
Exploring the Potential of SCOPE Model for Detection of Leaf Area Index and Sun-Induced Fluorescence of Peatland Canopy

 ,
,  ,
,  , ,
, ,
Abstract
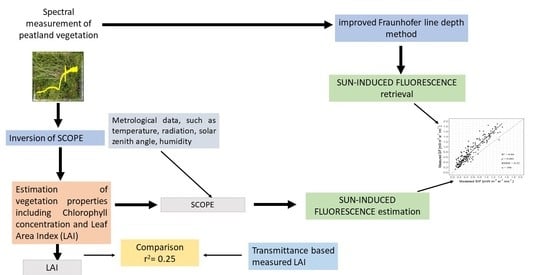
:
1. Introduction
2. Material and Method
2.1. Experimental Site
2.2. Measurement of Hyperspectral and Vegetation Properties
2.3. Implementation of SCOPE
2.4. Statistical Analysis
3. Results
3.1. Measured LAI
3.2. Vegetation Indices
3.3. Modeled LAI
3.4. Measured and Modeled SIF
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gorham, E. Northern peatlands: Role in the carbon cycle and probable responses to climatic warming. Ecol. Appl. 1991, 1, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Samson, M.; Słowińska, S.; Słowiński, M.; Lamentowicz, M.; Barabach, J.; Harenda, K.; Zielińska, M.; Robroek, B.; Jassey, V.; Buttler, A.; et al. The Impact of Experimental Temperature and Water Level Manipulation on Carbon Dioxide Release in a Poor Fen in Northern Poland. Wetlands 2018, 38, 551–563. [Google Scholar] [CrossRef]
- Rastogi, A.; Stróżecki, M.; Kalaji, H.M.; Łuców, D.; Lamentowicz, M.; Juszczak, R. Impact of warming and reduced precipitation on photosynthetic and remote sensing properties of peatland vegetation. Environ. Exp. Bot. 2019, 160, 71–80. [Google Scholar] [CrossRef]
- Loisel, J.; Gallego-Sala, A.V.; Amesbury, M.J.; Magnan, G.; Anshari, G.; Beilman, D.W.; Benavides, J.C.; Blewett, J.; Camill, P.; Charman, D.J.; et al. Expert assessment of future vulnerability of the global peatland carbon sink. Nat. Clim. Chang. 2021, 11, 70–77. [Google Scholar] [CrossRef]
- Belyea, L.R. Nonlinear dynamics of peatlands and potential feedbacks on the climate system. In Geophysical Monograph Series; Baird, A.J., Belyea, L.R., Comas, X., Reeve, A.S., Slater, L.D., Eds.; American Geophysical Union: Washington, DC, USA, 2013; pp. 5–18. ISBN 978-1-118-66666-1. [Google Scholar]
- Wang, R.; Gamon, J.A. Remote sensing of terrestrial plant biodiversity. Remote Sens. Environ. 2019, 231, 111218. [Google Scholar] [CrossRef]
- Chasmer, L.; Mahoney, C.; Millard, K.; Nelson, K.; Peters, D.; Merchant, M.; Hopkinson, C.; Brisco, B.; Niemann, O.; Montgomery, J.; et al. Remote Sensing of Boreal Wetlands 2: Methods for Evaluating Boreal Wetland Ecosystem State and Drivers of Change. Remote Sens. 2020, 12, 1321. [Google Scholar] [CrossRef]
- Xue, J.; Su, B. Significant Remote Sensing Vegetation Indices: A Review of Developments and Applications. J. Sens. 2017, 2017, 1353691. [Google Scholar] [CrossRef]
- Sishodia, R.P.; Ray, R.L.; Singh, S.K. Applications of Remote Sensing in Precision Agriculture: A Review. Remote Sens. 2020, 12, 3136. [Google Scholar] [CrossRef]
- Badgley, G.; Field, C.B.; Berry, J.A. Canopy near-infrared reflectance and terrestrial photosynthesis. Sci. Adv. 2017, 3, e1602244. [Google Scholar] [CrossRef]
- Meroni, M.; Rossini, M.; Guanter, L.; Alonso, L.; Rascher, U.; Colombo, R.; Moreno, J. Remote sensing of solar-induced chlorophyll fluorescence: Review of methods and applications. Remote Sens. Environ. 2009, 113, 2037–2051. [Google Scholar] [CrossRef]
- Bandopadhyay, S.; Rastogi, A.; Juszczak, R. Review of top-of-canopy sun-induced fluorescence (SIF) studies from ground, uav, airborne to spaceborne observations. Sensors 2020, 20, 1144. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; van der Tol, C.; Campbell, P.K.E.; Middleton, E.M. Fluorescence Correction Vegetation Index (FCVI): A physically based reflectance index to separate physiological and non-physiological information in far-red sun-induced chlorophyll fluorescence. Remote Sens. Environ. 2020, 240, 111676. [Google Scholar] [CrossRef]
- Harris, A.; Charnock, R.; Lucas, R.M. Hyperspectral Remote Sensing of Peatland Floristic Gradients. Remote Sens. Environ. 2015, 162, 99–111. [Google Scholar] [CrossRef]
- Kalacska, M.; Lalonde, M.; Moore, T. Estimation of foliar chlorophyll and nitrogen content in an ombrotrophic bog from hyperspectral data: Scaling from leaf to image. Remote Sens. Environ. 2015, 169, 270–279. [Google Scholar] [CrossRef]
- McPartland, M.Y.; Falkowski, M.J.; Reinhardt, J.R.; Kane, E.S.; Kolka, R.; Turetsky, M.R.; Douglas, T.A.; Anderson, J.; Edwards, J.D.; Palik, B.; et al. Characterizing Boreal Peatland Plant Composition and Species Diversity with Hyperspectral Remote Sensing. Remote Sens. 2019, 11, 1685. [Google Scholar] [CrossRef]
- Granlund, L.; Vesakoski, V.; Sallinen, A.; Kolari, T.H.M.; Wolff, F.; Tahvanainen, T. Recent Lateral Expansion of Sphagnum Bogs Over Central Fen Areas of Boreal Aapa Mire Complexes. Ecosystems 2021. [Google Scholar] [CrossRef]
- Górecki, K.; Rastogi, A.; Stróżecki, M.; Gąbka, M.; Lamentowicz, M.; Łuców, D.; Kayzer, D.; Juszczak, R. Water table depth, experimental warming, and reduced precipitation impact on litter decomposition in a temperate Sphagnum-peatland. Sci. Total Environ. 2021, 771, 145452. [Google Scholar] [CrossRef]
- Bréda, N.J.J. Leaf area index S. In Encyclopedia of Ecology; Brian, E.F., Ed.; Academic Press: Oxford, UK, 2008; pp. 2148–2154. [Google Scholar]
- Van der Tol, C.; Rossini, M.; Cogliati, S.; Verhoef, W.; Colombo, R.; Rascher, U.; Mohammed, G. A model and measurement comparison of diurnal cycles of sun-induced chlorophyll fluorescence of crops. Remote Sens. Environ. 2016, 186, 663–677. [Google Scholar] [CrossRef]
- Celesti, M.; van der Tol, C.; Cogliati, S.; Panigada, C.; Yang, P.; Pinto, F.; Rascher, U.; Miglietta, F.; Colombo, R.; Rossini, M. Exploring the physiological information of Sun-induced chlorophyll fluorescence through radiative transfer model inversion. Remote Sens. Environ. 2018, 215, 97–108. [Google Scholar] [CrossRef]
- Shan, N.; Zhang, Y.; Chen, J.M.; Ju, W.; Migliavacca, M.; Peñuelas, J.; Yang, X.; Zhang, Z.; Nelson, J.A.; Goulas, Y. A model for estimating transpiration from remotely sensed solar-induced chlorophyll fluorescence. Remote Sens. Environ. 2021, 252, 112134. [Google Scholar] [CrossRef]
- Yang, P.; Prikaziuk, E.; Verhoef, W.; van der Tol, C. SCOPE 2.0: A model to simulate vegetated land surface fluxes and satellite signals. Geosci. Model Dev. 2021, 14, 4697–4712. [Google Scholar] [CrossRef]
- Pacheco-Labrador, J.; El-Madany, T.S.; van der Tol, C.; Martin, M.P.; Gonzalez-Cascon, R.; Perez-Priego, O.; Guan, J.; Moreno, G.; Carrara, A.; Reichstein, M.; et al. senSCOPE: Modeling mixed canopies combining green and brown senesced leaves. Evaluation in a Mediterranean Grassland. Remote Sens. Environ. 2021, 257, 112352. [Google Scholar] [CrossRef]
- De Cannière, S.; Herbst, M.; Vereecken, H.; Defourny, P.; Jonard, F. Constraining water limitation of photosynthesis in a crop growth model with sun-induced chlorophyll fluorescence. Remote Sens. Environ. 2021, 267, 112722. [Google Scholar] [CrossRef]
- Bandopadhyay, S.; Rastogi, A.; Rascher, U.; Rademske, P.; Schickling, A.; Cogliati, S.; Julitta, T.; Mac Arthur, A.; Hueni, A.; Tomelleri, E.; et al. Hyplant-derived Sun-Induced Fluorescence-A new opportunity to disentangle complex vegetation signals from diverse vegetation types. Remote Sens. 2019, 11, 1691. [Google Scholar] [CrossRef]
- Mac Arthur, A.; Robinson, I.; Hagdorn, M.; Wood, J.; Kershaw, R.; Taylor, R. Piccolo spectrometer system for reflectance and fluorescence measurement from mobile and fixed platforms. In Proceedings of the Innovative Optical Tools for Proximal Sensing of Ecophysiological Processes (OPTIMISE), Vienna, Austria, 23–28 April 2017. [Google Scholar]
- Alonso, L.; Gómez-Chova, L.; Vila-Francés, J.; Amorós-López, J.; Guanter, L.; Calpe, J.; Moreno, J. Improved fraunhofer line discrimination method for vegetation fluorescence quantification. IEEE Geosci. Remote Sens. Lett. 2008, 5, 620–624. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. NASA Spec. Publ. 1974, 351, 309. [Google Scholar]
- Liu, H.Q.; Huete, A.R. A feedback based modification of the NDVI to minimize canopy background and atmospheric noise. IEEE Trans. Geosci. Remote Sens. 1995, 33, 457–465. [Google Scholar] [CrossRef]
- Rastogi, A.; Bandopadhyay, S.; Strózecki, M.; Juszczak, R. Monitoring the impact of environmental manipulation on peatland surface by simple remote sensing indices. ITM Web Conf. 2018, 23, 00030. [Google Scholar] [CrossRef]
- Van der Tol, C.; Vilfan, N.; Dauwe, D.; Cendrero-Mateo, M.P.; Yang, P.Q. The scattering and reabsorption of red and near-infrared chlorophyll fluorescence in the models Fluspect and SCOPE. Remote Sens. Environ. 2019, 232, 111292. [Google Scholar] [CrossRef]
- Pope, G.; Treitz, P. Leaf area index (LAI) estimation in boreal mixedwood forest of Ontario, Canada using light detection and ranging (LiDAR) and WorldView-2 imagery. Remote Sens. 2013, 5, 5040–5063. [Google Scholar] [CrossRef]
- Räsänen, A.; Juutinen, S.; Kalacska, M.; Aurela, M.; Heikkinen, P.; Mäenpää, K.; Rimali, A.; Virtanen, T. Peatland leaf-area index and biomass estimation with ultra-high resolution remote sensing. GIScience Remote Sens. 2020, 57, 943–964. [Google Scholar] [CrossRef]
- Antala, M.; Juszczak, R.; van der Tol, C.; Rastogi, A. Impact of climate change-induced alterations in peatland vegetation phenology and composition on carbon balance. Sci. Total Environ. 2022, 827, 154294. [Google Scholar] [CrossRef] [PubMed]
- Bandopadhyay, S.; Rastogi, A.; Cogliati, S.; Rascher, U.; Gąbka, M.; Juszczak, R. Can Vegetation Indices Serve as Proxies for Potential Sun-Induced Fluorescence (SIF)? A Fuzzy Simulation Approach on Airborne Imaging Spectroscopy Data. Remote Sens. 2021, 13, 2545. [Google Scholar] [CrossRef]
- Tesfaye, A.A.; Awoke, B.G. Evaluation of the saturation property of vegetation indices derived from sentinel-2 in mixed crop-forest ecosystem. Spat. Inf. Res. 2021, 29, 109–121. [Google Scholar] [CrossRef]
- Liu, L.; Yang, X.; Gong, F.; Su, Y.; Huang, G.; Chen, X. The Novel Microwave Temperature Vegetation Drought Index (MTVDI) Captures Canopy Seasonality across Amazonian Tropical Evergreen Forests. Remote Sens. 2021, 13, 339. [Google Scholar] [CrossRef]
- Camps-Valls, G.; Campos-Taberner, M.; Moreno-Martínez, A.; Walther, S.; Duveiller, G.; Cescatti, A.; Mahecha, M.D.; Muñoz-Marí, J.; García-Haro, F.J.; Guanter, L.; et al. A unified vegetation index for quantifying the terrestrial biosphere. Sci. Adv. 2021, 7, eabc7447. [Google Scholar] [CrossRef] [PubMed]
- Mishra, K.B.; Mishra, A.; Kubásek, J.; Urban, O.; Heyer, A.G. Low temperature induced modulation of photosynthetic induction in non-acclimated and cold-acclimated Arabidopsis thaliana: Chlorophyll a fluorescence and gas-exchange measurements. Photosynth. Res. 2019, 139, 123–143. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Indices | Formula | References |
|---|---|---|
| NDVI | [29] | |
| EVI | [30] | |
| NIRv | [9] |
| Parameters | Unit | Lower Boundary | Upper Boundary | Initial Value |
|---|---|---|---|---|
| Soil brightness (B) | - | 0 | 0.9 | 0.89 |
| Spectral shape latitude (lat) | - | 20 | 40 | 40 |
| Spectral shape longitude (lon) | - | 40 | 60 | 40 |
| Soil moisture volume percentage (smp) | % | 5 | 80 | 50 |
| Chlorophyll content (Cab) | µg cm−2 | 5 | 40 | 15 |
| Dry matter content (Cdm) | g cm−2 | 0.00 | 0.02 | 0.007 |
| Leaf water thickness equivalent (Cw) | cm | 0 | 0.2 | 0.080 |
| Senescent material (Cs) | fraction | 0 | 0.4 | 0.114 |
| Carotenoids content (Cca) | µg cm−2 | 0 | 25 | 8.381 |
| Anthocyanin content (Cant) | µg cm−2 | 0 | 40 | 1.4 |
| Leaf structure parameter (N) | - | 1 | 3.5 | 1.5 |
| Leaf area index (LAI) | m2 m−2 | 0.1 | 5 | 2 |
| Leaf inclination (lidfa) | - | −1 | 1 | −0.3 |
| Leaf inclination bimodality (lidfb) | - | −1 | 1 | −0.114 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rastogi, A.; Antala, M.; Prikaziuk, E.; Yang, P.; van der Tol, C.; Juszczak, R. Exploring the Potential of SCOPE Model for Detection of Leaf Area Index and Sun-Induced Fluorescence of Peatland Canopy. Remote Sens. 2022, 14, 4010. https://doi.org/10.3390/rs14164010
Rastogi A, Antala M, Prikaziuk E, Yang P, van der Tol C, Juszczak R. Exploring the Potential of SCOPE Model for Detection of Leaf Area Index and Sun-Induced Fluorescence of Peatland Canopy. Remote Sensing. 2022; 14(16):4010. https://doi.org/10.3390/rs14164010
Chicago/Turabian StyleRastogi, Anshu, Michal Antala, Egor Prikaziuk, Peiqi Yang, Christiaan van der Tol, and Radoslaw Juszczak. 2022. "Exploring the Potential of SCOPE Model for Detection of Leaf Area Index and Sun-Induced Fluorescence of Peatland Canopy" Remote Sensing 14, no. 16: 4010. https://doi.org/10.3390/rs14164010
APA StyleRastogi, A., Antala, M., Prikaziuk, E., Yang, P., van der Tol, C., & Juszczak, R. (2022). Exploring the Potential of SCOPE Model for Detection of Leaf Area Index and Sun-Induced Fluorescence of Peatland Canopy. Remote Sensing, 14(16), 4010. https://doi.org/10.3390/rs14164010