
Monitoring Natural Ecosystem and Ecological Gradients: Perspectives with EnMAP

 , and
, and
Abstract
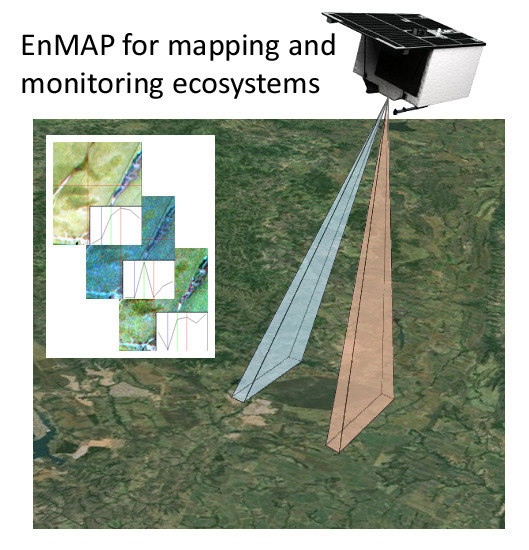
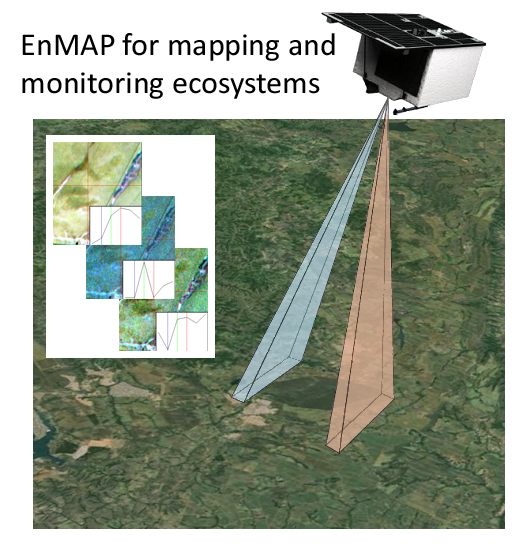
:
1. Introduction

2. Monitoring Ecological Gradients with Spaceborne IS Data
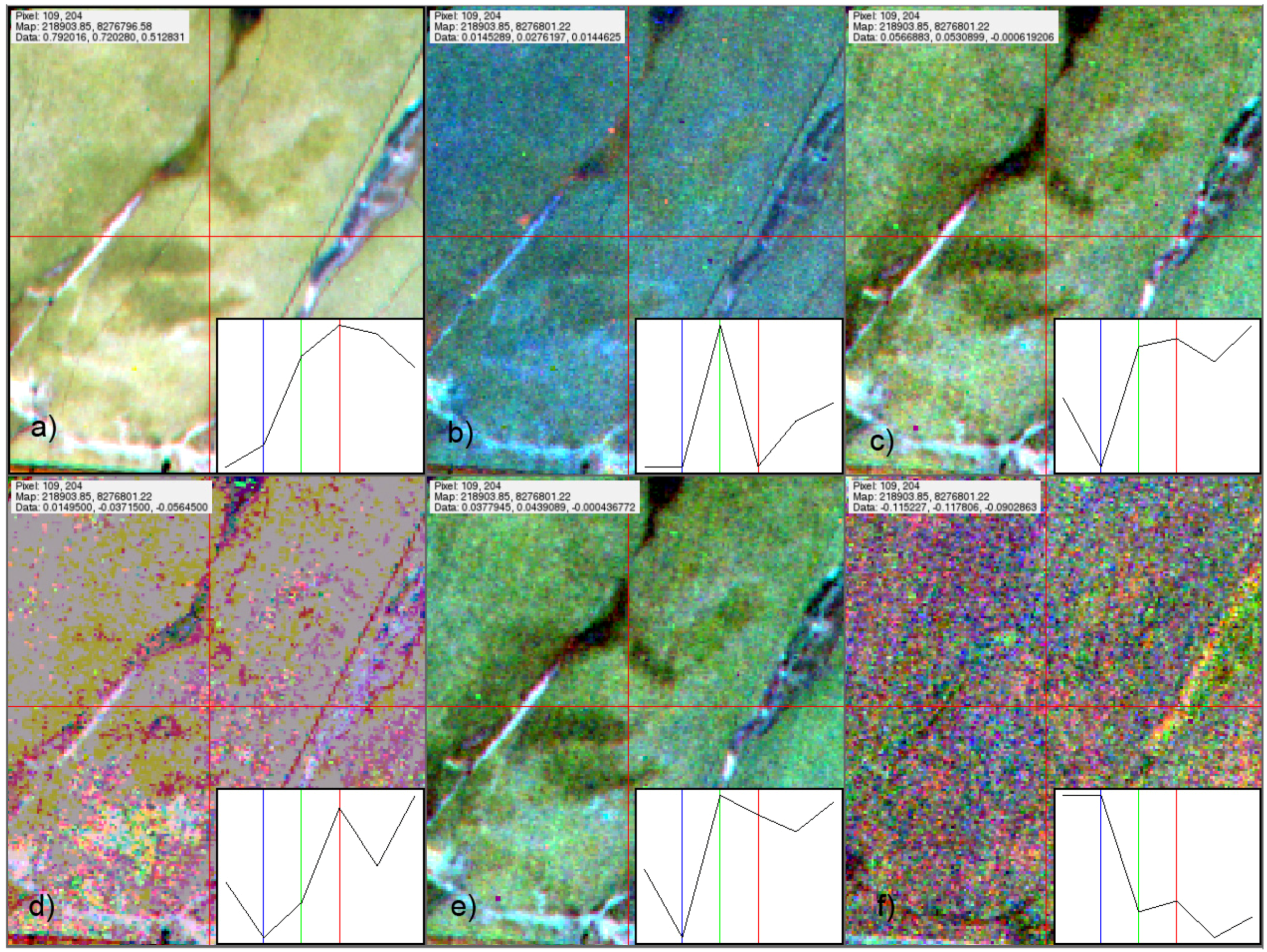
2.1. Common Methodological Approach

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Usage | Spectral Bands (nm) | Reference |
|---|---|---|---|
| Normalized Difference Vegetation Index (NDVI) | Structure, vigor | 670, 800 | [60] |
| Modified Chlorophyll Absorption in Reflectance Index (MCARI) | Chlorophyll | 550, 670, 700 | [65] |
| Leaf Water Vegetation Index (LWVI2) | Leaf water | 1094, 1205 | [66] |
| Cellulose Absorption Index (CAI) | Cellulose | 2000, 2100, 2200 | [67] |
| Normalized Difference Lignin Index (NDLI) | Lignin | 1680, 1754 | [61] |
| Normalized Difference Nitrogen Index (NDNI) | Nitrogen | 1510, 1680 | [61] |
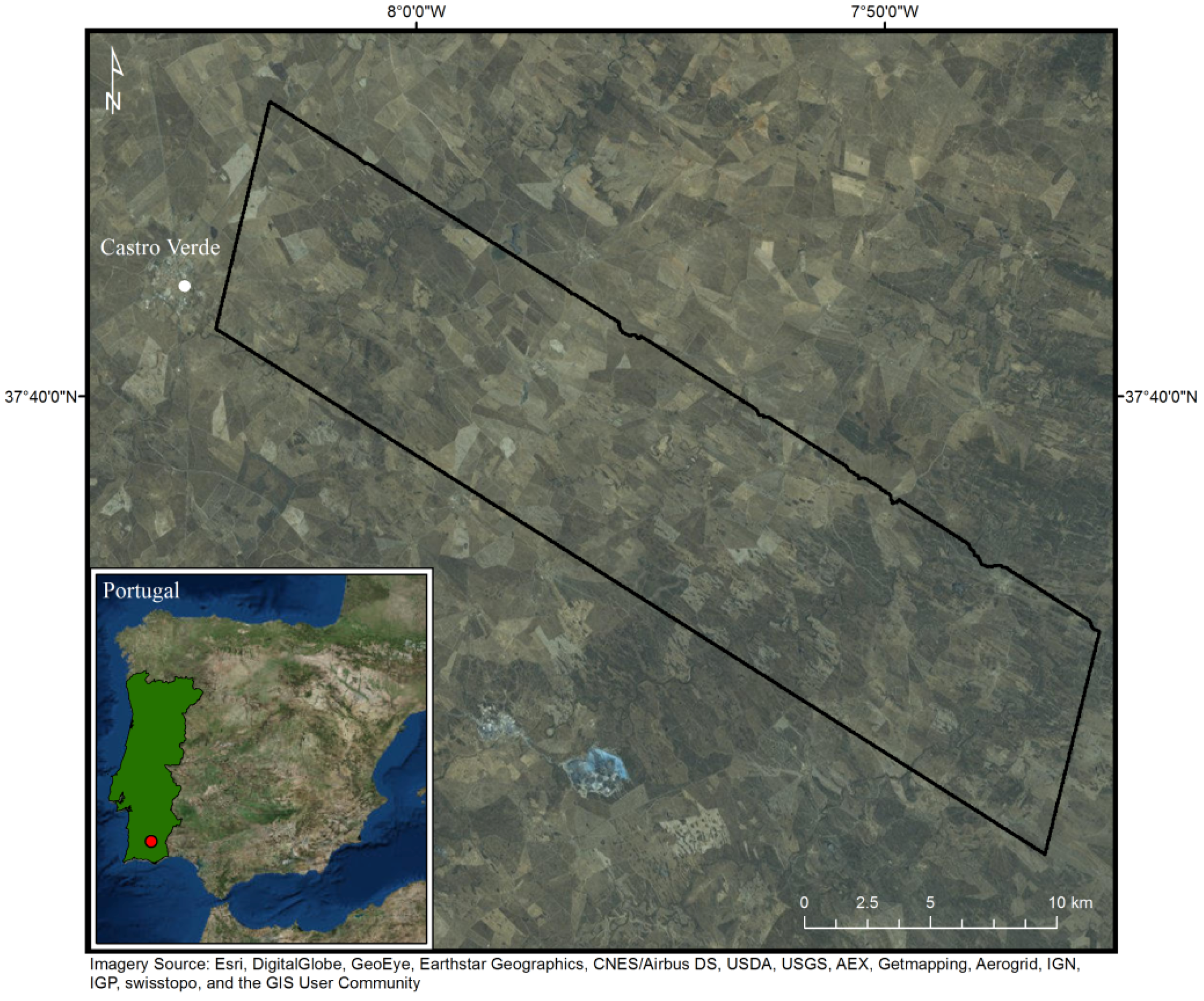
2.2. Gradual Ecosystem Transitions: Shrub Encroachment in Southern Portugal

| April | August | Time-Stack | |
|---|---|---|---|
| NDVI | 23% | 61% | 46% (August) |
| MCARI | 24% | • | 13% (April) |
| LWVI2 | • | 24% | 19% (August) |
| CAI | 19% | • | • |
| NDLI | 34% | • | 18% (April) |
| NDNI | • | • | • |
| r2 | 0.159 | 0.331 | 0.446 |

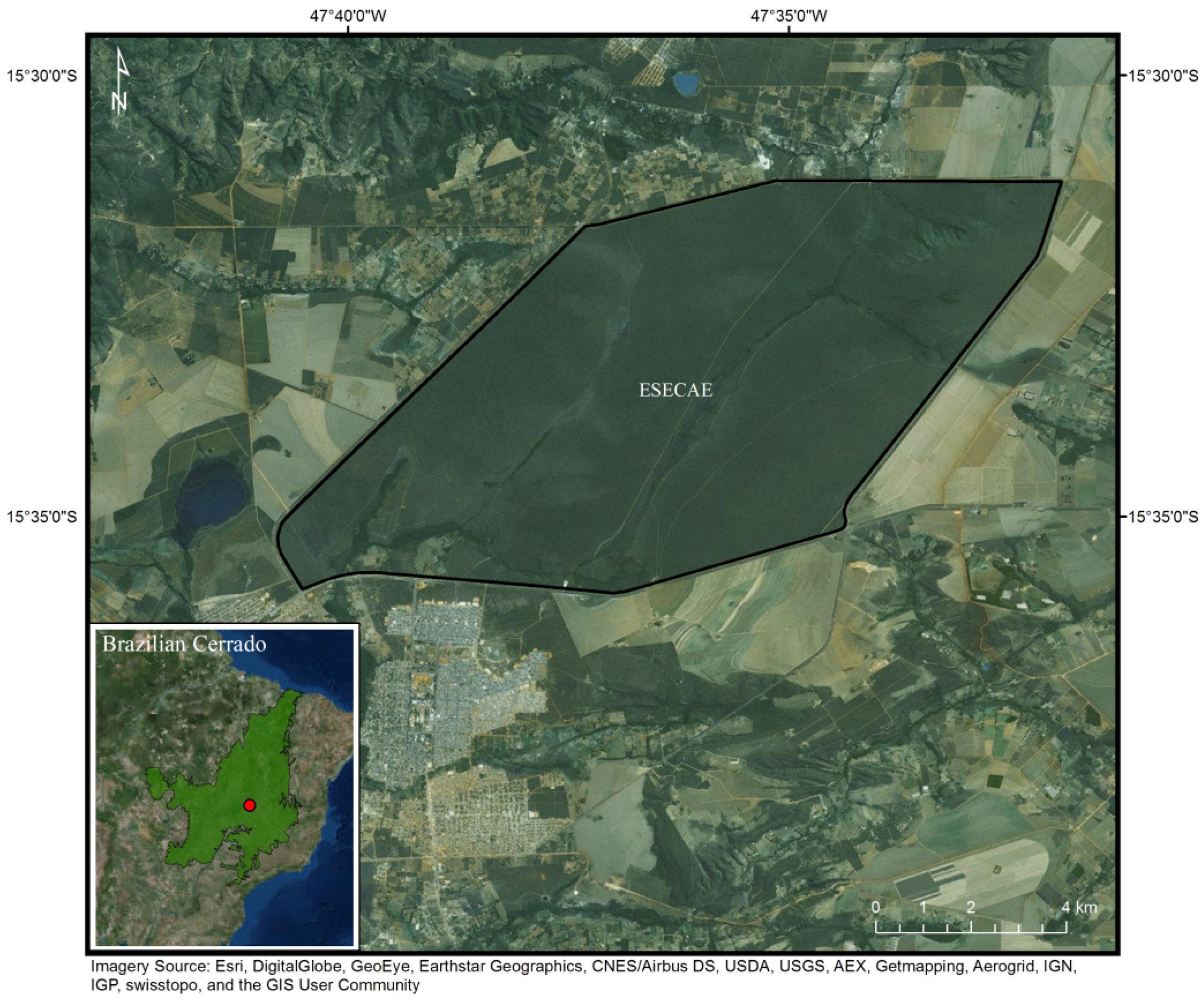
2.3. Brazilian Cerrado

| Date | View Angle (°) | Season |
|---|---|---|
| 2006-08-29 | 6.25 | Dry season |
| 2006-09-13 | 13.12 | End of dry season |
| 2006-11-19 | −5.77 | Beginning of wet season |
| 2007-02-10 | 17.51 | Wet season |
| 2007-03-02 | 2.20 | Wet season |
| 2007-05-17 | 6.70 | End of wet season |


| September | November | February | March | May | Time-Stack | |
|---|---|---|---|---|---|---|
| NDVI | 96% | 33% | 57% | 70% | 77% | 85% (September) |
| MCARI | • | • | • | • | • | • |
| LWVI2 | • | 28% | 34% | • | 19% | • |
| CAI | • | • | • | 20% | • | • |
| NDLI | • | • | • | • | • | • |
| NDNI | • | • | • | • | • | • |
| r2 | 0.688 | 0.608 | 0.392 | 0.492 | 0.590 | 0.681 |
| NDVI | MCARI | LWVI2 | CAI | NDLI | NDNI | |
|---|---|---|---|---|---|---|
| September | 94% | 81% | 18% | 25% | 16% | 60% |
| November | • | 19% | 77% | • | 79% | • |
| February | • | • | • | • | • | 39% |
| March | • | • | • | 50% | • | • |
| May | • | • | • | 22% | • | • |
| r2 | 0.680 | 0.539 | 0.581 | 0.477 | 0.572 | 0.097 |
3. Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Kareiva, P.; Watts, S.; McDonald, R.; Boucher, T. Domesticated nature: Shaping landscapes and ecosystems for human welfare. Science 2007, 316, 1866–1869. [Google Scholar] [CrossRef]
- Sitch, S.; Smith, B.; Prentice, I.C.; Arneth, A.; Bondeau, A.; Cramer, W.; Kaplan, J.O.; Levis, S.; Lucht, W.; Sykes, M.T.; et al. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model. Glob. Change Biol. 2003, 9, 161–185. [Google Scholar] [CrossRef]
- McIntosh, R.P. Continuum concept of vegetation. Bot. Rev. 1967, 33, 130–187. [Google Scholar] [CrossRef]
- Muller, F. Gradients in ecological systems. Ecol. Model. 1998, 108, 3–21. [Google Scholar] [CrossRef]
- Gosz, J.R. Gradient analysis of ecological change in time and space: Implications for forest management. Ecol. Appl. 1992, 2, 248–261. [Google Scholar] [CrossRef]
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Schimel, D.S. Terrestrial ecosystems and the carbon-cycle. Glob. Change Biol. 1995, 1, 77–91. [Google Scholar] [CrossRef]
- Ardö, J.; Mölder, M.; El-Tahir, B.A.; Abdalla, H.; Elkhidir, M. Seasonal variation of carbon fluxes in a sparse savanna in semi arid Sudan. Carbon Balance Manag. 2008, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Serreze, M.C.; Walsh, J.E.; Chapin, F.S.; Osterkamp, T.; Dyurgerov, M.; Romanovsky, V.; Oechel, W.C.; Morison, J.; Zhang, T.; Barry, R.G. Observational evidence of recent change in the northern high-latitude environment. Clim. Change 2000, 46, 159–207. [Google Scholar] [CrossRef]
- McDonnell, M.J.; Pickett, S.T.A. Ecosystem structure and function along urban rural gradients: An unexploited opportunity for ecology. Ecology 1990, 71, 1232–1237. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Bowker, M.A.; Maestre, F.T.; Roger, E.; Reynolds, J.F.; Whitford, W.G. Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecol. Lett. 2011, 14, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Attiwill, P.M. The disturbance of forest ecosystems—The ecological basis for conservative management. For. Ecol. Manag. 1994, 63, 247–300. [Google Scholar] [CrossRef]
- Adler, P.B.; Raff, D.A.; Lauenroth, W.K. The effect of grazing on the spatial heterogeneity of vegetation. Oecologia 2001, 128, 465–479. [Google Scholar]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Estel, S.; Kuemmerle, T.; Alcántara, C.; Levers, C.; Prishchepov, A.; Hostert, P. Mapping farmland abandonment and recultivation across Europe using MODIS NDVI time series. Remote Sens. Environ. 2015, 163, 312–325. [Google Scholar] [CrossRef]
- Müller, H.; Leitão, P.J.; Hostert, P. Vegetation dynamics, carbon stocks and turnover rates in the Amazon—Upscaling local processes with remote sensing time series. In 43rd Annual Meeting of the Ecological Society of Germany, Austria and Switzerland, Potsdam, Germany, 9–13 September 2013.
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef] [Green Version]
- Lavorel, S.; Grigulis, K.; Lamarque, P.; Colace, M.-P.; Garden, D.; Girel, J.; Pellet, G.; Douzet, R. Using plant functional traits to understand the landscape distribution of multiple ecosystem services. J. Ecol. 2011, 99, 135–147. [Google Scholar] [CrossRef]
- Sala, O.E.; Maestre, F.T. Grass-woodland transitions: determinants and consequences for ecosystem functioning and provisioning of services. J. Ecol. 2014, 102, 1357–1362. [Google Scholar] [CrossRef]
- Field, C.B.; Randerson, J.T.; Malmstrom, C.M. Global net primary production: Combining ecology and remote-sensing. Remote Sens. Environ. 1995, 51, 74–88. [Google Scholar] [CrossRef]
- De Fries, R.; Pagiola, S.; Adamowicz, W.L.; Akçakaya, H.R.; Arcenas, A.; Babu, S.; Balk, D.; Confalonieri, U.; Cramer, W.; Falconí, F.; et al. Analytical approaches for assessing ecosystem condition and human well-being. In Ecosystems and Human Well-Being: Current State and Trends; Hassan, R., Scholes, R., Ash, N., Eds.; Island Press: Washington, DC, USA, 2005; Chapter 2; pp. 37–71. [Google Scholar]
- Hoare, D.; Frost, P. Phenological description of natural vegetation in southern Africa using remotely-sensed vegetation data. Appl. Veg. Sci. 2004, 7, 19–28. [Google Scholar] [CrossRef]
- Fisher, J.I.; Mustard, J.F.; Vadeboncoeur, M.A. Green leaf phenology at Landsat resolution: Scaling from the field to the satellite. Remote Sens. Environ. 2006, 100, 265–279. [Google Scholar] [CrossRef]
- Kennedy, R.E.; Andrefouet, S.; Cohen, W.B.; Gomez, C.; Griffiths, P.; Hais, M.; Healey, S.P.; Helmer, E.H.; Hostert, P.; Lyons, M.B.; et al. Bringing an ecological view of change to landsat-based remote sensing. Front. Ecol. Environ. 2014, 12, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Vogelmann, J.E.; Xian, G.; Homer, C.; Tolk, B. Monitoring gradual ecosystem change using Landsat time series analyses: Case studies in selected forest and rangeland ecosystems. Remote Sens. Environ. 2012, 122, 92–105. [Google Scholar] [CrossRef]
- Homer, C.G.; Meyer, D.K.; Aldridge, C.L.; Schell, S.J. Detecting annual and seasonal changes in a sagebrush ecosystem with remote sensing-derived continuous fields. J. Appl. Remote Sens. 2013, 7 (1), 073508. [Google Scholar] [CrossRef]
- Okujeni, A.; van der Linden, S.; Hostert, P. Extending the vegetation-impervious-soil model using simulated EnMAP data and machine learning. Remote Sens. Environ. 2015, 158, 69–80. [Google Scholar] [CrossRef]
- Shoshany, M. Satellite remote sensing of natural Mediterranean vegetation: A review within an ecological context. Progr. Phys. Geogr. 2000, 24, 153–178. [Google Scholar] [CrossRef]
- Ustin, S.L.; Gamon, J.A. Remote sensing of plant functional types. New Phytol. 2010, 186, 795–816. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Cohen, W.B.; Kennedy, R.E.; Maiersperger, T.K.; Gower, S.T. Hyperspectral versus multispectral data for estimating leaf area index in four different biomes. Remote Sens. Environ. 2004, 91, 508–520. [Google Scholar] [CrossRef]
- Smith, M.L.; Ollinger, S.V.; Martin, M.E.; Aber, J.D.; Hallett, R.A.; Goodale, C.L. Direct estimation of aboveground forest productivity through hyperspectral remote sensing of canopy nitrogen. Ecol. Appl. 2002, 12, 1286–1302. [Google Scholar] [CrossRef]
- Mutanga, O.; Skidmore, A.K. Narrow band vegetation indices overcome the saturation problem in biomass estimation. Int. J. Remote Sens. 2004, 25, 3999–4014. [Google Scholar] [CrossRef]
- Fuentes, D.A.; Gamon, J.A.; Cheng, Y.; Claudio, H.C.; Qiu, H.-L.; Mao, Z.; Sims, D.A.; Rahman, A.F.; Oechel, W.; Luo, H. Mapping carbon and water vapor fluxes in a chaparral ecosystem using vegetation indices derived from AVIRIS. Remote Sens. Environ. 2006, 103, 312–323. [Google Scholar] [CrossRef]
- Asner, G.P.; Elmore, A.J.; Hughes, R.F.; Warner, A.S.; Vitousek, P.M. Ecosystem structure along bioclimatic gradients in Hawaii from imaging spectroscopy. Remote Sens. Environ. 2005, 96, 497–508. [Google Scholar] [CrossRef]
- Schmidtlein, S.; Zimmermann, P.; Schüpferling, R.; Weiß, C. Mapping the floristic continuum: Ordination space position estimated from imaging spectroscopy. J. Veg. Sci. 2007, 18, 131–140. [Google Scholar] [CrossRef]
- Leitão, P.J.; Schwieder, M.; Suess, S.; Catry, I.; Milton, E.J.; Moreira, F.; Osborne, P.E.; Pinto, M.J.; van der Linden, S.; Hostert, P. Mapping beta diversity from space: Sparse generalised dissimilarity modelling (SGDM) for analysing high-dimensional data. Methods Ecol. Evol. 2015. [Google Scholar] [CrossRef]
- Oldeland, J.; Dorigo, W.; Wesuls, D.; Jürgens, N. Mapping bush encroaching species by seasonal differences in hyperspectral imagery. Remote Sens. 2010, 2, 1416–1438. [Google Scholar] [CrossRef]
- Harris, A.T.; Asner, G.P. Grazing gradient detection with airborne imaging spectroscopy on a semi-arid rangeland. J. Arid Environ. 2003, 55, 391–404. [Google Scholar] [CrossRef]
- Underwood, E.; Ustin, S.; DiPietro, D. Mapping nonnative plants using hyperspectral imagery. Remote Sens. Environ. 2003, 86, 150–161. [Google Scholar] [CrossRef]
- Guanter, L.; Kaufmann, H.; Segl, K.; Chabrillat, S.; Förster, S.; Rogass, C.; Kuester, T.; Hollstein, A.; Rossner, G.; Chlebek, C.; et al. The EnMAP spaceborne imaging spectroscopy mission for Earth Observation. Remote Sens. 2015, 7, 8830–8857. [Google Scholar] [CrossRef]
- Segl, K.; Guanter, L.; Rogass, C.; Kuester, T.; Roessner, S.; Kaufmann, H.; Sang, B.; Mogulsky, V.; Hofer, S. EeteS—The EnMAP End-to-End Simulation Tool. IEEE J. Sel. Topics Appl. Earth Obs. Remote Sens. 2012, 5, 522–530. [Google Scholar] [CrossRef]
- Schwieder, M.; Leitão, P.J.; Suess, S.; Senf, C.; Hostert, P. Estimating fractional shrub cover using simulated EnMAP data: A comparison of three machine learning regression techniques. Remote Sens. 2014, 6, 3427–3445. [Google Scholar] [CrossRef]
- Ustin, S.L.; Roberts, D.A.; Gamon, J.A.; Asner, G.P.; Green, R.O. Using imaging spectroscopy to study ecosystem processes and properties. BioScience 2004, 54, 523–534. [Google Scholar] [CrossRef]
- Asner, G.P.; Nepstad, D.; Cardinot, G.; Ray, D. Drought stress and carbon uptake in an Amazon forest measured with spaceborne imaging spectroscopy. Proc. Natl. Acad. Sci. USA 2004, 101, 6039–6044. [Google Scholar] [CrossRef] [PubMed]
- Stagakis, S.; Markos, N.; Sykioti, O.; Kyparissis, A. Monitoring canopy biophysical and biochemical parameters in ecosystem scale using satellite hyperspectral imagery: An application on a Phlomis fruticosa Mediterranean ecosystem using multiangular CHRIS/PROBA observations. Remote Sens. Environ. 2010, 114, 977–994. [Google Scholar] [CrossRef]
- Torres-Madronero, M.C.; Velez-Reyes, M.; Van Bloem, S.J.; Chinea, J.D. Multi-temporal unmixing analysis of Hyperion images over the Guanica Dry Forest. IEEE J. Sel. Topics Appl. Earth Obs. Remote Sens. 2012, 6, 329–338. [Google Scholar]
- Campbell, P.K.E.; Middleton, E.M.; Thome, K.J.; Kokaly, R.F.; Huemmrich, K.F.; Lagomasino, D.; Novick, K.A.; Brunsell, N.A. EO-1 Hyperion reflectance time series at calibration and validation sites: Stability and sensitivity to seasonal dynamics. IEEE J. Sel. Topics Appl. Earth Obs. Remote Sens. 2013, 6, 276–290. [Google Scholar] [CrossRef]
- Somers, B.; Asner, G.P. Hyperspectral time series analysis of native and invasive species in Hawaiian rainforests. Remote Sens. 2012, 4, 2510–2529. [Google Scholar] [CrossRef]
- Somers, B.; Asner, G.P. Invasive species mapping in Hawaiian rainforests using multi-temporal Hyperion spaceborne imaging spectroscopy. IEEE J. Sel. Topics Appl. Earth Obs. Remote Sens. 2013, 6, 351–359. [Google Scholar] [CrossRef]
- Numata, I.; Cochrane, M.A.; Souza, C.M., Jr.; Sales, M.H. Carbon emissions from deforestation and forest fragmentation in the Brazilian Amazon. Environ. Res. Lett. 2011, 6, 044003. [Google Scholar] [CrossRef]
- Kaufmann, H.; Förster, S.; Wulf, H.; Segl, K.; Guanter, L.; Bochow, M.; Heiden, U.; Müller, A.; Heldens, W.; Schneiderhan, T.; et al. Science Plan of the Environmental Mapping and Analysis Program (EnMAP); Deutsches Geo Forschungs Zentrum GFZ: Potsdam, Gemany, 2012. [Google Scholar]
- Suess, S.; van der Linden, S.; Okujeni, A.; Schwieder, M.; Leitão, P.J.; Hostert, P. Using class-probabilities to map gradual transitions in shrub vegetation maps from simulated EnMAP data. Remote Sens. 2015, 7, 10668–10688. [Google Scholar] [CrossRef]
- Vapnik, V.N. An overview of statistical learning theory. IEEE Trans. Neural Netw. 1999, 10, 988–999. [Google Scholar] [CrossRef] [PubMed]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Verrelst, J.; Munoz, J.; Alonso, L.; Delegido, J.; Rivera, J.P.; Camps-Valls, G.; Moreno, J. Machine learning regression algorithms for biophysical parameter retrieval: Opportunities for Sentinel-2 and -3. Remote Sens. Environ. 2012, 118, 127–139. [Google Scholar] [CrossRef]
- Pal, M.; Foody, G.M. Feature selection for classification of hyperspectral data by SVM. IEEE Trans. Geosci. Remote Sens. 2010, 48, 2297–2307. [Google Scholar] [CrossRef]
- Held, M.; Rabe, A.; Senf, C.; van der Linden, S.; Hostert, P. Analyzing hyperspectral and hypertemporal data by decoupling feature redundancy and feature relevance. IEEE Geosci. Remote Sens. Lett. 2015, 12, 983–987. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Pattey, E.; Zarco-Tejada, P.J.; Strachan, I.B. Hyperspectral vegetation indices and novel algorithms for predicting green LAI of crop canopies: Modeling and validation in the context of precision agriculture. Remote Sens. Environ. 2004, 90, 337–352. [Google Scholar] [CrossRef]
- Serrano, L.; Peñuelas, J.; Ustin, S.L. Remote sensing of nitrogen and lignin in Mediterranean vegetation from AVIRIS data: Decomposing biochemical from structural signals. Remote Sens. Environ. 2002, 81, 355–364. [Google Scholar] [CrossRef]
- Ferwerda, J.G.; Skidmore, A.K.; Mutanga, O. Nitrogen detection with hyperspectral normalized ratio indices across multiple plant species. Int. J. Remote Sens. 2005, 26, 4083–4095. [Google Scholar] [CrossRef]
- Austin, M.P. Spatial prediction of species distribution: An interface between ecological theory and statistical modelling. Ecol. Model. 2002, 157, 101–118. [Google Scholar] [CrossRef]
- van der Linden, S.; Rabe, A.; Held, M.; Jakimow, B.; Leitão, P.J.; Okujeni, A.; Suess, S.; Hostert, P. The EnMAP-Box—A toolbox and application programming interface for EnMAP data processing. Remote Sens. 2015, 7, 11249–11266. [Google Scholar] [CrossRef]
- Daughtry, C.S.T.; Walthall, C.L.; Kim, M.S.; de Colstoun, E.B.; McMurtrey, J.E. Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance. Remote Sens. Environ. 2000, 74, 229–239. [Google Scholar] [CrossRef]
- Galvão, L.S.; Formaggio, A.R.; Tisot, D.A. Discrimination of sugarcane varieties in Southeastern Brazil with EO-1 Hyperion data. Remote Sens. Environ. 2005, 94, 523–534. [Google Scholar] [CrossRef]
- Nagler, P.L.; Inoue, Y.; Glenn, E.P.; Russ, A.L.; Daughtry, C.S.T. Cellulose absorption index (CAI) to quantify mixed soil-plant litter scenes. Remote Sens. Environ. 2003, 87, 310–325. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Shirley, S.M.; Yang, Z.; Hutchinson, R.A.; Alexander, J.D.; McGarigal, K.; Betts, M.G. Species distribution modelling for the people: Unclassified landsat TM imagery predicts bird occurrence at fine resolutions. Divers. Distrib. 2013, 19, 855–866. [Google Scholar] [CrossRef]
- Cheong, Y.L.; Leitão, P.J.; Lakes, T. Assessment of land use factors associated with dengue cases in Malaysia using Boosted Regression Trees. Spat. Spatio-Temporal Epidemiol. 2014, 10, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Müller, D.; Leitão, P.J.; Sikor, T. Comparing the determinants of cropland abandonment in Albania and Romania using Boosted Regression Trees. Agric. Syst. 2013, 117, 66–77. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; Version 2.6.0; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Ridgeway, G. Generalized Boosted Models: A Guide to the GBM Package, Version 2.1.1. 2007.
- Leitão, P.J.; Moreira, F.; Osborne, P.E. Breeding habitat selection by steppe birds in Castro Verde: A remote sensing and advanced statistics approach. Ardeola 2010, 57, 93–116. [Google Scholar]
- Moreira, F.; Leitão, P.J.; Morgado, R.; Alcazar, R.; Cardoso, A.; Carrapato, C.; Delgado, A.; Geraldes, P.; Gordinho, L.; Henriques, I.; et al. Spatial distribution patterns, habitat correlates and population estimates of steppe birds in Castro Verde. Airo 2007, 17, 5–30. [Google Scholar]
- Kuemmerle, T.; Hostert, P.; Radeloff, V.C.; Perzanowski, K.; Kruhlov, I. Post-socialist farmland abandonment in the Carpathians. Ecosystems 2008, 11, 614–628. [Google Scholar] [CrossRef]
- Marta-Pedroso, C.; Domingos, T.; Freitas, H.; de Groot, R.S. Cost-benefit analysis of the Zonal Program of Castro Verde (Portugal): Highlighting the trade-off between biodiversity and soil conservation. Soil Tillage Res. 2007, 97, 79–90. [Google Scholar] [CrossRef]
- Calvão, T.; Palmeirim, J.M. Mapping Mediterranean scrub with satellite imagery: Biomass estimation and spectral behaviour. Int. J. Remote Sens. 2004, 25, 3113–3126. [Google Scholar] [CrossRef]
- Klink, C.A.; Machado, R.B. Conservation of the Brazilian Cerrado. Conserv. Biol. 2005, 19, 707–713. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Ratter, J.A.; Ribeiro, J.F.; Bridgewater, S. The Brazilian cerrado vegetation and threats to its biodiversity. Ann. Bot. 1997, 80, 223–230. [Google Scholar] [CrossRef]
- Ferreira, M.E.; Ferreira, L.G.; Miziara, F.; Soares-Filho, B.S. Modeling landscape dynamics in the central Brazilian savanna biome: future scenarios and perspectives for conservation. J. Land Use Sci. 2012, 8, 403–421. [Google Scholar] [CrossRef]
- Batlle-Bayer, L.; Batjes, N.H.; Bindraban, P.S. Changes in organic carbon stocks upon land use conversion in the Brazilian Cerrado: A review. Agric. Ecosyst. Environ. 2010, 137, 47–58. [Google Scholar] [CrossRef]
- Sano, E.E.; Rosa, R.; Brito, J.L.S.; Ferreira, L.G. Land cover mapping of the tropical savanna region in Brazil. Environ. Monit. Assess. 2010, 166, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.S.; Cox, F.R. Cerrado vegetation in Brazil: An edaphic gradient. Agron. J. 1977, 69, 828–831. [Google Scholar] [CrossRef]
- De Castro, E.A.; Kauffman, J.B. Ecosystem structure in the Brazilian Cerrado: A vegetation gradient of aboveground biomass, root mass and consumption by fire. J. Trop. Ecol. 1998, 14, 263–283. [Google Scholar] [CrossRef]
- Ribeiro, L.F.; Tabarelli, M. A structural gradient in cerrado vegetation of Brazil: Changes in woody plant density, species richness, life history and plant composition. J. Trop. Ecol. 2002, 18, 775–794. [Google Scholar]
- Schwieder, M.; Leitão, P.J.P.; Rabe, A.; Bustamante, M.M.C.; Ferreira, L.G.; Hostert, P. Mapping Cerrado physiognomies using Landsat time series based phenological profiles. In Proceedings of the XVII Simpósio Brasileiro de Sensoriamento Remoto, INPE, João Pessoa, Brazil, 27 April 2015; pp. 3656–3663.
- Rogass, C.; Guanter, L.; Mielke, C.; Scheffler, D.; Boesche, N.K.; Lubitz, C.; Brell, M.; Spengler, D.; Segl, K. An automated processing chain for the retrieval of georeferenced reflectance data from hyperspectral EO-1 Hyperion acquisitions. In Proceedings of the 34th EARSeL Symposium, Warsaw, Poland, 16–20 June 2014. [CrossRef]
- Ferreira, L.G.; Yoshioka, H.; Huete, A.; Sano, E.E. Seasonal landscape and spectral vegetation index dynamics in the Brazilian Cerrado: An analysis within the Large-Scale Biosphere-Atmosphere Experiment in Amazonia (LBA). Remote Sens. Environ. 2003, 87, 534–550. [Google Scholar] [CrossRef]
- Ratana, P.; Huete, A.R.; Ferreira, L. Analysis of cerrado physiognomies and conversion in the MODIS seasonal-temporal domain. Earth Interact. 2005, 9, 1–22. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leitão, P.J.; Schwieder, M.; Suess, S.; Okujeni, A.; Galvão, L.S.; Linden, S.V.d.; Hostert, P. Monitoring Natural Ecosystem and Ecological Gradients: Perspectives with EnMAP. Remote Sens. 2015, 7, 13098-13119. https://doi.org/10.3390/rs71013098
Leitão PJ, Schwieder M, Suess S, Okujeni A, Galvão LS, Linden SVd, Hostert P. Monitoring Natural Ecosystem and Ecological Gradients: Perspectives with EnMAP. Remote Sensing. 2015; 7(10):13098-13119. https://doi.org/10.3390/rs71013098
Chicago/Turabian StyleLeitão, Pedro J., Marcel Schwieder, Stefan Suess, Akpona Okujeni, Lênio Soares Galvão, Sebastian Van der Linden, and Patrick Hostert. 2015. "Monitoring Natural Ecosystem and Ecological Gradients: Perspectives with EnMAP" Remote Sensing 7, no. 10: 13098-13119. https://doi.org/10.3390/rs71013098
APA StyleLeitão, P. J., Schwieder, M., Suess, S., Okujeni, A., Galvão, L. S., Linden, S. V. d., & Hostert, P. (2015). Monitoring Natural Ecosystem and Ecological Gradients: Perspectives with EnMAP. Remote Sensing, 7(10), 13098-13119. https://doi.org/10.3390/rs71013098