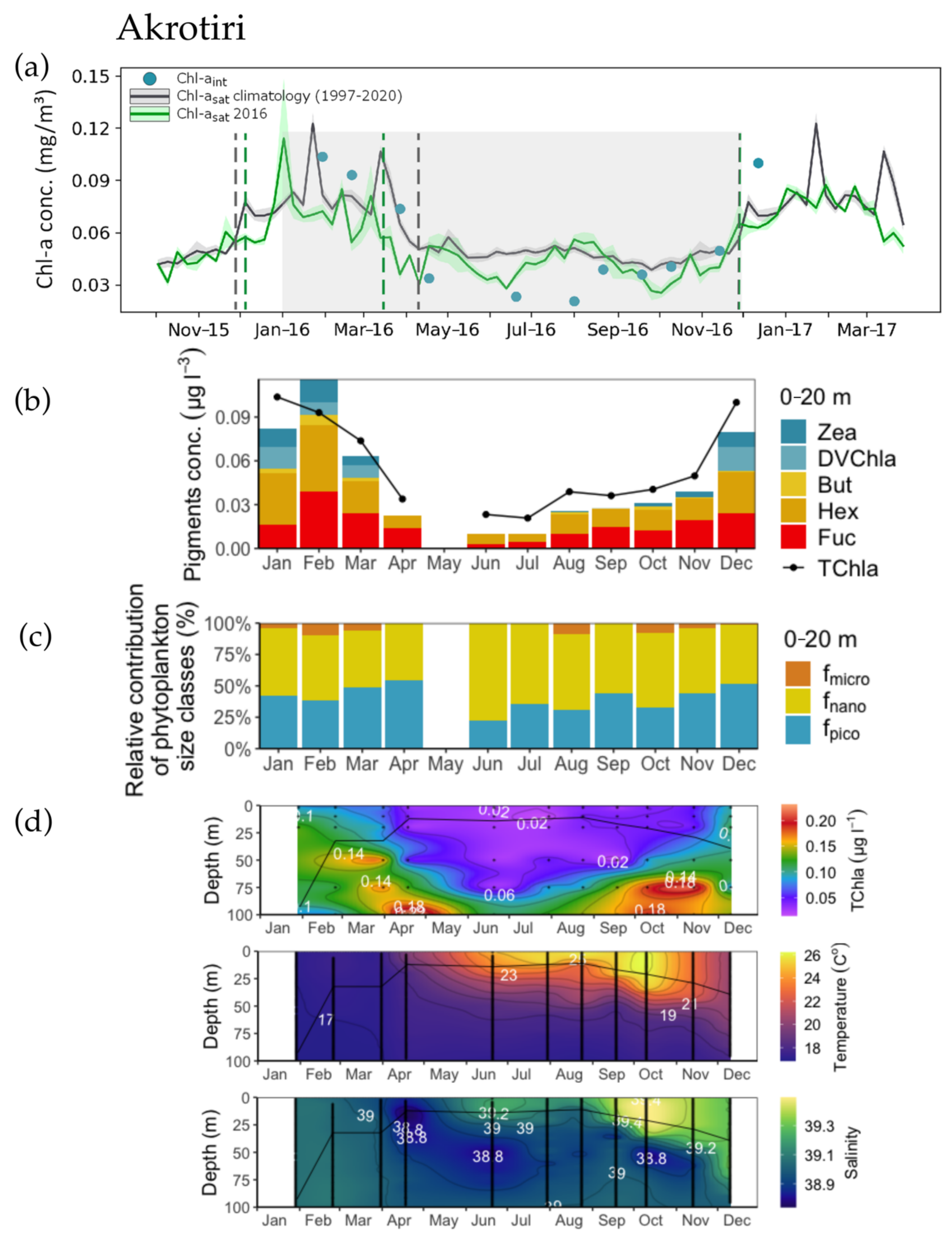
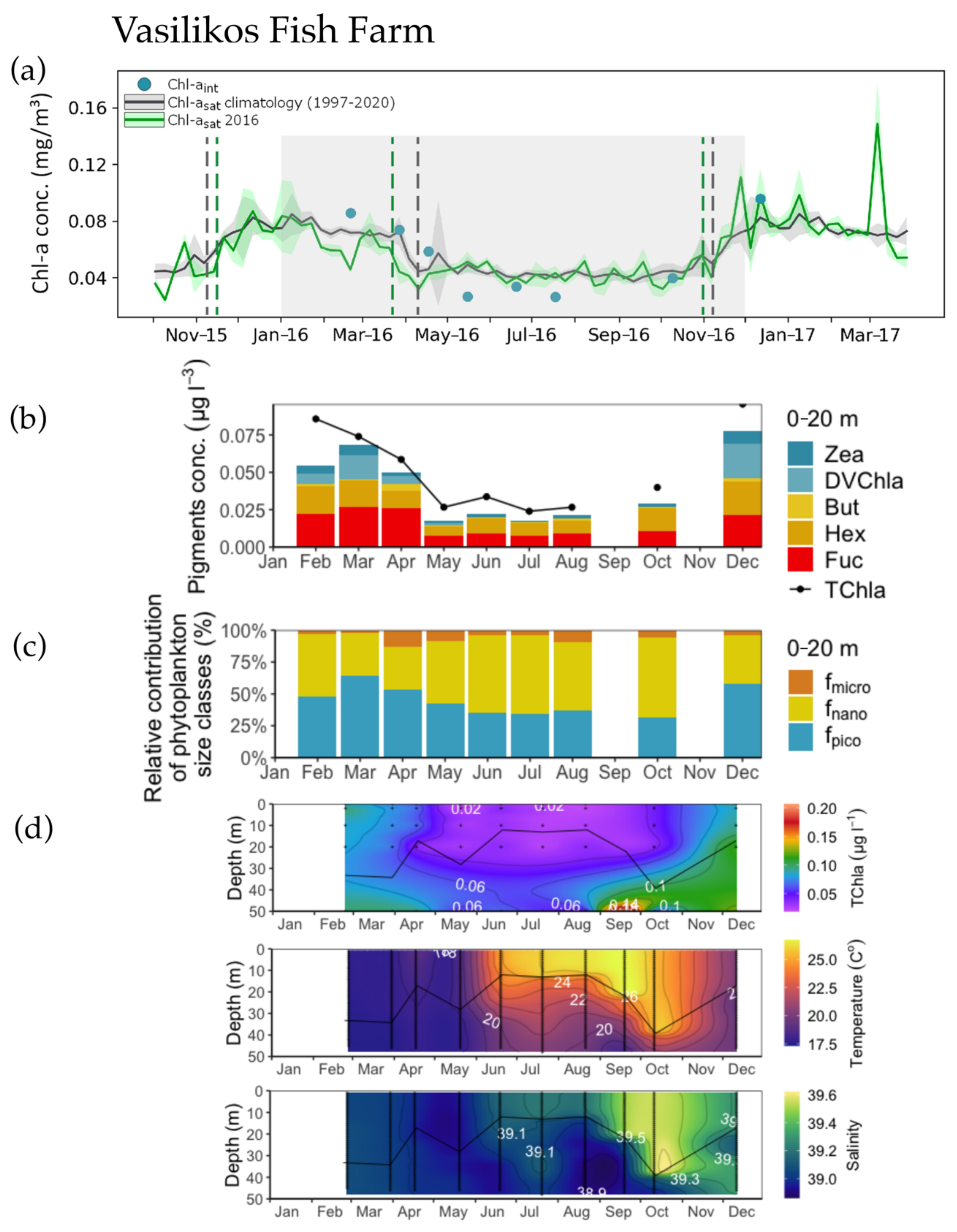
3.1. Phenology Metrics from Satellite and in situ Data Retrievals
3.2. Concentration and Spatial Distribution of Phytoplankton Pigments
3.3. Phytoplankton Size Structure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azov, Y. Eastern Mediterranean—A Marine Desert? Mar. Pollut. Bull. 1991, 23, 225–232. [Google Scholar] [CrossRef]
- Krom, M.D.; Groom, S.; Zohary, T. The Eastern Mediterranean. In Biogeochemistry of Marine Systems; Black, K.D., Shimmield, G.B., Eds.; Blackwell: Oxford, UK, 2003; p. 384. ISBN 978-1-4051-4779-8. [Google Scholar]
- Jeffrey, S.W.; Vesk, M. Introduction to Marine Phytoplankton and Their Pigment Signatures. In Phytoplankton Pigments in Oceanography Guidlines to Modern Methods; Jeffrey, S.W., Mantoura, R.F.C., Wright, S.W., Eds.; UNESCO: Paris, France, 1997. [Google Scholar]
- Berman, T.; Townsend, D.W.; Elsayed, S.Z.; Trees, C.C.; Azov, Y. Optical Transparency, Chlorophyll and Primary Productivity in the Eastern Mediterranean near the Israeli Coast. Oceanol. Acta 1984, 7, 367–372. [Google Scholar]
- Berman, T.; Azov, Y.; Townsend, D. Understanding Oligotrophic Oceans: Can the Eastern Mediterranean Be a Useful Model? In Marine Phytoplankton and Productivity; Holm-Hansen, D.O., Bolis, P.L., Gilles, P.R., Eds.; Lecture Notes on Coastal and Estuarine Studies; Springer: Berlin/Heidelberg, Germany, 1984; pp. 101–112. ISBN 978-3-540-13333-9. [Google Scholar]
- Azov, Y. Seasonal Patterns of Phytoplankton Productivity and Abundance in Nearshore Oligotrophic Waters of the Levant Basin (Mediterranean). J. Plankton Res. 1986, 8, 41–53. [Google Scholar] [CrossRef]
- Li, W.K.W.; Zohary, T.; Yacobi, Y.Z.; Wood, A.M. Ultraphytoplankton in the Eastern Mediterranean Sea: Towards Deriving Phytoplankton Biomass from Flow Cytometric Measurements of Abundance, Fluorescence and Light Scatter. Mar. Ecol. Prog. Ser. 1993, 79–87. [Google Scholar] [CrossRef]
- Yacobi, Y.Z.; Zohary, T.; Kress, N.; Hecht, A.; Robarts, R.D.; Waiser, M.; Wood, A.M.; Li, W.K.W. Chlorophyll Distribution throughout the Southeastern Mediterranean in Relation to the Physical Structure of the Water Mass. J. Mar. Syst. 1995, 6, 179–190. [Google Scholar] [CrossRef]
- Psarra, S.; Tselepides, A.; Ignatiades, L. Primary Productivity in the Oligotrophic Cretan Sea (NE Mediterranean): Seasonal and Interannual Variability. Prog. Oceanogr. 2000, 46, 187–204. [Google Scholar] [CrossRef]
- Christaki, U.; Giannakourou, A.; Van Wambeke, F.; Grégori, G. Nanoflagellate Predation on Auto- and Heterotrophic Picoplankton in the Oligotrophic Mediterranean Sea. J. Plankton Res. 2001, 23, 1297–1310. [Google Scholar] [CrossRef] [Green Version]
- Vidussi, F.; Claustre, H.; Manca, B.B.; Luchetta, A.; Marty, J.-C. Phytoplankton Pigment Distribution in Relation to Upper Thermocline Circulation in the Eastern Mediterranean Sea during Winter. J. Geophys. Res. 2001, 106, 19939–19956. [Google Scholar] [CrossRef]
- Petrou, A.; Kallianiotis, A.; Hannides, A.K.; Charalambidou, I.; Hadjichristoforou, M.; Hayes, D.R.; Lambridis, C.; Lambridi, V.; Loizidou, X.I.; Orfanidis, S.; et al. Initial Assessment of the Marine Environment of Cyprus: Part I - Characteristics; Ministry of Agriculture, Natural Resources, and the Environment, Department of Fisheries and Marine Research: Nicosia, Cyprus, 2012. [Google Scholar]
- EEA EIONET Central Data Repository. Available online: http://cdr.eionet.europa.eu/cy/eea/me1/ (accessed on 7 January 2021).
- Bianchi, T.S.; Demetropoulos, A.; Hadjichristophorou, M.; Argyrou, M.; Baskaran, M.; Lambert, C. Plant Pigments as Biomarkers of Organic Matter Sources in Sediments and Coastal Waters of Cyprus (Eastern Mediterranean). Estuar. Coast. Shelf Sci. 1996, 42, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Sofroniou, A.; Bishop, S. Water Scarcity in Cyprus: A Review and Call for Integrated Policy. Water 2014, 6, 2898–2928. [Google Scholar] [CrossRef] [Green Version]
- WDD Water Resources—Introduction. Available online: http://www.moa.gov.cy/moa/wdd/Wdd.nsf/page08_en/page08_en?opendocument (accessed on 15 December 2020).
- Georgiou, A. Assessment of Groundwater Resources of Cyprus; Reassessment of the island’s water resources and demand-Objective 1–Output 1.4.2; Water Development Department—Ministry of Agriculture, Rural Development and Environment: Nicosia, Cyprus, 2002; p. 234. [Google Scholar]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falkowski, P.G.; Raven, J.A. Aquatic Photosynthesis, 2nd ed.; Princeton University Press: Princeton, NJ, USA, 2007; ISBN 978-0-691-11550-4. [Google Scholar]
- Reynolds, C.S. Ecology of Phytoplankton; Cambridge University Press: Cambridge, NY, USA, 2006; ISBN 978-0-511-19181-7. [Google Scholar]
- Platt, T.; Sathyendranath, S. Ecological Indicators for the Pelagic Zone of the Ocean from Remote Sensing. Remote Sens. Environ. 2008, 112, 3426–3436. [Google Scholar] [CrossRef]
- Edwards, M.; Richardson, A.J. Impact of Climate Change on Marine Pelagic Phenology and Trophic Mismatch. Nature 2004, 430, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Racault, M.-F.; Le Quéré, C.; Buitenhuis, E.; Sathyendranath, S.; Platt, T. Phytoplankton Phenology in the Global Ocean. Ecol. Indic. 2012, 14, 152–163. [Google Scholar] [CrossRef]
- Racault, M.-F.; Platt, T.; Sathyendranath, S.; Airba, E.; Martinez Vicente, V.; Brewin, R. Plankton Indicators and Ocean Observing Systems: Support to the Marine Ecosystem State Assessment. J. Plankton Res. 2014, 36, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Gittings, J.A.; Raitsos, D.E.; Krokos, G.; Hoteit, I. Impacts of Warming on Phytoplankton Abundance and Phenology in a Typical Tropical Marine Ecosystem. Sci. Rep. 2018, 8, 2240. [Google Scholar] [CrossRef]
- Robinson, C. Phytoplankton Biogeochemical Cycles. In Marine Plankton: A Pracitcal Guide to Ecology, Methodology, and Taxonomy; Oxford University Press: Oxford, UK, 2017; pp. 42–51. [Google Scholar]
- Koeller, P.; Fuentes-Yaco, C.; Platt, T.; Sathyendranath, S.; Richards, A.; Ouellet, P.; Orr, D.; Skúladóttir, U.; Wieland, K.; Savard, L.; et al. Basin-Scale Coherence in Phenology of Shrimps and Phytoplankton in the North Atlantic Ocean. Science 2009, 324, 791–793. [Google Scholar] [CrossRef]
- Cushing, D.H. Plankton Production and Year-Class Strength in Fish Populations: An Update of the Match/Mismatch Hypothesis. Adv. Mar. Biol. 1990, 26, 249–293. [Google Scholar] [CrossRef]
- Platt, T.; Fuentes-Yaco, C.; Frank, K.T. Spring Algal Bloom and Larval Fish Survival. Nature 2003, 423, 398–399. [Google Scholar] [CrossRef]
- Gittings, J.A.; Raitsos, D.E.; Brewin, R.J.W.; Hoteit, I. Links between Phenology of Large Phytoplankton and Fisheries in the Northern and Central Red Sea. Remote Sensing 2021, 13, 231. [Google Scholar] [CrossRef]
- Wright, S.W.; Jeffrey, S.W. Pigment Markers for Phytoplankton Production. In Marine Organic Matter: Biomarkers, Isotopes and DNA; Volkman, J.K., Ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 2006; Volume 2N, pp. 71–104. ISBN 978-3-540-28401-7. [Google Scholar]
- Volpe, G.; Colella, S.; Brando, V.E.; Forneris, V.; La Padula, F.; Di Cicco, A.; Sammartino, M.; Bracaglia, M.; Artuso, F.; Santoleri, R. Mediterranean Ocean Colour Level 3 Operational Multi-Sensor Processing. Ocean Sci. 2019, 15, 127–146. [Google Scholar] [CrossRef] [Green Version]
- Berthon, J.-F.; Zibordi, G. Bio-Optical Relationships for the Northern Adriatic Sea. Int. J. Remote Sens. 2004, 25, 1527–1532. [Google Scholar] [CrossRef]
- D’Ortenzio, F.; Santoleri, R.; Marullo, S.; Ragni, M.; Ribera d’Alcala, M. Empirical SeaWiFS Chlorophyll Algorithm Validation for the Mediterranean Sea; Bostater, C.R., Jr., Santoleri, R., Eds.; SPIE: Barcelona, Spain, 2000; pp. 124–131. [Google Scholar]
- Volpe, G.; Santoleri, R.; Vellucci, V.; Ribera d’Alcalà, M.; Marullo, S.; D’Ortenzio, F. The Colour of the Mediterranean Sea: Global versus Regional Bio-Optical Algorithms Evaluation and Implication for Satellite Chlorophyll Estimates. Remote Sens. Environ. 2007, 107, 625–638. [Google Scholar] [CrossRef]
- Brewin, R.J.W.; Raitsos, D.E.; Pradhan, Y.; Hoteit, I. Comparison of Chlorophyll in the Red Sea Derived from MODIS-Aqua and in Vivo Fluorescence. Remote Sens. Environ. 2013, 136, 218–224. [Google Scholar] [CrossRef]
- Racault, M.-F.; Raitsos, D.E.; Berumen, M.L.; Brewin, R.J.W.; Platt, T.; Sathyendranath, S.; Hoteit, I. Phytoplankton Phenology Indices in Coral Reef Ecosystems: Application to Ocean-Color Observations in the Red Sea. Remote Sens. Environ. 2015, 160, 222–234. [Google Scholar] [CrossRef] [Green Version]
- Gittings, J.A.; Raitsos, D.E.; Kheireddine, M.; Racault, M.-F.; Claustre, H.; Hoteit, I. Evaluating Tropical Phytoplankton Phenology Metrics Using Contemporary Tools. Sci. Rep. 2019, 9, 674. [Google Scholar] [CrossRef]
- Salgado-Hernanz, P.M.; Racault, M.-F.; Font-Muñoz, J.S.; Basterretxea, G. Trends in Phytoplankton Phenology in the Mediterranean Sea Based on Ocean-Colour Remote Sensing. Remote Sens. Environ. 2019, 221, 50–64. [Google Scholar] [CrossRef]
- Van Heukelem, L.; Thomas, C.S. Computer-Assisted High-Performance Liquid Chromatography Method Development with Applications to the Isolation and Analysis of Phytoplankton Pigments. J. Chromatogr. A 2001, 910, 31–49. [Google Scholar] [CrossRef]
- Lagaria, A.; Mandalakis, M.; Mara, P.; Frangoulis, C.; Karatsolis, B.-T.; Pitta, P.; Triantaphyllou, M.; Tsiola, A.; Psarra, S. Phytoplankton Variability and Community Structure in Relation to Hydrographic Features in the NE Aegean Frontal Area (NE Mediterranean Sea). Cont. Shelf Res. 2017, 149, 124–137. [Google Scholar] [CrossRef]
- Mandalakis, M.; Stravinskaitė, A.; Lagaria, A.; Psarra, S.; Polymenakou, P. Ultrasensitive and High-Throughput Analysis of Chlorophyll a in Marine Phytoplankton Extracts Using a Fluorescence Microplate Reader. Anal. Bioanal. Chem. 2017, 409, 4539–4549. [Google Scholar] [CrossRef]
- Laws, E.A. Mathematical Methods for Oceanographers: An Introduction; Wiley: New York, NY, USA, 1997; ISBN 978-0-471-16221-6. [Google Scholar]
- Al-Naimi, N.; Raitsos, D.E.; Ben-Hamadou, R.; Soliman, Y. Evaluation of Satellite Retrievals of Chlorophyll-a in the Arabian Gulf. Remote Sens. 2017, 9, 301. [Google Scholar] [CrossRef] [Green Version]
- De Boyer Montégut, C. Mixed Layer Depth over the Global Ocean: An Examination of Profile Data and a Profile-Based Climatology. J. Geophys. Res. 2004, 109, C12003. [Google Scholar] [CrossRef]
- Amante, C.; Eakins, B.W. ETOPO1 1 Arc-Minute Global Relief Model: Procedures, Data Sources and Analysis; National Geophysical Data Center: Boulder, CO, USA, 2009. [Google Scholar]
- Amante, C.; Eakins, B.W. ETOPO1 1 Arc-Minute Global Relief Model; NOAA National Geophysical Data Center: Boulder, CO, USA, 2009. [Google Scholar]
- Uitz, J.; Claustre, H.; Morel, A.; Hooker, S.B. Vertical Distribution of Phytoplankton Communities in Open Ocean: An Assessment Based on Surface Chlorophyll. J. Geophys. Res. 2006, 111, C08005. [Google Scholar] [CrossRef]
- Gittings, J.A.; Brewin, R.J.W.; Raitsos, D.E.; Kheireddine, M.; Ouhssain, M.; Jones, B.H.; Hoteit, I. Remotely Sensing Phytoplankton Size Structure in the Red Sea. Remote Sens. Environ. 2019, 234, 111387. [Google Scholar] [CrossRef]
- Brewin, R.J.W.; Morán, X.A.G.; Raitsos, D.E.; Gittings, J.A.; Calleja, M.L.; Viegas, M.; Ansari, M.I.; Al-Otaibi, N.; Huete-Stauffer, T.M.; Hoteit, I. Factors Regulating the Relationship Between Total and Size-Fractionated Chlorophyll-a in Coastal Waters of the Red Sea. Front. Microbiol. 2019, 10, 1964. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, S.W.; Wright, S.W.; Zapata, M. Microalgal Classes and Their Signature Pigments. In Phytoplankton Pigments: Characterization, Chemotaxonomy, and Applications in Oceanography; Cambridge University Press: Cambridge, NY, USA, 2011; ISBN 978-1-107-00066-7. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Finley, A.; Banerjee, S.; Hjelle, Ø. MBA: Multilevel B-Spline Approximation; CRAN, 2017. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Barbieux, M.; Uitz, J.; Gentili, B.; Pasqueron de Fommervault, O.; Mignot, A.; Poteau, A.; Schmechtig, C.; Taillandier, V.; Leymarie, E.; Penkerc’h, C.; et al. Bio-Optical Characterization of Subsurface Chlorophyll Maxima in the Mediterranean Sea from a Biogeochemical-Argo Float Database. Biogeosciences 2019, 16, 1321–1342. [Google Scholar] [CrossRef] [Green Version]
- Krom, M.D.; Thingstad, T.F.; Brenner, S.; Carbo, P.; Drakopoulos, P.; Fileman, T.W.; Flaten, G.A.F.; Groom, S.; Herut, B.; Kitidis, V.; et al. Summary and Overview of the CYCLOPS P Addition Lagrangian Experiment in the Eastern Mediterranean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2005, 52, 3090–3108. [Google Scholar] [CrossRef]
- Salgado-Hernanz, P.M. Patterns of Phytoplankton and Primary Production Variability in the Mediterranean Sea Based on Remote Sensing Data. Ph.D. Thesis, Universitat de les Illes Balears, Palma, Spain, 2019. [Google Scholar]
- Gallisai, R.; Peters, F.; Volpe, G.; Basart, S.; Baldasano, J.M. Saharan Dust Deposition May Affect Phytoplankton Growth in the Mediterranean Sea at Ecological Time Scales. PLoS ONE 2014, 9, e110762. [Google Scholar] [CrossRef] [Green Version]
- Deflorio, M.; Aprea, A.; Corriero, A.; Santamaria, N.; De Metrio, G. Incidental Captures of Sea Turtles by Swordfish and Albacore Longlines in the Ionian Sea. Fish. Sci. 2005, 71, 1010–1018. [Google Scholar] [CrossRef]
- D’Ortenzio, F.; Ribera d’Alcalà, M. On the Trophic Regimes of the Mediterranean Sea: A Satellite Analysis. Biogeosciences 2009, 6, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Yücel, N. Monthly Changes in Primary and Bacterial Productivity in the North-Eastern Mediterranean Shelf Waters. Ph.D. Thesis, Middle East Technical University, Ankara, Turkey, 2013. [Google Scholar]
- Siokou-Frangou, I.; Christaki, U.; Mazzocchi, M.G.; Montresor, M.; Ribera d’Alcalá, M.; Vaqué, D.; Zingone, A. Plankton in the Open Mediterranean Sea: A Review. Biogeosciences 2010, 7, 1543–1586. [Google Scholar] [CrossRef] [Green Version]
- Tsagaraki, T.; Pitta, P.; Frangoulis, C.; Petihakis, G.; Karakassis, I. Plankton Response to Nutrient Enrichment Is Maximized at Intermediate Distances from Fish Farms. Mar. Ecol. Prog. Ser. 2013, 493, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Zohary, T.; Krom, M.D.; Law, C.S.; Pitta, P.; Psarra, S.; Rassoulzadegan, F.; Thingstad, T.F.; Tselepides, A.; Woodward, E.M.S.; et al. Microbial Community Structure and Function in the Levantine Basin of the Eastern Mediterranean. Deep Sea Res. Part I Oceanogr. Res. Pap. 2007, 54, 1721–1743. [Google Scholar] [CrossRef]
- Zohary, T.; Brenner, S.; Krom, M.; Angel, D.; Kress, N.; Li, W.; Neori, A.; Yacobi, Y. Buildup of Microbial Biomass during Deep Winter Mixing in a Mediterranean Warm-Core Eddy. Mar. Ecol. Prog. Ser. 1998, 167, 47–57. [Google Scholar] [CrossRef]
- Psarra, S.; Zohary, T.; Krom, M.D.; Mantoura, R.F.C.; Polychronaki, T.; Stambler, N.; Tanaka, T.; Tselepides, A.; Frede Thingstad, T. Phytoplankton Response to a Lagrangian Phosphate Addition in the Levantine Sea (Eastern Mediterranean). Deep Sea Res. Part II Top. Stud. Oceanogr. 2005, 52, 2944–2960. [Google Scholar] [CrossRef]
- Uitz, J.; Stramski, D.; Gentili, B.; D’Ortenzio, F.; Claustre, H. Estimates of Phytoplankton Class-Specific and Total Primary Production in the Mediterranean Sea from Satellite Ocean Color Observations. Global Biogeochem. Cycles 2012, 26, GB2024. [Google Scholar] [CrossRef]
- Yücel, N. Spatio-Temporal Variability of the Size-Fractionated Primary Production and Chlorophyll in the Levantine Basin (Northeastern Mediterranean). Oceanologia 2018, 60, 288–304. [Google Scholar] [CrossRef]
- Zodiatis, G.; Lardner, R.; Hayes, D.R.; Georgiou, G.; Sofianos, S.; Skliris, N.; Lascaratos, A. Operational Ocean Forecasting in the Eastern Mediterranean: Implementation and Evaluation. Ocean Sci. 2008, 4. [Google Scholar] [CrossRef] [Green Version]
- Mauri, E.; Lardner, R.; Sitz, L.; Gerin, R.; Poulain, P.-M.; Hayes, D.; Gildor, H. On the Variability of the Circulation and Water Mass Properties in the Eastern Levantine Sea between September 2016–August 2017. Water 2019, 11, 1741. [Google Scholar] [CrossRef] [Green Version]
- Ediger, D.; Yilmaz, A. Characteristics of Deep Chlorphyll Maximum in the Northeastern Mediterranean with Respect to Environmental Conditions. J. Mar. Syst. 1996, 9, 291–303. [Google Scholar] [CrossRef]
- Krom, M.D.; Herut, B.; Mantoura, R.F.C. Nutrient Budget for the Eastern Mediterranean: Implications for Phosphorus Limitation. Limnol. Oceanogr. 2004, 49, 1582–1592. [Google Scholar] [CrossRef]
- Herut, B.; Zohary, T.; Krom, M.D.; Mantoura, R.F.C.; Pitta, P.; Psarra, S.; Rassoulzadegan, F.; Tanaka, T.; Frede Thingstad, T. Response of East Mediterranean Surface Water to Saharan Dust: On-Board Microcosm Experiment and Field Observations. Deep Sea Research Part II: Topical Studies in Oceanography 2005, 52, 3024–3040. [Google Scholar] [CrossRef]
- Markaki, Z.; Oikonomou, K.; Kocak, M.; Kouvarakis, G.; Chaniotaki, A.; Kubilay, N.; Mihalopoulos, N. Atmospheric Deposition of Inorganic Phosphorus in the Levantine Basin, Eastern Mediterranean: Spatial and Temporal Variability and Its Role in Seawater Productivity. Limnol. Oceanogr. 2003, 48, 1557–1568. [Google Scholar] [CrossRef] [Green Version]
- Markaki, Z.; Loÿe-Pilot, M.D.; Violaki, K.; Benyahya, L.; Mihalopoulos, N. Variability of Atmospheric Deposition of Dissolved Nitrogen and Phosphorus in the Mediterranean and Possible Link to the Anomalous Seawater N/P Ratio. Mar. Chem. 2010, 120, 187–194. [Google Scholar] [CrossRef]
- Brewin, R.J.W.; Sathyendranath, S.; Hirata, T.; Lavender, S.J.; Barciela, R.M.; Hardman-Mountford, N.J. A Three-Component Model of Phytoplankton Size Class for the Atlantic Ocean. Ecol. Model. 2010, 221, 1472–1483. [Google Scholar] [CrossRef]
- Sammartino, M.; Di Cicco, A.; Marullo, S.; Santoleri, R. Spatio-Temporal Variability of Micro-, Nano- and Pico-Phytoplankton in the Mediterranean Sea from Satellite Ocean Colour Data of SeaWiFS. Ocean Sci. 2015, 11, 759–778. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pigments | Abbreviations | Taxonomic Significance | Size μm |
|---|---|---|---|
| Zeaxanthin | Zea | Cyanobacteria and Prochlorophytes | <2 |
| Divinyl-chlorophyll a | DVChl-a | Prochlorophytes | <2 |
| 19′ hexanoyloxyfucoxanthin | Hex | Prymnesiophytes (major) | 2–20 |
| 19′ butanoyloxyfucoxanthin | But | Pelagophytes (major), Prymnesiophytes | 2–20 |
| Alloxanthin | Allo | Cryptophytes | 2–20 |
| Fucoxanthin | Fuc | Diatoms (major), Prymnesiophytes | >20 |
| Peridinin | Peri | Dinoflagellates | >20 |
| Between Stations | Between Seasons | Between Depths | ||||||
|---|---|---|---|---|---|---|---|---|
| Water Column | 0–20 m | |||||||
| Variable | F | p | F | p | F | p | F | p |
| TChl-a | (2, 20) = 0.68 | (1, 21) = 2.25 | (1, 17) = 6.07 | * | (1, 41) = 15.89 | *** | ||
| Zea | (2, 20) = 0.18 | (1, 21) = 6.78 | * | (1, 17) = 8.20 | ** | (1, 41) = 0.63 | ||
| DVChl-a | (2, 20) = 1.35 | (1, 21) = 0.03 | (1, 9) = 0.007 | (1, 33) = 3.00 | ||||
| But | (2, 20) = 1.30 | (1, 21) = 1.42 | (1, 17) = 6.02 | * | (1, 41) = 10.16 | ** | ||
| Hex | (2, 20) = 6.81 | ** | (1, 21) = 0.69 | (1, 17) = 2.12 | (1, 41) = 19.17 | *** | ||
| Fuc | (2, 20) = 0.40 | (1, 21) = 1.90 | (1, 16) = 1.83 | (1, 41) = 2.40 | * | |||
| fpico | (2, 20) = 4.65 | * | (1, 21) = 0.32 | (1, 17) = 4.60 | * | (1, 41) = 2.40 | ||
| fnano | (2, 20) = 5.67 | ** | (1, 21) = 0.75 | (1, 17) = 5.56 | * | (1, 41) = 9.57 | ** | |
| fmicro | (2, 20) = 1.74 | (1, 21) = 1.24 | (1, 17) = 0.92 | (1, 41) = 0.06 | ||||
| fpico (%) | fnano (%) | fmicro (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| Station | Period | Depth | Range | Mean (±SD) | Range | Mean (±SD) | Range | Mean (±SD) |
| PYR | Mixed | 0–20 | 34–52 | 44 ± 9 | 37–61 | 46 ± 13 | 2–6 | 3 ± 2 |
| 0–100 | 38–50 | 44 ± 6 | 46–57 | 51 ± 6 | 2–5 | 3 ± 1 | ||
| Stratified | 0–20 | 35–54 | 43 ± 6 | 45–59 | 52 ± 5 | 3–10 | 5 ± 4 | |
| 0–100 | 36–54 | 47 ± 6 | 42–55 | 48 ± 4 | 2–11 | 5 ± 3 | ||
| AKR | Mixed | 0–20 | 38–54 | 46 ± 7 | 45–54 | 49 ± 4 | 4–10 | 7 ± 3 |
| 0–100 | 33–53 | 43 ± 1 | 45–57 | 51 ± 6 | 2–10 | 6 ± 4 | ||
| Stratified | 0–20 | 22–52 | 37 ± 1 | 47–78 | 60 ± 10 | 3–8 | 4 ± 4 | |
| 0–100 | 26–57 | 39 ± 13 | 42–73 | 58 ± 13 | 1–7 | 3 ± 2 | ||
| VAS | Mixed | 0–20 | 48–64 | 55 ± 8 | 34–49 | 39 ± 9 | 2–13 | 6 ± 6 |
| 0–50 | 47–64 | 55 ± 9 | 32–49 | 39 ± 8 | 4–10 | 6 ± 4 | ||
| Stratified | 0–20 | 30–58 | 40 ± 10 | 38–62 | 54 ± 9 | 4–10 | 6 ± 3 | |
| 0–50 | 20–55 | 38 ± 13 | 40–73 | 55 ± 13 | 5–9 | 7 ± 2 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demetriou, M.; Raitsos, D.E.; Kournopoulou, A.; Mandalakis, M.; Sfenthourakis, S.; Psarra, S. Phytoplankton Phenology in the Coastal Zone of Cyprus, Based on Remote Sensing and In Situ Observations. Remote Sens. 2022, 14, 12. https://doi.org/10.3390/rs14010012
Demetriou M, Raitsos DE, Kournopoulou A, Mandalakis M, Sfenthourakis S, Psarra S. Phytoplankton Phenology in the Coastal Zone of Cyprus, Based on Remote Sensing and In Situ Observations. Remote Sensing. 2022; 14(1):12. https://doi.org/10.3390/rs14010012
Chicago/Turabian StyleDemetriou, Monica, Dionysios E. Raitsos, Antonia Kournopoulou, Manolis Mandalakis, Spyros Sfenthourakis, and Stella Psarra. 2022. "Phytoplankton Phenology in the Coastal Zone of Cyprus, Based on Remote Sensing and In Situ Observations" Remote Sensing 14, no. 1: 12. https://doi.org/10.3390/rs14010012
APA StyleDemetriou, M., Raitsos, D. E., Kournopoulou, A., Mandalakis, M., Sfenthourakis, S., & Psarra, S. (2022). Phytoplankton Phenology in the Coastal Zone of Cyprus, Based on Remote Sensing and In Situ Observations. Remote Sensing, 14(1), 12. https://doi.org/10.3390/rs14010012