
Impacts of Extreme Precipitation and Diurnal Temperature Events on Grassland Productivity at Different Elevations on the Plateau

 ,
,  ,
, 
Abstract
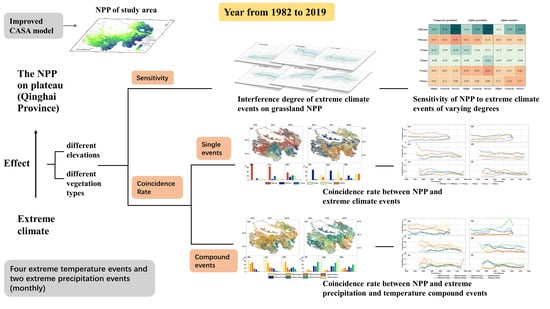
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Datasets
2.3. Methods
2.3.1. Identify Extreme Events
2.3.2. Events Coincidence Analysis
2.3.3. Sensitivity Analysis
3. Results
3.1. Distribution of NPP along Elevation Gradient
3.2. Sensitivity of Grasslands to Extreme Climate Events
3.3. The Coincidence Rate between Grasslands and Individual Extreme Climate Events
3.4. The Coincidence Rate between Grasslands and Compound Extreme Climate Events
3.5. Lag Analysis of Grassland Response to Extreme Climate Events
4. Discussion
4.1. Identification of Extreme Events
4.2. Grassland Response to Extreme Climate Events at Different Elevations
4.3. Response of Different Grasslands to Extreme Climate Events
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Stott, P. How Climate Change Affects Extreme Weather Events Research Can Increasingly Determine the Contribution of Climate Change to Extreme Events Such as Droughts. Science 2016, 352, 1517–1518. [Google Scholar] [CrossRef] [PubMed]
- Cleverly, J.; Eamus, D.; Edwards, W.; Grant, M.; Grundy, M.J.; Held, A.; Karan, M.; Lowe, A.J.; Prober, S.M.; Sparrow, B.; et al. TERN, Australia’s Land Observatory: Addressing the Global Challenge of Forecasting Ecosystem Responses to Climate Variability and Change. Environ. Res. Lett. 2019, 14, 095004. [Google Scholar] [CrossRef]
- Duane, A.; Castellnou, M.; Brotons, L. Towards a Comprehensive Look at Global Drivers of Novel Extreme Wildfire Events. Clim. Chang. 2021, 165, 43. [Google Scholar] [CrossRef]
- Hulme, P.E. Addressing the Threat to Biodiversity from Botanic Gardens. Trends Ecol. Evol. 2011, 26, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.C.; King, A.D.; Mitchell, D.M. Australia’s Unprecedented Future Temperature Extremes under Paris Limits to Warming. Geophys. Res. Lett. 2017, 44, 9947–9956. [Google Scholar] [CrossRef]
- Mishra, V.; Aadhar, S.; Asoka, A.; Pai, S.; Kumar, R. On the Frequency of the 2015 Monsoon Season Drought in the Indo-Gangetic Plain. Geophys. Res. Lett. 2016, 43, 12102–12112. [Google Scholar] [CrossRef]
- Chen, Q.; Timmermans, J.; Wen, W.; van Bodegom, P.M. Ecosystems Threatened by Intensified Drought with Divergent Vulnerability. Remote Sens. Environ. 2023, 289, 113512. [Google Scholar] [CrossRef]
- Katic, P.G.; Cerretelli, S.; Haggar, J.; Santika, T.; Walsh, C. Mainstreaming Biodiversity in Business Decisions: Taking Stock of Tools and Gaps. Biol. Conserv. 2023, 277, 109831. [Google Scholar] [CrossRef]
- Saleem, A.; Awange, J.L.; Kuhn, M.; John, B.; Hu, K. Impacts of Extreme Climate on Australia’s Green Cover (2003–2018): A MODIS and Mascon Probe. Sci. Total Environ. 2021, 766, 142567. [Google Scholar] [CrossRef]
- Fensholt, R.; Proud, S.R. Evaluation of Earth Observation Based Global Long Term Vegetation Trends—Comparing GIMMS and MODIS Global NDVI Time Series. Remote Sens. Environ. 2012, 119, 131–147. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, X.; Silander, J.A. Deciduous Forest Responses to Temperature, Precipitation, and Drought Imply Complex Climate Change Impacts. Proc. Natl. Acad. Sci. USA 2015, 112, 13585–13590. [Google Scholar] [CrossRef]
- Piao, S.; Zhang, X.; Chen, A.; Liu, Q.; Lian, X.; Wang, X.; Peng, S.; Wu, X. The Impacts of Climate Extremes on the Terrestrial Carbon Cycle: A Review. Sci. China Earth Sci. 2019, 62, 1551–1563. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, Y.; Shafeeque, M. Interpretation of Vegetation Phenology Changes Using Daytime and Night-Time Temperatures across the Yellow River Basin, China. Sci. Total Environ. 2019, 693, 133553. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Wang, X.; Xiao, J.; Zhang, K.; Wang, Y.; He, H.; Li, W.; Chen, H. Daytime and Nighttime Warming Has No Opposite Effects on Vegetation Phenology and Productivity in the Northern Hemisphere. Sci. Total Environ. 2022, 822, 153386. [Google Scholar] [CrossRef] [PubMed]
- Balducci, L.; Deslauriers, A.; Rossi, S.; Giovannelli, A. Stem Cycle Analyses Help Decipher the Nonlinear Response of Trees to Concurrent Warming and Drought. Ann. For. Sci. 2019, 76, 88. [Google Scholar] [CrossRef]
- Wen, Y.; Liu, X.; Pei, F.; Li, X.; Du, G. Non-Uniform Time-Lag Effects of Terrestrial Vegetation Responses to Asymmetric Warming. Agric. For. Meteorol. 2018, 252, 130–143. [Google Scholar] [CrossRef]
- Wen, Y.; Liu, X.; Yang, J.; Lin, K.; Du, G. NDVI Indicated Inter-Seasonal Non-Uniform Time-Lag Responses of Terrestrial Vegetation Growth to Daily Maximum and Minimum Temperature. Glob. Planet. Chang. 2019, 177, 27–38. [Google Scholar] [CrossRef]
- Zheng, L.; Li, D.; Xu, J.; Xia, Z.; Hao, H.; Chen, Z. A Twenty-Years Remote Sensing Study Reveals Changes to Alpine Pastures under Asymmetric Climate Warming. ISPRS J. Photogramm. Remote Sens. 2022, 190, 69–78. [Google Scholar] [CrossRef]
- Siegmund, J.F.; Wiedermann, M.; Donges, J.F.; Donner, R.V. Impact of Climate Extremes on Wildlife Plant Flowering over Germany. Biogeosci. Discuss. 2015, 12, 18389–18423. [Google Scholar]
- Baumbach, L.; Siegmund, J.F.; Mittermeier, M.; Donner, R.V. Impacts of Temperature Extremes on European Vegetation during the Growing Season. Biogeosciences 2017, 14, 4891–4903. [Google Scholar] [CrossRef]
- Fan, X.; Zhu, D.; Sun, X.; Wang, J.; Wang, M.; Wang, S.; Watson, A.E.E. Impacts of Extreme Temperature and Precipitation on Crops during the Growing Season in South Asia. Remote Sens. 2022, 14, 6093. [Google Scholar] [CrossRef]
- Tao, J.; Zhang, Y.; Dong, J.; Fu, Y.; Zhu, J.; Zhang, G.; Jiang, Y.; Tian, L.; Zhang, X.; Zhang, T.; et al. Elevation-Dependent Relationships between Climate Change and Grassland Vegetation Variation across the Qinghai-Xizang Plateau. Int. J. Climatol. 2015, 35, 1638–1647. [Google Scholar] [CrossRef]
- Zhao, R.; Chen, B.; Zhang, W.; Yang, S.; Xu, X. Moisture Source Anomalies Connected to Flood-Drought Changes over the Three-Rivers Headwater Region of Tibetan Plateau. Int. J. Climatol. 2023, 43, 5303–5316. [Google Scholar] [CrossRef]
- Donges, J.F.; Schleussner, C.-F.; Siegmund, J.F.; Donner, R.V. Event Coincidence Analysis for Quantifying Statistical Interrelationships between Event Time Series. Eur. Phys. J. Spec. Top. 2016, 225, 471–487. [Google Scholar] [CrossRef]
- Siegmund, J.F.; Siegmund, N.; Donner, R.V. CoinCalc—A New R Package for Quantifying Simultaneities of Event Series. Comput. Geosci. 2017, 98, 64–72. [Google Scholar] [CrossRef]
- You, Q.; Kang, S.; Aguilar, E.; Yan, Y. Changes in Daily Climate Extremes in the Eastern and Central Tibetan Plateau during 1961–2005. J. Geophys. Res. 2008, 113, D07101. [Google Scholar] [CrossRef]
- Liu, X.; Yin, Z.; Shao, X.; Qin, N. Temporal Trends and Variability of Daily Maximum and Minimum, Extreme Temperature Events, and Growing Season Length over the Eastern and Central Tibetan Plateau during 1961–2003. J. Geophys. Res. 2006, 111, D19109. [Google Scholar] [CrossRef]
- Qin, W.; Huang, Y.; Wang, L.; Lin, G.; Yang, J.; Song, P.; Gao, H.; Zhang, J.; Zhang, T. Gut Microbiota Enabled Goitered Gazelle (Gazella Subgutturosa) to Adapt to Seasonal Changes. Pak. J. Zool. 2020, 52, 1637–1646. [Google Scholar] [CrossRef]
- Frensch, J. Primary Responses of Root and Leaf Elongation to Water Deficits in the Atmosphere and Soil Solution. J. Exp. Bot. 1997, 48, 985–999. [Google Scholar] [CrossRef]
- He, C.; Davies, F.T.; Lacey, R.E. Separating the Effects of Hypobaria and Hypoxia on Lettuce: Growth and Gas Exchange. Physiol. Plant. 2007, 131, 226–240. [Google Scholar] [CrossRef]
- Kato, Y.; Abe, J.; Kamoshita, A.; Yamagishi, J. Genotypic Variation in Root Growth Angle in Rice (Oryza Sativa L.) and Its Association with Deep Root Development in Upland Fields with Different Water Regimes. Plant Soil 2006, 287, 117–129. [Google Scholar] [CrossRef]
- Turunen, M.; Latola, K. UV-B Radiation and Acclimation in Timberline Plants. Environ. Pollut. 2005, 137, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Lyu, M.; Sun, M.; Penuelas, J.; Sardans, J.; Sun, J.; Chen, X.; Zhong, Q.; Cheng, D. Thermal Acclimation of Foliar Carbon Metabolism in Pinus Taiwanensis along an Elevational Gradient. Front. Plant Sci. 2022, 12, 778045. [Google Scholar] [CrossRef]
- Tong, S.; Xu, D.; Wang, Y.; Zhang, X.; Li, Y.; Wu, H.; Ye, N. Influence of Ocean Acidification on Thermal Reaction Norms of Carbon Metabolism in the Marine Diatom Phaeodactylum Tricornutum. Mar. Environ. Res. 2021, 164, 105233. [Google Scholar] [CrossRef] [PubMed]
- Currier, C.M.; Sala, O.E. Precipitation versus Temperature as Phenology Controls in Drylands. Ecology 2022, 103, e3793. [Google Scholar] [CrossRef]
- Crozier, L. Warmer Winters Drive Butterfly Range Expansion by Increasing Survivorship. Ecology 2004, 85, 231–241. [Google Scholar] [CrossRef]
- Goulden, M.L.; Bales, R.C. Mountain Runoff Vulnerability to Increased Evapotranspiration with Vegetation Expansion. Proc. Natl. Acad. Sci. USA 2014, 111, 14071–14075. [Google Scholar] [CrossRef] [PubMed]
- Berg, A.; Sheffield, J. Evapotranspiration Partitioning in CMIP5 Models: Uncertainties and Future Projections. J. Clim. 2019, 32, 2653–2671. [Google Scholar] [CrossRef]
- Schwingshackl, C.; Davin, E.L.; Hirschi, M.; Sorland, S.L.; Wartenburger, R.; Seneviratne, S.I. Regional Climate Model Projections Underestimate Future Warming Due to Missing Plant Physiological CO2 Response. Environ. Res. Lett. 2019, 14, 114019. [Google Scholar] [CrossRef]
- Peng, S.; Piao, S.; Ciais, P.; Myneni, R.B.; Chen, A.; Chevallier, F.; Dolman, A.J.; Janssens, I.A.; Peñuelas, J.; Zhang, G.; et al. Asymmetric Effects of Daytime and Night-Time Warming on Northern Hemisphere Vegetation. Nature 2013, 501, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Xia, J.; Liu, W.; Niu, S. Photosynthetic Overcompensation under Nocturnal Warming Enhances Grassland Carbon Sequestration. Ecology 2009, 90, 2700–2710. [Google Scholar] [CrossRef]
- Chen, Z.; Yu, G.; Ge, J.; Sun, X.; Hirano, T.; Saigusa, N.; Wang, Q.; Zhu, X.; Zhang, Y.; Zhang, J.; et al. Temperature and Precipitation Control of the Spatial Variation of Terrestrial Ecosystem Carbon Exchange in the Asian Region. Agric. For. Meteorol. 2013, 182–183, 266–276. [Google Scholar] [CrossRef]
- Fu, Y.; Zheng, Z.; Yu, G.; Hu, Z.; Sun, X.; Shi, P.; Wang, Y.; Zhao, X. Environmental Influences on Carbon Dioxide Fluxes over Three Grassland Ecosystems in China. Biogeosciences 2009, 6, 2879–2893. [Google Scholar] [CrossRef]
- Li, H.; Wang, C.; Zhang, F.; He, Y.; Shi, P.; Guo, X.; Wang, J.; Zhang, L.; Li, Y.; Cao, G.; et al. Atmospheric Water Vapor and Soil Moisture Jointly Determine the Spatiotemporal Variations of CO2 Fluxes and Evapotranspiration across the Qinghai-Tibetan Plateau Grasslands. Sci. Total Environ. 2021, 791, 148379. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Li, W.; Chen, S.; Tan, X.; Wang, S.; Chen, M.; Ren, T.; Xia, J.; Huang, J.; Han, X. Changing Precipitation Exerts Greater Influence on Soil Heterotrophic than Autotrophic Respiration in a Semiarid Steppe. Agric. For. Meteorol. 2019, 271, 413–421. [Google Scholar] [CrossRef]
- Jiang, S.; Chen, X.; Huang, R.; Wang, T.; Smettem, K. Effect of the Altitudinal Climate Change on Growing Season Length for Deciduous Broadleaved Forest in Southwest China. Sci. Total Environ. 2022, 828, 154306. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Event Type | Basic Indicator | Interpretation of Event |
|---|---|---|
| NPPmin | NPP (month) | Months in which NPP value < −1 STD |
| NPPmax | Months in which NPP value > 1 STD | |
| PREmin | Precipitation (month) | Months in which Precipitation value < −1 STD |
| PREmax | Months in which Precipitation value > 1 STD | |
| TNmin | Monthly minimum nighttime temperature | Months in which minimum nighttime temperature value < −1 STD |
| TDmin | Monthly minimum daytime temperature | Months in which minimum daytime temperature value < −1 STD |
| TNmax | Monthly maximum nighttime temperature | Months in which maximum nighttime temperature value > 1 STD |
| TDmax | Monthly maximum daytime temperature | Months in which maximum daytime temperature value > 1 STD |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, H.; Zhai, J.; Song, X.; Wang, G.; Zhong, Y.; Zhang, K.; Sun, W. Impacts of Extreme Precipitation and Diurnal Temperature Events on Grassland Productivity at Different Elevations on the Plateau. Remote Sens. 2024, 16, 317. https://doi.org/10.3390/rs16020317
An H, Zhai J, Song X, Wang G, Zhong Y, Zhang K, Sun W. Impacts of Extreme Precipitation and Diurnal Temperature Events on Grassland Productivity at Different Elevations on the Plateau. Remote Sensing. 2024; 16(2):317. https://doi.org/10.3390/rs16020317
Chicago/Turabian StyleAn, Hexuan, Jun Zhai, Xiaoyan Song, Gang Wang, Yu Zhong, Ke Zhang, and Wenyi Sun. 2024. "Impacts of Extreme Precipitation and Diurnal Temperature Events on Grassland Productivity at Different Elevations on the Plateau" Remote Sensing 16, no. 2: 317. https://doi.org/10.3390/rs16020317
APA StyleAn, H., Zhai, J., Song, X., Wang, G., Zhong, Y., Zhang, K., & Sun, W. (2024). Impacts of Extreme Precipitation and Diurnal Temperature Events on Grassland Productivity at Different Elevations on the Plateau. Remote Sensing, 16(2), 317. https://doi.org/10.3390/rs16020317