Spatio-Temporal Patterns and Climate Variables Controlling of Biomass Carbon Stock of Global Grassland Ecosystems from 1982 to 2006

 ,
,
Abstract
:1. Introduction
2. Methods and Data
2.1. Biomass Data Collection
2.2. NDVI and Climate Dataset
2.3. Biomass Carbon Density Model Development
2.4. Mapping the Spatial and Temporal Dynamics of Grassland Biomass Worldwide
2.5. Correlation and Trend Analysis
3. Results
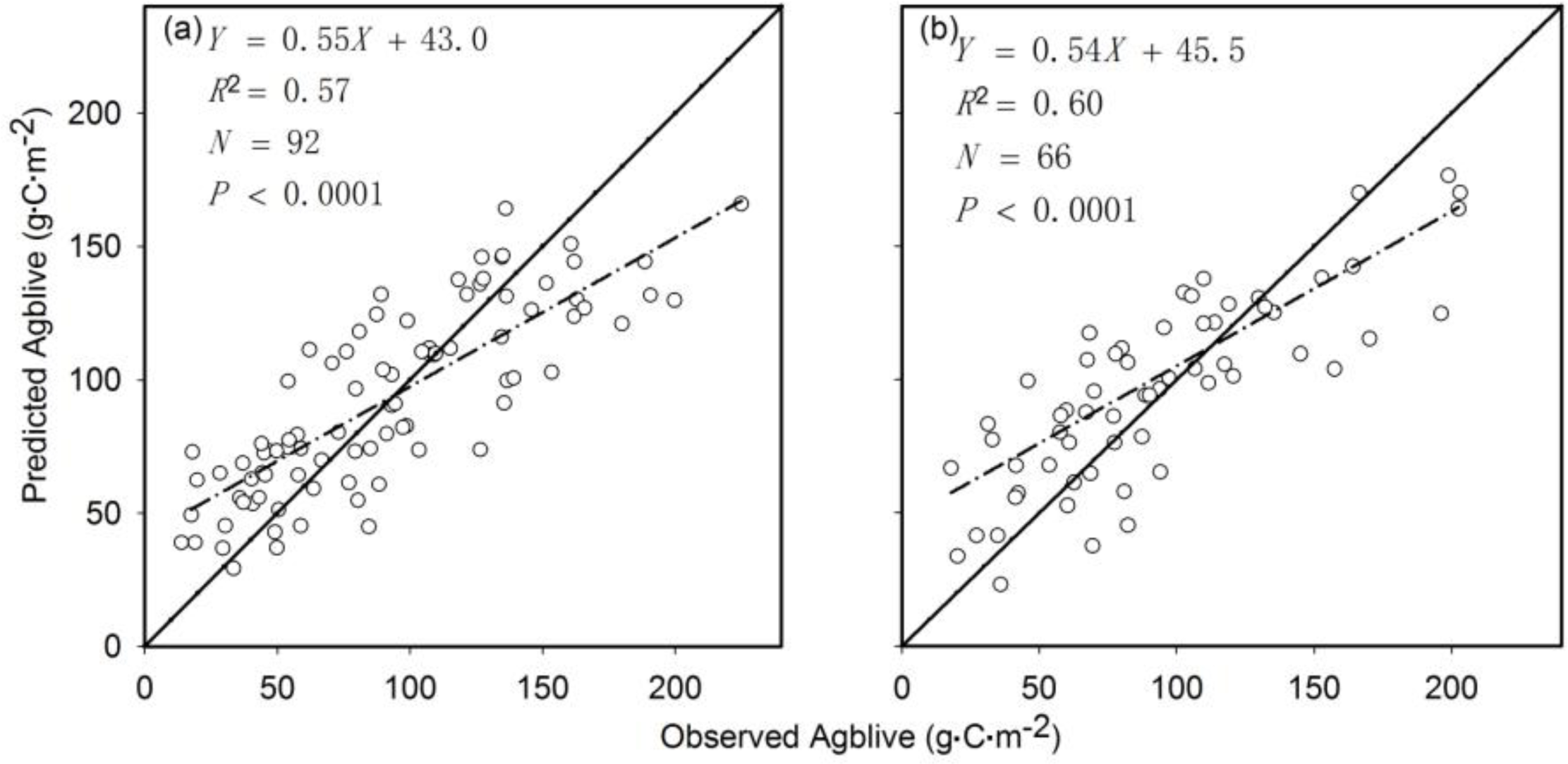
3.1. The Aboveground Live Biomass Model for Grassland Worldwide
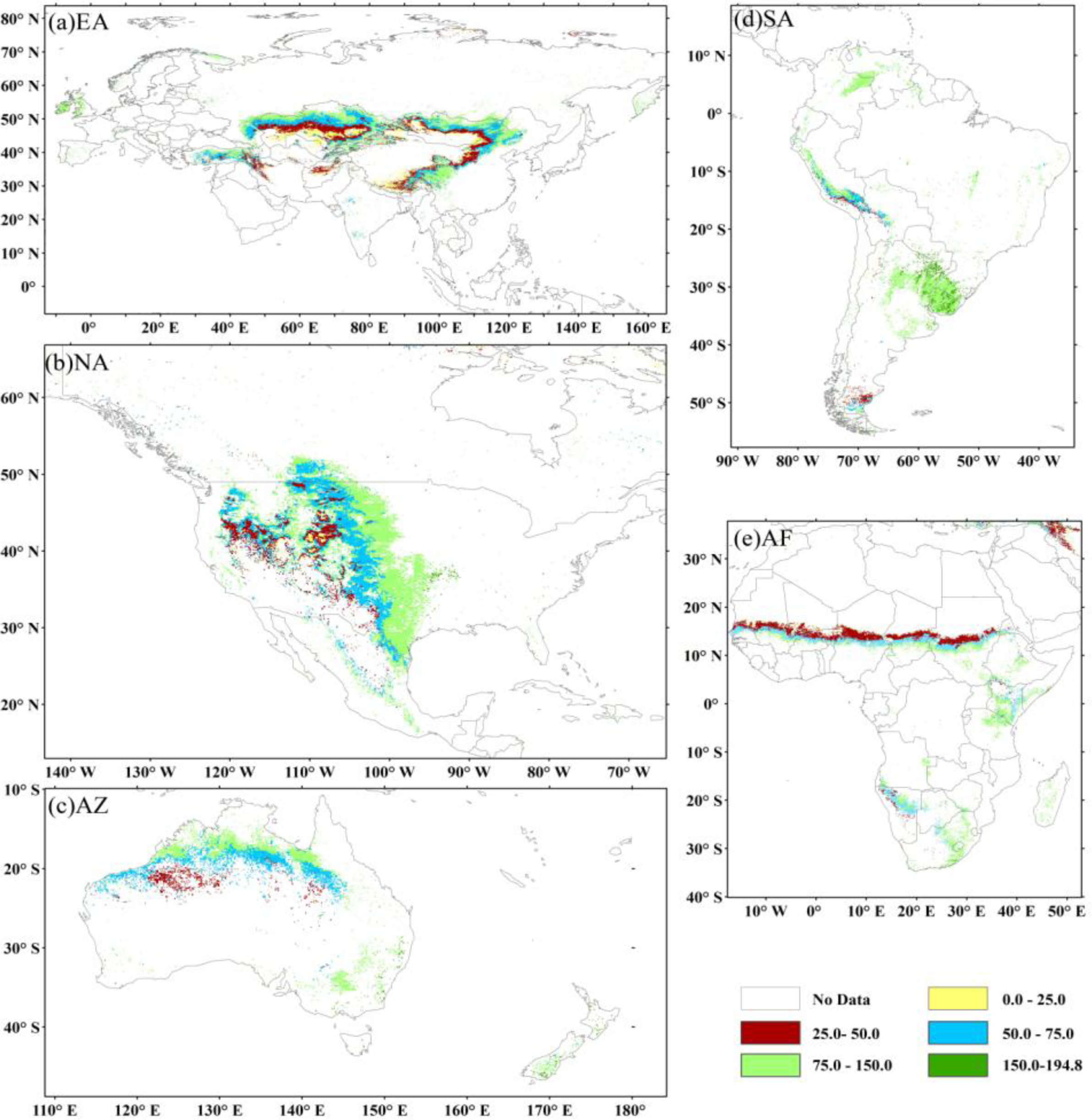
3.2. Global Biomass Carbon Stock and its Spatial Pattern
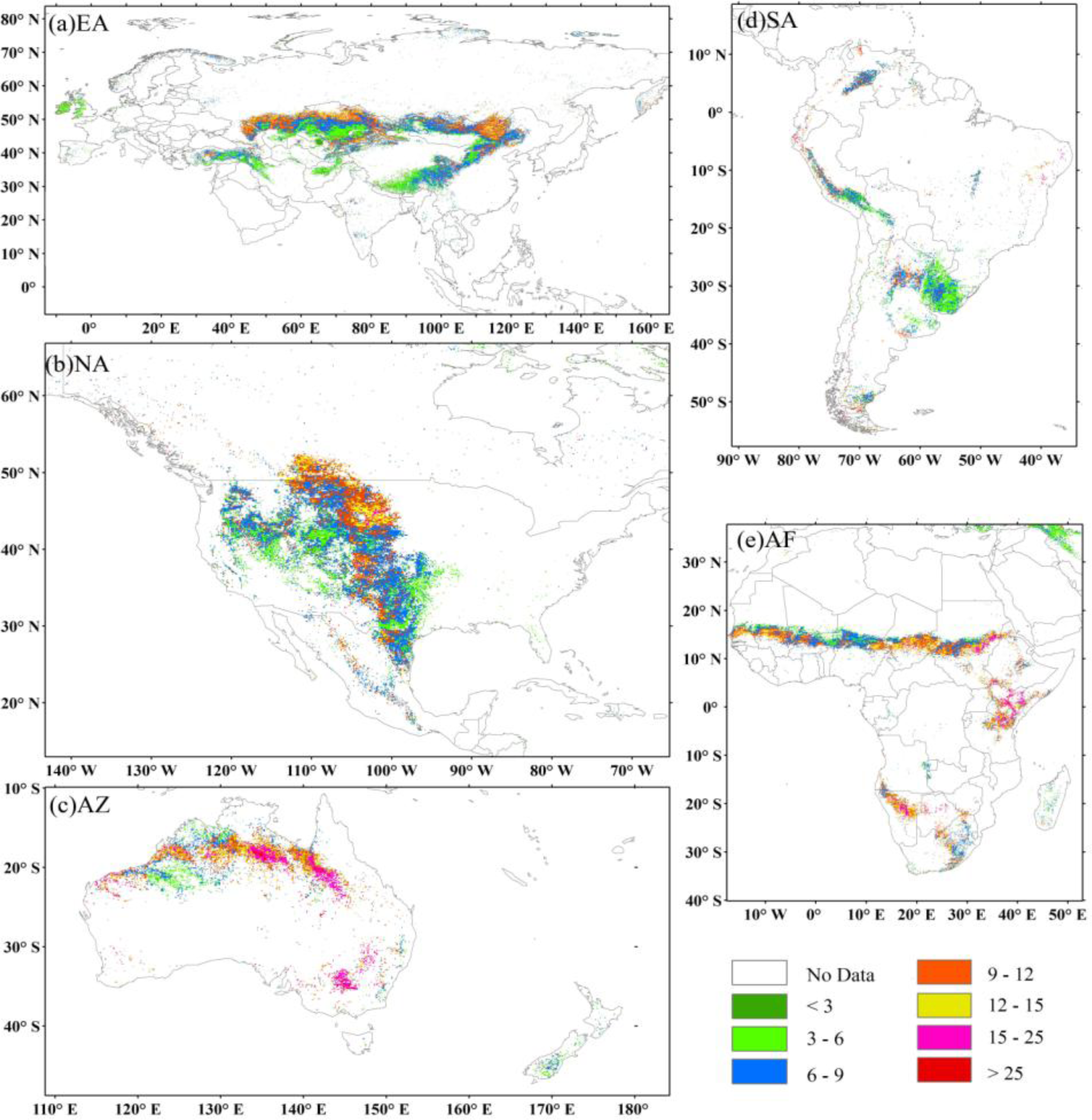
3.3. Temporal Variability of Biomass Carbon Stock
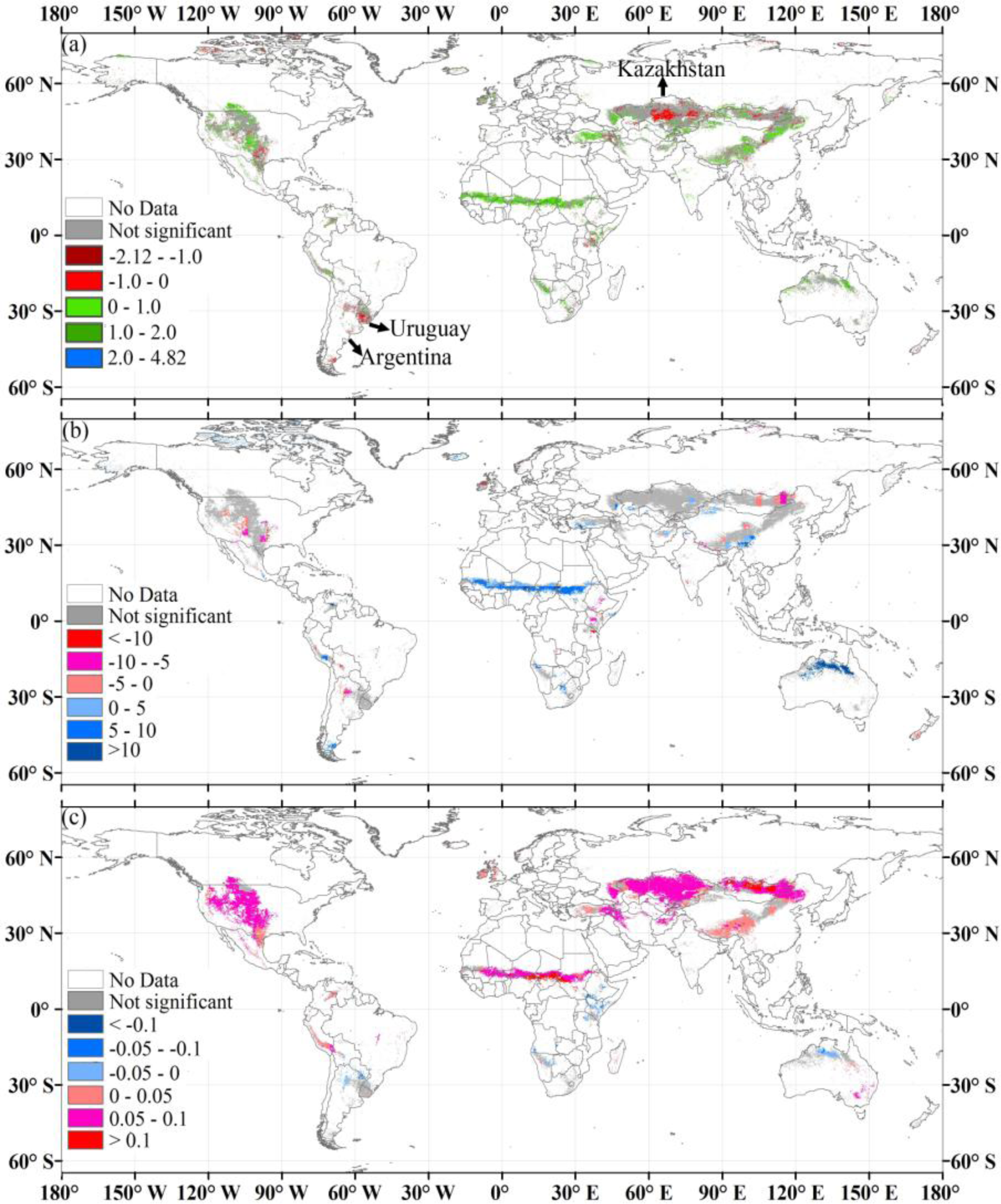
3.4. Temporal Trends of Biomass Carbon and Climate Change at Continental and Global Scales
3.5. Biomass, Climate, and Their Correlations at the Pixel Level
4. Discussion
4.1. Evaluation of the Biomass Carbon Density Model
4.2. Comparison of Biomass Estimates
4.3. Controlling Factors of Biomass Carbon Change
4.4. Uncertainties
5. Summary
remotesensing-06-01783-s001.pdf
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hall, D.O.; Scurlock, J.M.O. Climate change and productivity of natural grasslands. Ann. Bot 1991, 67, 49–55. [Google Scholar]
- Scurlock, J.M.O; Hall, D.O. The global carbon sink: A grassland perspective. Glob. Chang. Biol 1998, 4, 229–233. [Google Scholar]
- Ma, W.; Fang, J.; Yang, Y.; Mohammat, A. Biomass carbon stocks and their changes in northern China’s grasslands during 1982–2006. Sci. China Life Sci 2010, 53, 841–850. [Google Scholar]
- Canadell, J.G.; Le Quéré, C.; Raupach, M.R.; Field, C.B.; Buitenhuis, E.T.; Ciais, P.; Conway, T.J.; Gillett, N.P.; Houghton, R.A.; Marland, G. Contributions to accelerating atmospheric CO2 growth from economic activity, carbon intensity, and efficiency of natural sinks. Proc. Natl. Acad. Sci. USA 2007, 104, 18866–18870. [Google Scholar]
- Kruska, R.L.; Reid, R.S.; Thornton, P.K.; Henninger, N.; Kristjanson, P.M. Mapping livestock-oriented agricultural production systems for the developing world. Agric. Syst 2003, 77, 39–63. [Google Scholar]
- Thornton, P.K.; van de Steeg, J.; Notenbaert, A.; Herrero, M. The impacts of climate change on livestock and livestock systems in developing countries: A review of what we know and what we need to know. Agric. Syst 2009, 101, 113–127. [Google Scholar]
- Sautier, M.; Martin-Clouaire, R.; Faivre, R.; Duru, M. Assessing climatic exposure of grassland-based livestock systems with seasonal-scale indicators. Clim. Chang 2013, 120, 1–15. [Google Scholar]
- Jones, M.B.; Donnelly, A. Carbon sequestration in temperate grassland ecosystems and the influence of management, climate and elevated CO2. New Phytol 2004, 164, 423–439. [Google Scholar]
- Sala, O.E.; Gherardi, L.A.; Reichmann, L.; Jobbágy, E.; Peters, D. Legacies of precipitation fluctuations on primary production: Theory and data synthesis. Philos. Trans. R. Soc. B. Biol. Sci 2012, 367, 3135–3144. [Google Scholar]
- Reichmann, L.G.; Sala, O.E.; Peters, D.P.C. Precipitation legacies in desert grassland primary production occur through previous-year tiller density. Ecology 2013, 94, 435–443. [Google Scholar]
- Tucker, C.J.; Vanpraet, C.L.; Sharman, M.J.; van Ittersum, G. Satellite remote sensing of total herbaceous biomass production in the Senegalese Sahel: 1980–1984. Remote Sens. Environ 1985, 17, 233–249. [Google Scholar]
- Wylie, B.K.; Meyer, D.J.; Tieszen, L.L.; Mannel, S. Satellite mapping of surface biophysical parameters at the biome scale over the North American grasslands: A case study. Remote Sens. Environ 2002, 79, 266–278. [Google Scholar]
- Asner, G.P.; Nepstad, D.; Cardinot, G.; Ray, D. Drought stress and carbon uptake in an Amazon forest measured with spaceborne imaging spectroscopy. Proc. Natl. Acad. Sci. USA 2004, 101, 6039–6044. [Google Scholar]
- Piao, S.; Fang, J.; Zhou, L.; Tan, K.; Tao, S. Changes in biomass carbon stocks in China’s grasslands between 1982 and 1999. Glob. Biogeochem. Cycles 2007. [Google Scholar] [CrossRef]
- Yang, Y.; Fang, J.; Ma, W.; Guo, D.; Mohammat, A. Large-scale pattern of biomass partitioning across China’s grasslands. Glob. Ecol. Biogeogr 2010, 19, 268–277. [Google Scholar]
- Ferreira, L.G.; Fernandez, L.E.; Sano, E.E.; Field, C.; Sousa, S.B.; Arantes, A.E.; Araújo, F.M. Biophysical properties of cultivated pastures in the Brazilian savanna biome: An analysis in the spatial-temporal domains based on ground and satellite data. Remote Sens 2013, 5, 307–326. [Google Scholar]
- Sala, O.E.; Parton, W.J.; Joyce, L.A; Lauenroth, W.K. Primary production of the central grassland region of the United States. Ecology 1988, 69, 40–45. [Google Scholar]
- McNaughton, S.J.; Sala, O.E.; Oesterheld, M. Comparative Ecology of African and South American Arid to Subhumid Ecosystems. In Biological Relationships between Africa and South America; Goldblatt, P., Ed.; Yale University Press: New Haven, CT, USA, 1993; pp. 548–567. [Google Scholar]
- Briggs, J.M.; Knapp, A.K. Interannual variability in primary production in tallgrass prairie: Climate, soil moisture, topographic position, and fire as determinants of aboveground biomass. Am. J. Bot 1995, 1024–1030. [Google Scholar]
- Bai, Y.; Wu, J.; Xing, Q.; Pan, Q.; Huang, J.; Yang, D.; Han, X. Primary production and rain use efficiency across a precipitation gradient on the Mongolia plateau. Ecology 2008, 89, 2140–2153. [Google Scholar]
- Lauenroth, W.K.; Sala, O.E. Long-term forage production of North American shortgrass steppe. Ecol. Appl 1992, 2, 397–403. [Google Scholar]
- Huxman, T.E.; Smith, M.D.; Fay, P.A.; Knapp, A.K.; Shaw, M.R.; Loik, M.E.; Smith, S.D.; Tissue, D.T.; Zak, J.C.; Weltzin, J.F.; et al. Convergence across biomes to a common rain-use efficiency. Nature 2004, 429, 651–654. [Google Scholar]
- Hu, Z.; Fan, J.; Zhong, H.; Yu, G. Spatio-temporal dynamics of aboveground primary productivity along a precipitation gradient in Chinese temperate grassland. Sci. China Ser. D: Earth Sci 2007, 50, 754–764. [Google Scholar]
- Herrmann, S.M.; Anyamba, A.; Tucker, C.J. Recent trends in vegetation dynamics in the African Sahel and their relationship to climate. Glob. Environ. Chang 2005, 15, 394–404. [Google Scholar]
- Scurlock, J.M.O.; Johnson, K.R.; Olson, R.J. NPP Grassland: NPP Estimates from Biomass Dynamics for 31 Sites, 1948–1994; Oak Ridge National Laboratory Distributed Active Archive Center: Oak Ridge, TN, USA. Available online: http://www.daac.ornl.gov (accessed on 4 February 2003).
- Olson, J.S.; Watts, J.A.; Allison, L.J. Carbon in Live Vegetation of Major World Ecosystem; Oak Ridge National Laboratory: Oak Ridge, TN, USA, 1983; p. 61. [Google Scholar]
- Fang, J.; Guo, Z.; Piao, S.; Chen, A. Terrestrial vegetation carbon sinks in China, 1981–2000. Sci. China Ser. D: Earth Sci 2007, 50, 1341–1350. [Google Scholar]
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R.; Vermote, E.F.; El Saleous, N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int. J. Remote Sens 2005, 26, 4485–4498. [Google Scholar]
- Holben, B.N. Characteristics of maximum-value composite images from temporal AVHRR data. Int. J. Remote Sens 1986, 7, 1417–1434. [Google Scholar]
- Global Modeling and Assimilation Office, File Specification for GEOSDAS Gridded Output Version 5.3; NASA Goddard Space Flight Cent: Greenbelt, MD, USA, 2004.
- Yuan, W.; Liu, S.; Yu, G.; Bonnefond, J.-M.; Chen, J.; Davis, K.; Desai, A.R.; Goldstein, A.H.; Gianelle, D.; Rossi, F.; et al. Global estimates of evapotranspiration and gross primary production based on MODIS and global meteorology data. Remote Sens. Environ 2010, 114, 1416–1431. [Google Scholar]
- Adler, R.F.; Huffman, G.J.; Chang, A.; Ferraro, R.; Xie, P.-P.; Janowiak, J.; Rudolf, B.; Schneider, U.; Curtis, S.; Bolvin, D.; et al. The version-2 global precipitation climatology project (GPCP) monthly precipitation analysis (1979–present). J. Hydrometeorol 2003, 4, 1147–1167. [Google Scholar]
- Rienecker, M.M.; Suarez, M.J.; Gelaro, R.; Todling, R.; Bacmeister, J.; Liu, E.; Bosilovich, M.G.; Schubert, S.D.; Takacs, L.; Kim, G.-K.; et al. MERRA: NASA’s modern-era retrospective analysis for research and applications. J. Clim 2011, 24, 3624–3648. [Google Scholar]
- Zhang, K.; Kimball, J.S.; Mu, Q.; Jones, L.A.; Goetz, S.J.; Running, S.W. Satellite based analysis of northern ET trends and associated changes in the regional water balance from 1983 to 2005. J. Hydrol 2009, 379, 92–110. [Google Scholar]
- Fensholt, R.; Rasmussen, K. Analysis of trends in the Sahelian “rain-use efficiency” using GIMMS NDVI, RFE and GPCP rainfall data. Remote Sens. Environ 2011, 115, 438–451. [Google Scholar]
- Chatterjee, S.; Hadi, A.S. Influential observations, high leverage points, and outliers in linear regression. Stat. Sci 1986, 1, 379–393. [Google Scholar]
- Toms, J.D.; Lesperance, M.L. Piecewise regression: A tool for identifying ecological thresholds. Ecology 2003, 84, 2034–2041. [Google Scholar]
- Parton, W.J.; Scurlock, J.M.O.; Ojima, D.S.; Gilmanov, T.G.; Scholes, R.J.; Schimel, D.S.; Kirchner, T.; Menaut, J.-C.; Seastedt, T.; Garcia Moya, E.; et al. Observations and modeling of biomass and soil organic matter dynamics for the grassland biome worldwide. Glob. Biogeochem. Cycle 1993, 7, 785–809. [Google Scholar]
- Kawamura, K.; Akiyama, T.; Yokota, H.-O.; Tsutsumi, M.; Yasuda, T.; Watanabe, O.; Wang, S. Quantifying grazing intensities using geographic information systems and satellite remote sensing in the Xilingol steppe region, Inner Mongolia, China. Agric. Ecosyst. Environ. 2005, 107, 83–93. [Google Scholar]
- Blanco, L.J.; Aguilera, M.O.; Paruelo, J.M.; Biurrun, F.N. Grazing effect on NDVI across an aridity gradient in Argentina. J. Arid Environ 2008, 72, 764–776. [Google Scholar]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar]
- Bradford, J.B.; Lauenroth, W.K.; Burke, I.C.; Paruelo, J.M. The influence of climate, soils, weather, and land use on primary production and biomass seasonality in the US Great Plains. Ecosystems 2006, 9, 934–950. [Google Scholar]
- Anyamba, A.; Tucker, C.J. Analysis of Sahelian vegetation dynamics using NOAA-AVHRR NDVI data from 1981–2003. J. Arid Environ 2005, 63, 596–614. [Google Scholar]
- John, R.; Chen, J.; Ou-Yang, Z.-T.; Xiao, J.; Becker, R.; Samanta, A.; Ganguly, S.; Yuan, W.; Batkhishig, O. Vegetation response to extreme climate events on the Mongolian Plateau from 2000 to 2010. Environ. Res. Lett 2013. [Google Scholar] [CrossRef]
- Mathews, K.H., Jr.; Vandeveer, M. Beef Production, Markets, and Trade in Argentina and Uruguay: An Overview; United States Department of Agriculture: Washington, DC, USA, 2007. [Google Scholar]
- Qi, J.; Chen, J.; Wan, S.; Ai, L. Understanding the coupled natural and human systems in dryland East Asia. Environ. Res. Lett 2012. [Google Scholar] [CrossRef]
- Tucker, C.J.; Nicholson, S.E. Variations in the size of the Sahara Desert from 1980 to 1997. Ambio 1999, 28, 587–591. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Overall Analysis (1982–2006) | Turning Point (year) | First Time Period | Second Time Period | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Area (104 km2) | Agblive (g·C·m−2) | Total C (Tg·C) | Trend (Tg·C·yr−1) | Rt | Rp | Trend (Tg·C·yr−1) | Rt | Rp | Trend (Tg·C·yr−1) | Rt | Rp | ||
| Global | 1473.76 | 71.4 | 1051.71 | 2.43 ** | 0.52 ** | 0.54 ** | 1994 | 7.71 ** | 0.42 | 0.52 | −2.17 | 0.13 | 0.39 |
| EA | 701.32 | 62.6 | 438.75 | 0.65 | 0.13 | 0.54 * | 1994 | 5.03 ** | 0.23 | 0.68 * | −1.97 * | −0.33 | 0.42 |
| AF | 273.88 | 68.5 | 187.49 | 1.21 ** | 0.3 | 0.86 * | 1994 | 2.0 * | −0.15 | 0.89 ** | 0.15 | −0.27 | 0.72 * |
| NA | 276.72 | 75.1 | 207.72 | 0.33 * | 0.25 | 0.42 * | 1993 | 0.87 | 0.03 | 0.61 * | −0.42 | −0.29 | 0.51 |
| SA | 125.2 | 113.4 | 141.97 | −0.04 | −0.19 | 0.35 | 1997 | 0.2 | −0.13 | 0.06 | −0.93 ** | 0.003 | 0.6 |
| AZ | 96.64 | 78.4 | 75.79 | 0.29 | −0.78 ** | 0.82 * | - | - | - | - | - | - | - |
| Kazakhstan | 203.48 | 55.9 | 113.8 | −0.13 | −0.24 | 0.47 * | 1993 | 2.14 ** | −0.14 | 0.66 * | −1.57 ** | −0.52 | 0.41 |
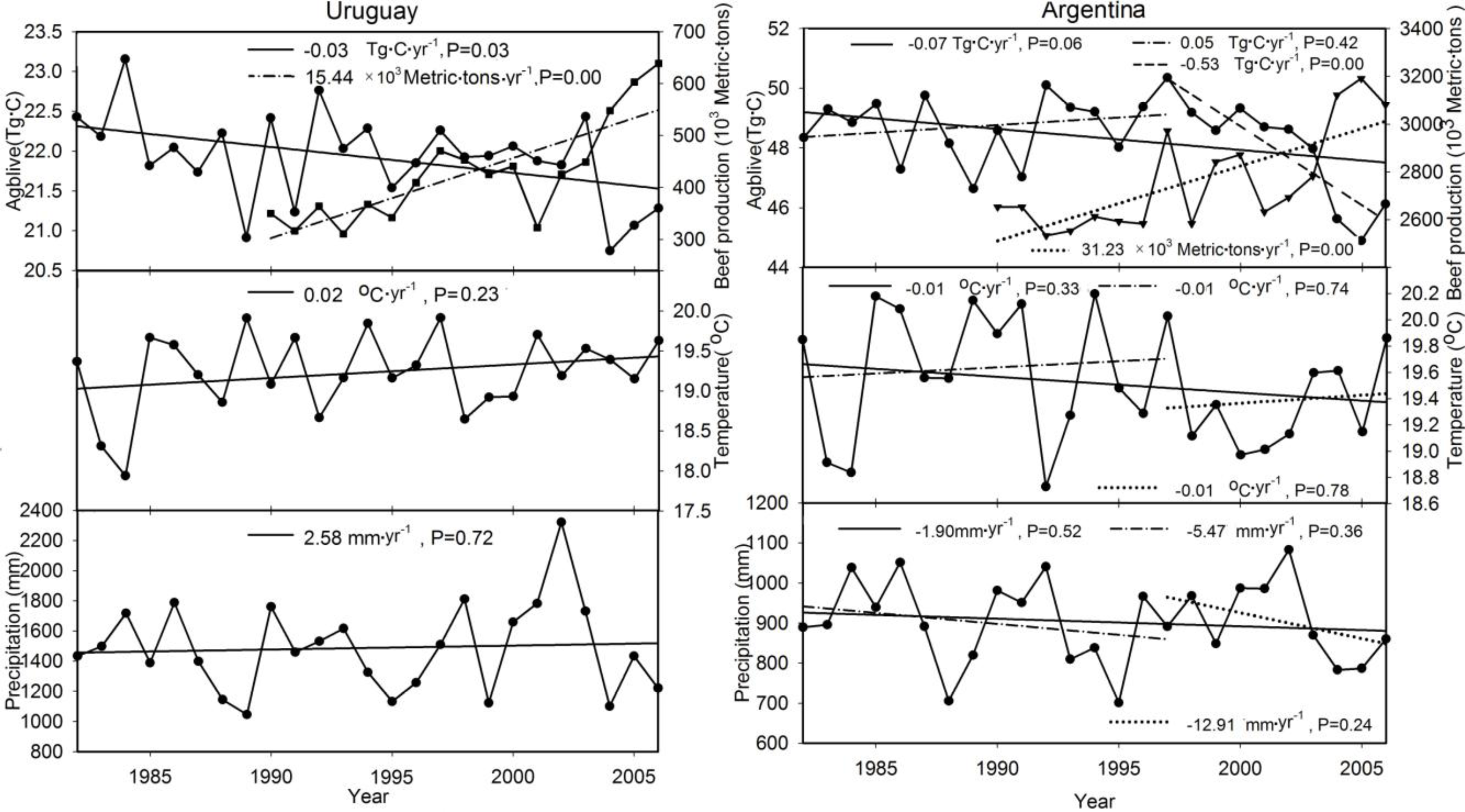
| Uruguay | 15 | 145.8 | 21.9 | −0.03 * | −0.48 * | 0.42 * | - | - | - | - | - | - | - |
| Argentina | 40.1 | 120.3 | 48.4 | −0.07 | −0.24 | 0.38 | 1997 | 0.05 | −0.42 | 0.16 | −0.53 ** | −0.08 | 0.66 * |
| Region | Overall Analysis (1982–2006) | First Time Period | Second Time Period | |||||
|---|---|---|---|---|---|---|---|---|
| Pr (mm) | Ta (°C) | TPr (mm·yr−1) | TTa (°C·yr−1) | TPr (mm·yr−1) | TTa (°C·yr−1) | TPr (mm·yr−1) | TTa (°C·yr−1) | |
| Global | 558.8 | 12.4 | 0.81 | 0.04 ** | 1.05 | 0.04 | 0.24 | 0.05 ** |
| EA | 453.7 | 5.3 | 0.07 | 0.05 ** | 3.50 | 0.02 | −0.19 | 0.06 * |
| AF | 465.5 | 26.5 | 3.37 ** | 0.04 ** | 5.15 | 0.04 | 0.01 | 0.05 * |
| NA | 522.7 | 9.3 | −1.62 | 0.06 ** | 0.02 | 0.07 | −5.59 | 0.09 ** |
| SA | 1294.4 | 18.3 | −0.81 | 0.00 | −7.48 | 0.02 | −14.16 | 0.01 |
| AZ | 736.7 | 25.3 | 8.00 | 0.00 | - | - | - | - |
| Precipitation Trend | Temperature Trend | Biomass C Trend | Regions |
|---|---|---|---|
| decrease | increase | Decrease | Southern Oklahoma, Central Texas, USA |
| increase | decrease | None | Northern Territory of Australia |
| increase | none | Increase | Southern Africa and Western Australia and Queensland of Australia |
| none | increase | Decrease | Central and Eastern Kazakhstan |
| increase | increase | Increase | Sahel region of Africa |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xia, J.; Liu, S.; Liang, S.; Chen, Y.; Xu, W.; Yuan, W. Spatio-Temporal Patterns and Climate Variables Controlling of Biomass Carbon Stock of Global Grassland Ecosystems from 1982 to 2006. Remote Sens. 2014, 6, 1783-1802. https://doi.org/10.3390/rs6031783
Xia J, Liu S, Liang S, Chen Y, Xu W, Yuan W. Spatio-Temporal Patterns and Climate Variables Controlling of Biomass Carbon Stock of Global Grassland Ecosystems from 1982 to 2006. Remote Sensing. 2014; 6(3):1783-1802. https://doi.org/10.3390/rs6031783
Chicago/Turabian StyleXia, Jiangzhou, Shuguang Liu, Shunlin Liang, Yang Chen, Wenfang Xu, and Wenping Yuan. 2014. "Spatio-Temporal Patterns and Climate Variables Controlling of Biomass Carbon Stock of Global Grassland Ecosystems from 1982 to 2006" Remote Sensing 6, no. 3: 1783-1802. https://doi.org/10.3390/rs6031783
APA StyleXia, J., Liu, S., Liang, S., Chen, Y., Xu, W., & Yuan, W. (2014). Spatio-Temporal Patterns and Climate Variables Controlling of Biomass Carbon Stock of Global Grassland Ecosystems from 1982 to 2006. Remote Sensing, 6(3), 1783-1802. https://doi.org/10.3390/rs6031783