Trait Estimation in Herbaceous Plant Assemblages from in situ Canopy Spectra



Abstract
:1. Introduction
2. Materials & Methods
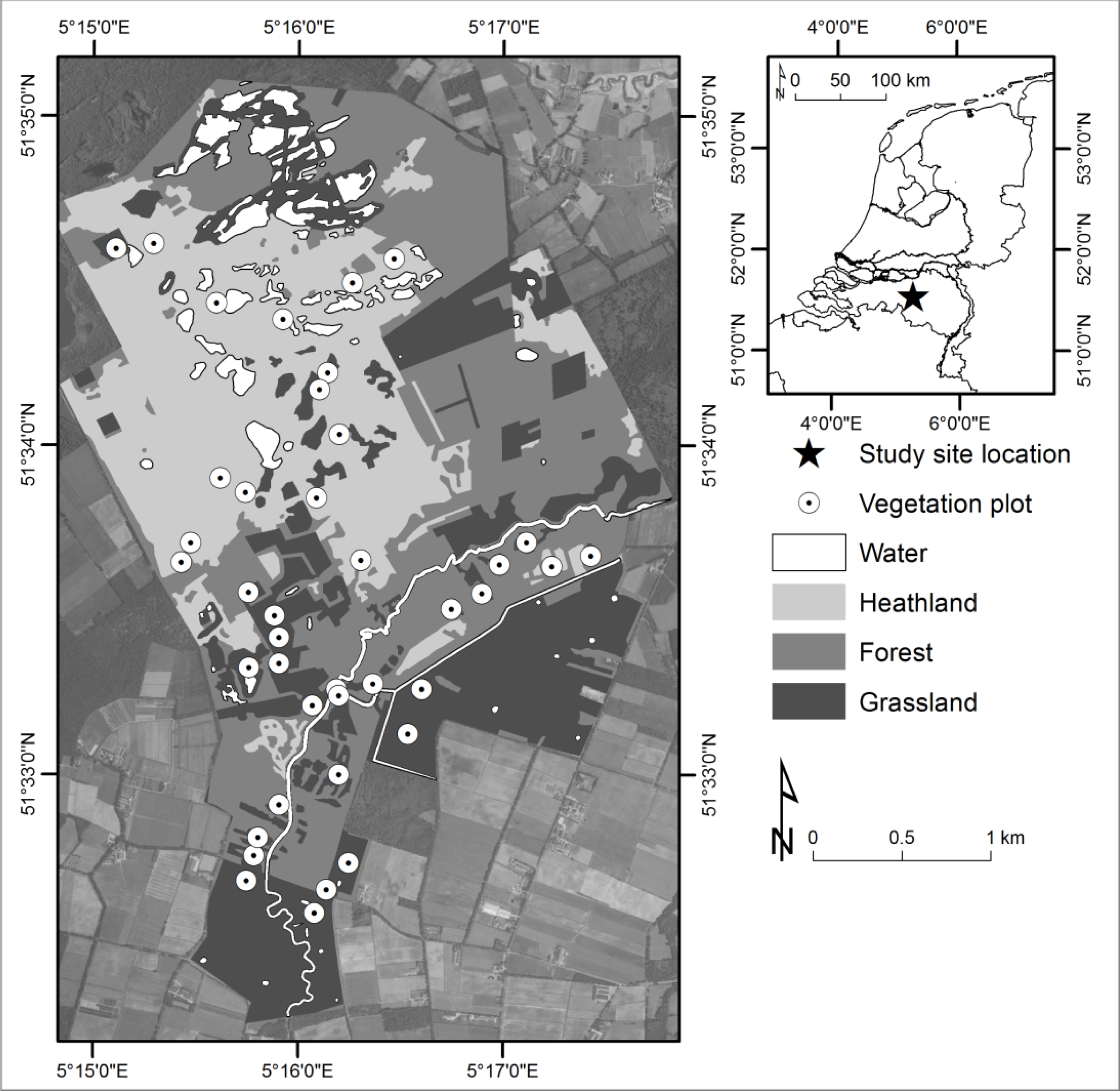
2.1. Study Site
2.2. Data Collection and Processing
2.2.1. Plot Surveying and Floristic Composition
- Dry heathlands, dominated by C. vulgaris, n = 4
- Moist heathlands, dominated by E. tetralix, n = 5
- Heathlands with grass encroachment, dominated by Molinia caerulea, n = 4
- Eutrophic grasslands, former meadows, among others species abundance of Trifolium sp, Holcus lanatus, Agrostis capillaris, n = 7
- Eutrophic meadows along walking paths, dominated by Phragmitum australis and Utrica dioica, n = 2.
- Mesotrophic wet grasslands, Cirsium dissectum, Succisa pratensis, n = 5.
- Eutrophic wet grasslands, Phalaris arundinacea, Carex acuta, n = 4.
- Mesotrophic, moist, sites within the recently converted agricultural areas, containing among others, Betula sp sapplings, Drosera intermedia, C. vulgaris and Salix repens sapplings, n = 5.
2.2.2. Plot Traits
2.2.3. Plot Co-Variates
2.2.4. Trait Expressions
2.2.5. Plot Spectra
2.3. Statistical Data Analysis
3. Results
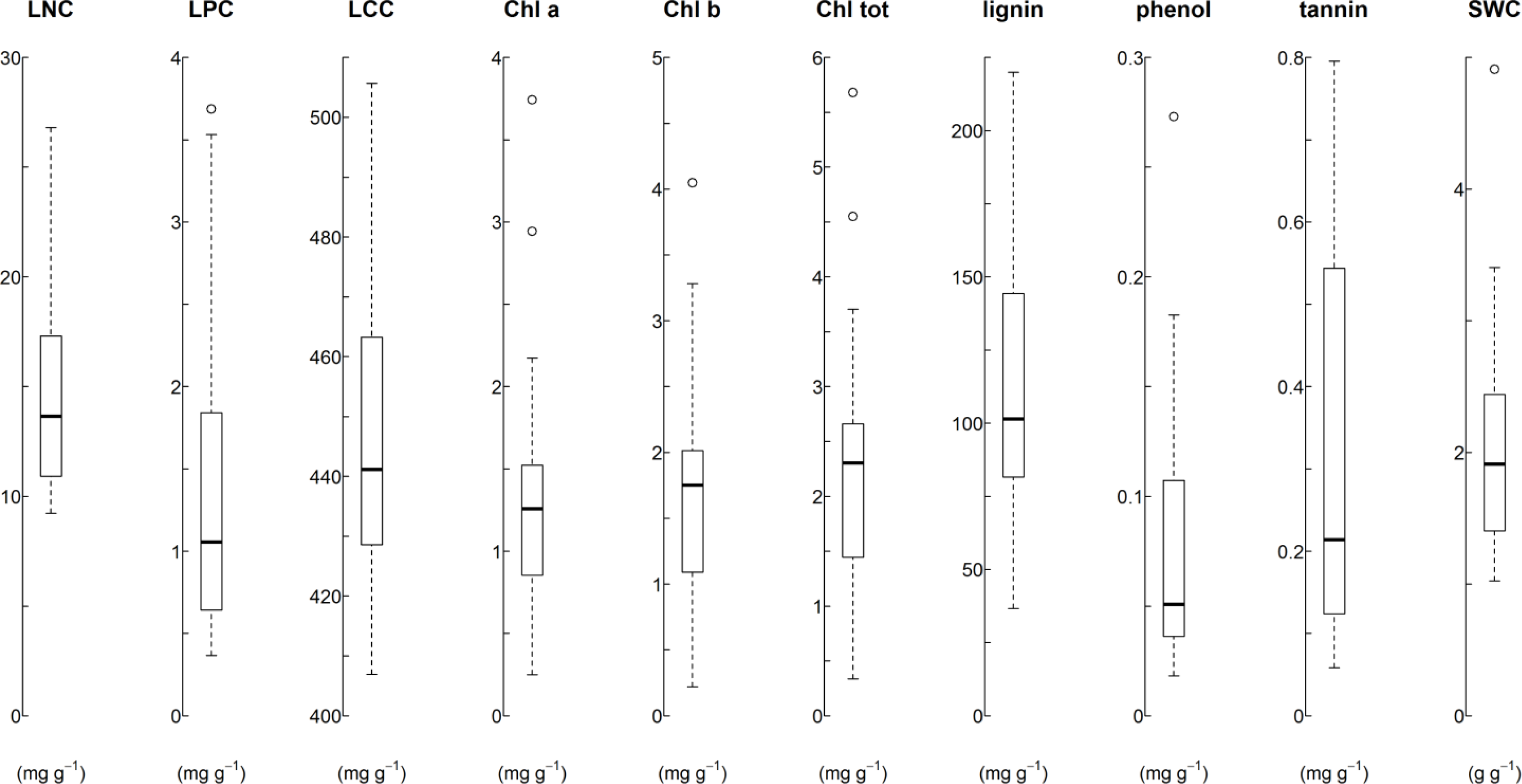
3.1. Traits Values and Plot Co-Variates Reflect Wide Range of Environmental Conditions
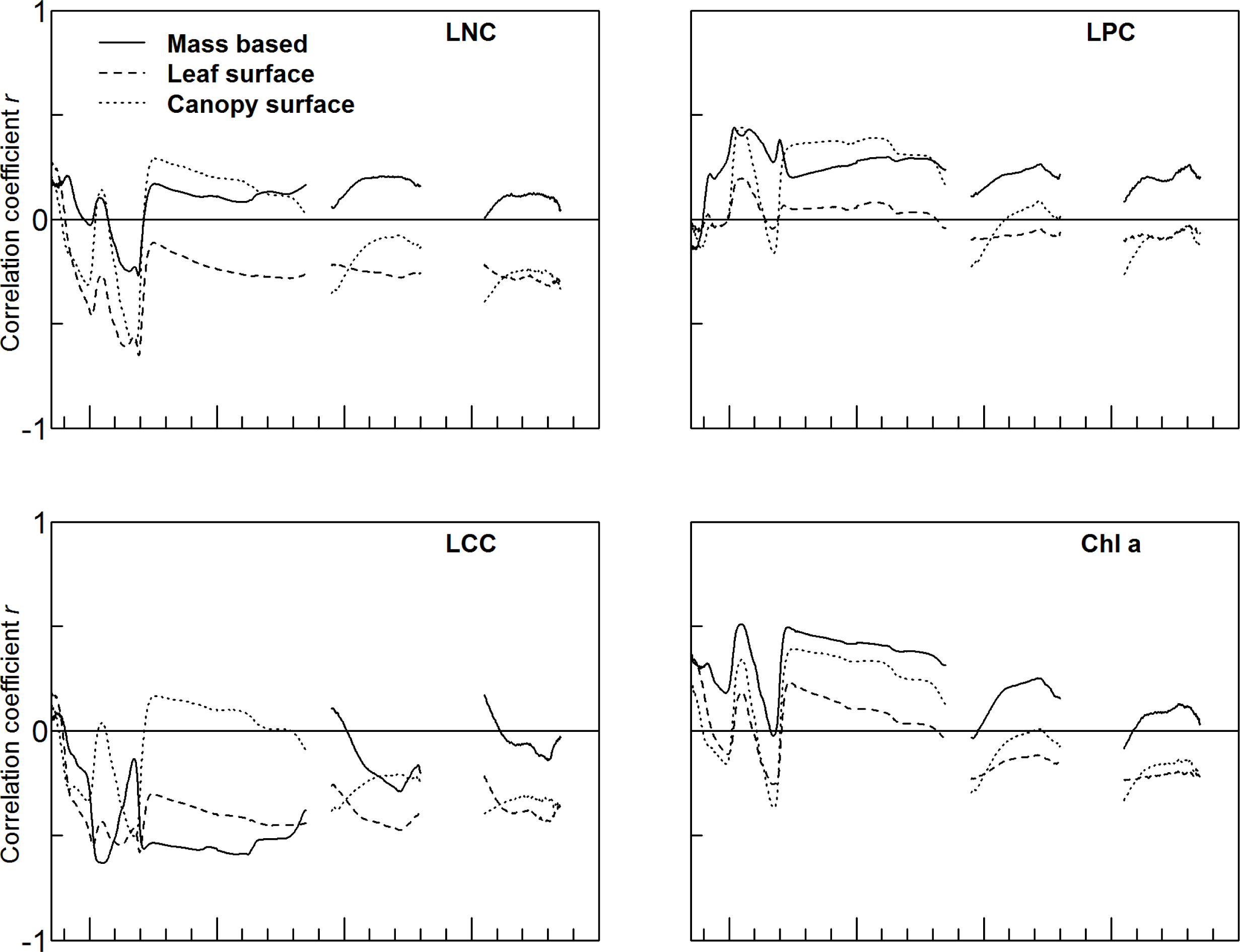
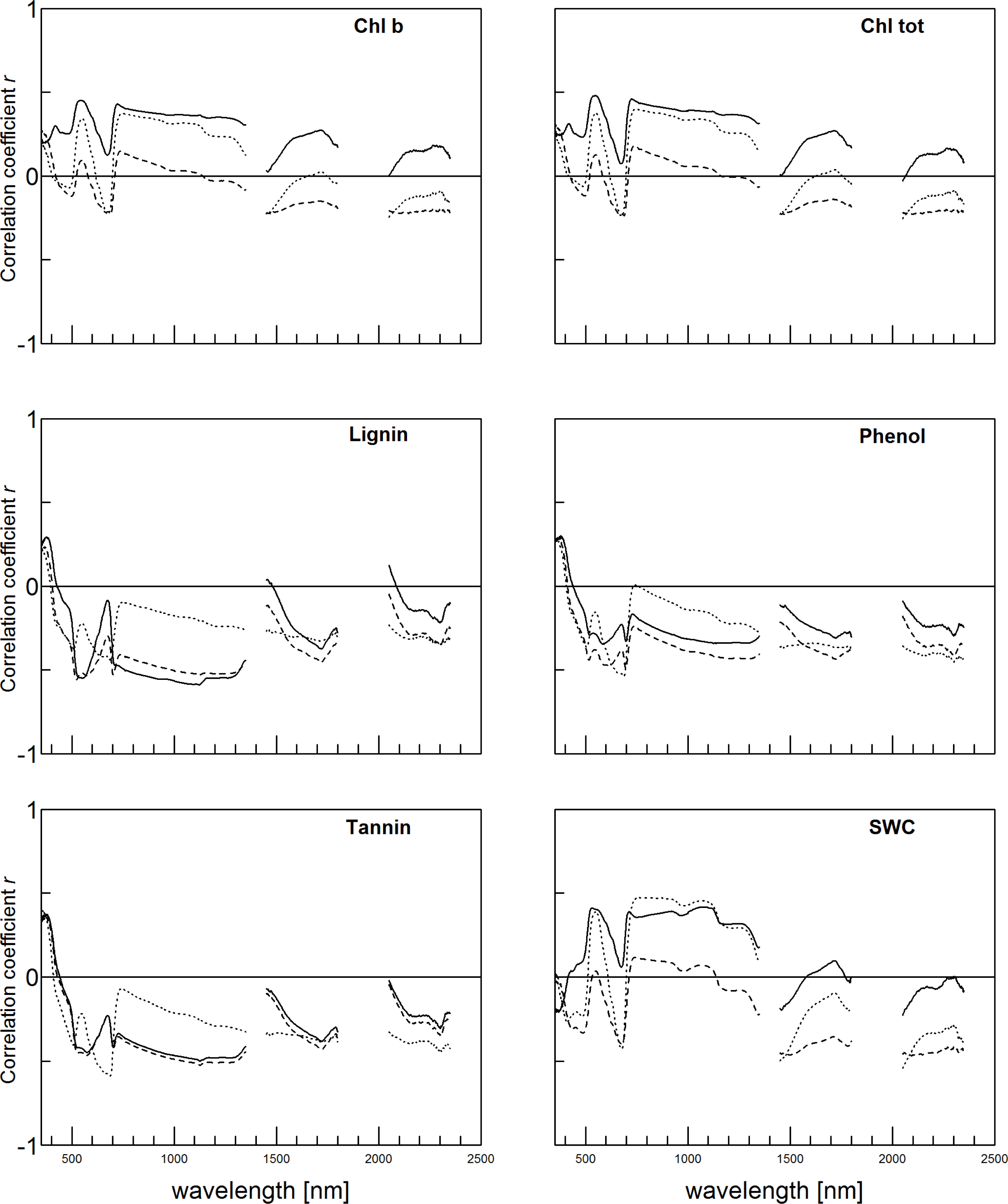
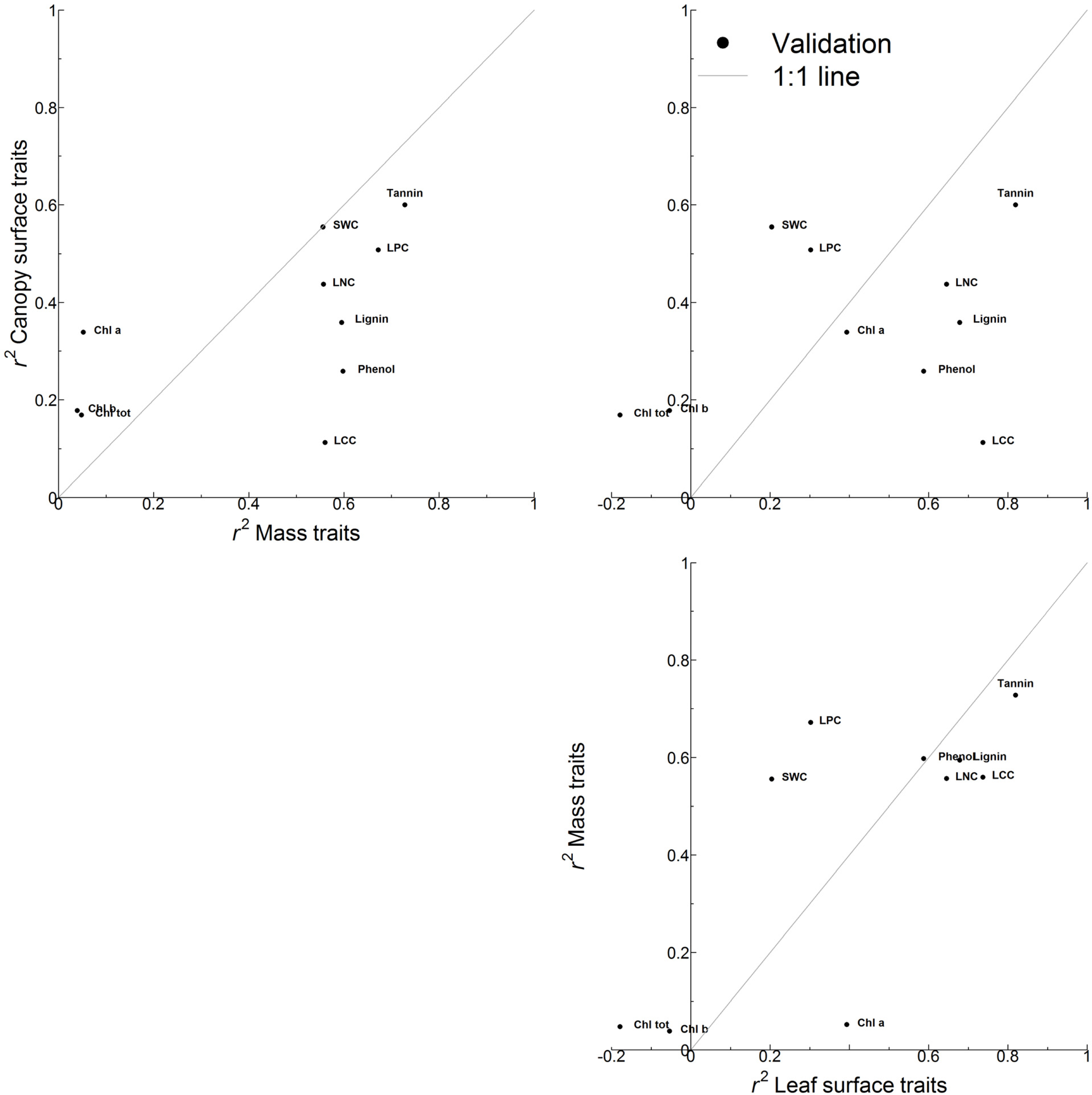
3.2. Accuracy of Trait Estimation Varies with Trait and Trait Expression
3.3. Plot Covariates May Be Additional Drivers of Canopy Reflectance
4. Discussion
4.1. Trait Estimation in Herbaceous Plant Assemblages Appears Feasible
4.2. Alternative Expressions of Trait Aggregation
4.3. Implications for Trait Estimations in Herbaceous Ecosystems
5. Conclusions
Supplementary Information
remotesensing-05-06323-s001.pdfAcknowledgments
Conflicts of Interest
References and Notes
- Lavorel, S.; Garnier, E. Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the holy grail. Funct. Ecol 2002, 16, 545–556. [Google Scholar]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot 2013, 61, 167–234. [Google Scholar]
- Svoray, T.; Perevolotsky, A.; Atkinson, P.M. Ecological sustainability in rangelands: The contribution of remote sensing. Int. J. Remote Sens 2013, 34, 6216–6242. [Google Scholar]
- Ustin, S.L. Remote sensing of canopy chemistry. Proc. Natl. Acad. Sci. USA 2013, 110, 804–805. [Google Scholar]
- Ramoelo, A.; Skidmore, A.K.; Cho, M.A.; Mathieu, R.; Heitkönig, I.M.A.; Dudeni-Tlhone, N.; Schlerf, M.; Prins, H.H.T. Non-linear partial least square regression increases the estimation accuracy of grass nitrogen and phosphorus using in situ hyperspectral and environmental data. ISPRS J. Photogramm. Remote Sens 2013, 82, 27–40. [Google Scholar]
- Curran, P.J.; Dungan, J.L.; Macler, B.A.; Plummer, S.E.; Peterson, D.L. Reflectance spectroscopy of fresh whole leaves for the estimation of chemical concentration. Remote Sens. Environ 1992, 39, 153–166. [Google Scholar]
- Doughty, C.E.; Asner, G.P.; Martin, R.E. Predicting tropical plant physiology from leaf and canopy spectroscopy. Oecologia 2011, 165, 289–299. [Google Scholar]
- Darvishzadeh, R.; Skidmore, A.; Schlerf, M.; Atzberger, C. Inversion of a radiative transfer model for estimating vegetation lai and chlorophyll in a heterogeneous grassland. Remote Sens. Environ 2007, 112, 2592–2604. [Google Scholar]
- Hansen, P.; Schjoerring, J. Reflectance measurement of canopy biomass and nitrogen status in wheat crops using normalized difference vegetation indices and partial least squares regression. Remote Sens. Environ 2003, 86, 542–553. [Google Scholar]
- Clevers, J.G.P.W.; Kooistra, L. Using hyperspectral remote sensing data for retrieving canopy chlorophyll and nitrogen content. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens 2012, 5, 574–583. [Google Scholar]
- Knox, N.M.; Skidmore, A.K.; Prins, H.H.; Asner, G.P.; van der Werff, H.; de Boer, W.F.; van der Waal, C.; de Knegt, H.J.; Kohi, E.M.; Slotow, R. Dry season mapping of savanna forage quality, using the hyperspectral carnegie airborne observatory sensor. Remote Sens. Environ 2011, 115, 1478–1488. [Google Scholar]
- Knyazikhin, Y.; Schull, M.A.; Stenberg, P.; Mõttus, M.; Rautiainen, M.; Yang, Y.; Marshak, A.; Carmona, P.L.; Kaufmann, R.K.; Lewis, P. Hyperspectral remote sensing of foliar nitrogen content. Proc. Natl. Acad. Sci. USA 2013, 110, E185–E192. [Google Scholar]
- Wallace, O.C.; Qi, J.; Heilma, P.; Marsett, R.C. Remote sensing for cover change assessment in southeast arizona. J. Range Manage 2003, 56, 402–409. [Google Scholar]
- Skidmore, A.K.; Ferwerda, J.G.; Mutanga, O.; Van Wieren, S.E.; Peel, M.; Grant, R.C.; Prins, H.H.; Balcik, F.B.; Venus, V. Forage quality of savannas—Simultaneously mapping foliar protein and polyphenols for trees and grass using hyperspectral imagery. Remote Sens. Environ 2010, 114, 64–72. [Google Scholar]
- Asner, G.P.; Martin, R.E. Canopy phylogenetic, chemical and spectral assembly in a lowland amazonian forest. New Phytol 2011, 189, 999–1012. [Google Scholar]
- Verrelst, J.; Geerling, G.W.; Sykora, K.V.; Clevers, J.G.P.W. Mapping of aggregated floodplain plant communities using image fusion of casi and lidar data. Int. J. Appl. Earth Obs. Geoinf 2009, 11, 83–94. [Google Scholar]
- Schmidt, K.S.; Skidmore, A.K.; Kloosterman, E.H.; Van Oosten, H.; Kumar, L.; Janssen, J.A.M. Mapping coastal vegetation using an expert system and hyperspectral imagery. Photogramm. Eng. Remote Sens 2004, 70, 703–715. [Google Scholar]
- Feilhauer, H.; Schmidtlein, S. On variable relations between vegetation patterns and canopy reflectance. Ecol. Inf 2011, 6, 83–92. [Google Scholar]
- Röder, A.; Kuemmerle, T.; Hill, J.; Papanastasis, V.; Tsiourlis, G. Adaptation of a grazing gradient concept to heterogeneous mediterranean rangelands using cost surface modelling. Ecol. Model 2007, 204, 387–398. [Google Scholar]
- Bello, F.; Thuiller, W.; Leps, J.; Choler, P.; Clement, J.C.; Macek, P.; Sebastia, M.T.; Lavorel, S. Partitioning of functional diversity reveals the scale and extent of trait convergence and divergence. J. Veg. Sci 2009, 20, 475–486. [Google Scholar]
- Darvishzadeh, R.; Skidmore, A.; Schlerf, M.; Atzberger, C.; Corsi, F.; Cho, M. LAI and chlorophyll estimation for a heterogeneous grassland using hyperspectral measurements. ISPRS J. Photogramm. Remote Sens 2008, 63, 409–426. [Google Scholar]
- Fava, F.; Colombo, R.; Bocchi, S.; Meroni, M.; Sitzia, M.; Fois, N.; Zucca, C. Identification of hyperspectral vegetation indices for mediterranean pasture characterization. Int. J. Appl. Earth Obs. Geoinf 2009, 11, 233–243. [Google Scholar]
- Mutanga, O.; Skidmore, A.K.; Prins, H. Predicting in situ pasture quality in the kruger national park, south africa, using continuum-removed absorption features. Remote Sens. Environ 2004, 89, 393–408. [Google Scholar]
- Ferwerda, J.G.; Skidmore, A.K. Can nutrient status of four woody plant species be predicted using field spectrometry? ISPRS J. Photogramm. Remote Sens 2007, 62, 406–414. [Google Scholar]
- Clevers, J.G.P.W.; Kooistra, L.; Schaepman, M.E. Estimating canopy water content using hyperspectral remote sensing data. Int. J. Appl. Earth Obs. Geoinf 2010, 12, 119–125. [Google Scholar]
- Verrelst, J.; Romijn, E.; Kooistra, L. Mapping vegetation density in a heterogeneous river floodplain ecosystem using pointable chris/proba data. Remote Sens 2012, 4, 2866–2889. [Google Scholar]
- Rivera, J.P.; Verrelst, J.; Leonenko, G.; Moreno, J. Multiple cost functions and regularization options for improved retrieval of leaf chlorophyll content and lai through inversion of the prosail model. Remote Sens 2013, 5, 3280–3304. [Google Scholar]
- Dorigo, W.; Richter, R.; Baret, F.; Bamler, R.; Wagner, W. Enhanced automated canopy characterization from hyperspectral data by a novel two step radiative transfer model inversion approach. Remote Sens 2009, 1, 1139–1170. [Google Scholar]
- Poorter, H.; Lambers, H.; Evans, J.R. Trait correlation networks: A whole-plant perspective on the recently criticized leaf economic spectrum. New Phytol. 2013. [Google Scholar] [CrossRef]
- Grossman, Y.; Ustin, S.; Jacquemoud, S.; Sanderson, E.; Schmuck, G.; Verdebout, J. Critique of stepwise multiple linear regression for the extraction of leaf biochemistry information from leaf reflectance data. Remote Sens. Environ 1996, 56, 182–193. [Google Scholar]
- Mirik, M.; Norland, J.E.; Crabtree, R.L.; Biondini, M.E. Hyperspectral one-meter-resolution remote sensing in yellowstone national park, wyoming: I. Forage nutritional values. Rangeland Ecol. Manage 2005, 58, 452–458. [Google Scholar]
- Pimstein, A.; Karnieli, A.; Bansal, S.K.; Bonfil, D.J. Exploring remotely sensed technologies for monitoring wheat potassium and phosphorus using field spectroscopy. Field Crops Res 2011, 121, 125–135. [Google Scholar]
- Aptroot, A. Flora- en Vegetatiekartering van Kampina in 2009; Natuurmonumenten: ‘s-Graveland, The Netherlands, 2009; p. 41. [Google Scholar]
- Murphy, J.; Riley, J. A modified single solution method for the determination of phosphate in natural waters. Anal. Chimica Acta 1962, 27, 31–36. [Google Scholar]
- Poorter, H.; Villar, R. The Fate of Acquired Carbon in Plants: Chemical Composition and Construction Costs. In Plant Resource Allocation; Academic Press: San Diego, CA, USA, 1997; pp. 39–72. [Google Scholar]
- Makkar, H.P.S. Quantification of Tannins in Tree and Shrub Foliage: A Laboratory Manual; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003. [Google Scholar]
- Porra, R.; Thompson, W.; Kriedemann, P. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochimica et Biophysica Acta (BBA)-Bioenergetics 1989, 975, 384–394. [Google Scholar]
- Diekmann, M. Species indicator values as an important tool in applied plant ecology—A review. Basic Appl. Ecol 2002, 4, 493–506. [Google Scholar]
- Witte, J.P.M.; Wójcik, R.B.; Torfs, P.J.J.F.; De Haan, M.W.H.; Hennekens, S. Bayesian classification of vegetation types with gaussian mixture density fitting to indicator values. J. Veg. Sci 2007, 18, 605–612. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Witrth, V.; Werner, W.; Paulißen, D. Zeigerwerte von pflanzen in mitteleuropa. Scripta Geobotanica 1991, 18, 1–97. [Google Scholar]
- Roelofsen, H.D.; Kooistra, L.; van Bodegom, P.M.; Verrelst, J.; Krol, J.; Witte, J.P.M. Mapping a-priori defined plant associations using remotely sensed vegetation characteristics. Remote Sens. Environ 2014, 140, 639–651. [Google Scholar]
- Zelený, D.; Schaffers, A.P. Too good to be true: Pitfalls of using mean ellenberg indicator values in vegetation analyses. J. Veg. Sci 2012, 23, 419–431. [Google Scholar]
- Welles, J.; Norman, J. Instrument for indirect measurement of canopy architecture. Agron. J 1991, 83, 818–825. [Google Scholar]
- Hunt, E.R., Jr.; Rock, B.N. Detection of changes in leaf water content using near- and middle-infrared reflectances. Remote Sens. Environ 1989, 30, 43–54. [Google Scholar]
- Morisette, J.T.; Baret, F.; Privette, J.L.; Myneni, R.B.; Nickeson, J.E.; Garrigues, S.; Shabanov, N.V.; Weiss, M.; Fernandes, R.A.; Leblanc, S.G. Validation of global moderate-resolution lai products: A framework proposed within the ceos land product validation subgroup. IEEE Trans. Geosci. Remote Sens 2006, 44, 1804–1817. [Google Scholar]
- Savitzky, A.; Golay, M.J. Smoothing and differentiation of data by simplified least squares procedures. Anal. Chem 1964, 36, 1627–1639. [Google Scholar]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemometr. Intell. Lab. Syst 2001, 58, 109–130. [Google Scholar]
- R_Core_Team. R: A Language and Environment for Statistical Computing; R foundation for statistical computing: Vienna, Austria, 2013. [Google Scholar]
- Mevik, B.H.; Wehrens, R. The PLS package: Principal component and partial least squares regression in r. J. Statist. Softw 2007, 18, 1–23. [Google Scholar]
- Feilhauer, H.; Asner, G.P.; Martin, R.E.; Schmidtlein, S. Brightness-normalized partial least squares regression for hyperspectral data. J. Quant. Spectrosc. Radiat. Transfer 2010, 111, 1947–1957. [Google Scholar]
- Ustin, S.L.; Gitelson, A.A.; Jacquemoud, S.; Schaepman, M.; Asner, G.P.; Gamon, J.A.; Zarco-Tejada, P. Retrieval of foliar information about plant pigment systems from high resolution spectroscopy. Remote Sens. Environ 2009, 113, S67–S77. [Google Scholar]
- Jacquemoud, S.; Baret, F. Prospect: A model of leaf optical properties spectra. Remote Sens. Environ 1990, 34, 75–91. [Google Scholar]
- Daughtry, C.S.T.; Walthall, C.L.; Kim, M.S.; De Colstoun, E.B.; McMurtrey, J.E., III. Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance. Remote Sens. Environ 2000, 74, 229–239. [Google Scholar]
- Cho, M.A.; Skidmore, A.K. A new technique for extracting the red edge position from hyperspectral data: The linear extrapolation method. Remote Sens. Environ 2006, 101, 181–193. [Google Scholar]
- Im, J.; Jensen, J.R.; Jensen, R.R.; Gladden, J.; Waugh, J.; Serrato, M. Vegetation cover analysis of hazardous waste sites in utah and arizona using hyperspectral remote sensing. Remote Sens 2012, 4, 327–353. [Google Scholar]
- Freschet, G.T.; Cornelissen, J.H.; Van Logtestijn, R.S.; Aerts, R. Evidence of the ‘plant economics spectrum’ in a subarctic flora. J. Ecol 2010, 98, 362–373. [Google Scholar]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornellssen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar]
- Chuvieco, E.; Aguado, I.; Yebra, M.; Nieto, H.; Salas, J.; Martín, M.P.; Vilar, L.; Martínez, J.; Martín, S.; Ibarra, P. Development of a framework for fire risk assessment using remote sensing and geographic information system technologies. Ecol. Model 2010, 221, 46–58. [Google Scholar]
- Sow, M.; Mbow, C.; Hély, C.; Fensholt, R.; Sambou, B. Estimation of herbaceous fuel moisture content using vegetation indices and land surface temperature from modis data. Remote Sens 2013, 5, 2617–2638. [Google Scholar]
- Laughlin, D.C.; Joshi, C.; Bodegom, P.M.; Bastow, Z.A.; Fulé, P.Z. A predictive model of community assembly that incorporates intraspecific trait variation. Ecol. Lett 2012, 15, 1291–1299. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acidity IV | Nutrient IV | Moisture IV | Height | Coverage | Bare soil | LAI | |
|---|---|---|---|---|---|---|---|
| LNCmass | −0.37 | ||||||
| LCCmass | −0.37 | ||||||
| Ligninmass | 0.31 | −0.33 | |||||
| SWCmass | −0.40 | 0.35 | −0.5 | ||||
| LNCleaf | 0.33 | ||||||
| LigninLeaf | 0.40 | ||||||
| PhenolLeaf | −0.32 | ||||||
| SWCleaf | −0.4 | ||||||
| LNCCanopy | 0.49 | 0.47 | |||||
| LCCCanopy | 0.34 | 0.53 | |||||
| LigninCanopy | 0.49 | 0.47 | −0.38 | 0.44 | |||
| Phenolcanopy | 0.31 | ||||||
| SWCcanopy | −0.4 | 0.36 | 0.48 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Roelofsen, H.D.; Van Bodegom, P.M.; Kooistra, L.; Witte, J.-P.M. Trait Estimation in Herbaceous Plant Assemblages from in situ Canopy Spectra. Remote Sens. 2013, 5, 6323-6345. https://doi.org/10.3390/rs5126323
Roelofsen HD, Van Bodegom PM, Kooistra L, Witte J-PM. Trait Estimation in Herbaceous Plant Assemblages from in situ Canopy Spectra. Remote Sensing. 2013; 5(12):6323-6345. https://doi.org/10.3390/rs5126323
Chicago/Turabian StyleRoelofsen, Hans D., Peter M. Van Bodegom, Lammert Kooistra, and Jan-Philip M. Witte. 2013. "Trait Estimation in Herbaceous Plant Assemblages from in situ Canopy Spectra" Remote Sensing 5, no. 12: 6323-6345. https://doi.org/10.3390/rs5126323
APA StyleRoelofsen, H. D., Van Bodegom, P. M., Kooistra, L., & Witte, J. -P. M. (2013). Trait Estimation in Herbaceous Plant Assemblages from in situ Canopy Spectra. Remote Sensing, 5(12), 6323-6345. https://doi.org/10.3390/rs5126323