
Airborne Lidar for Woodland Habitat Quality Monitoring: Exploring the Significance of Lidar Data Characteristics when Modelling Organism-Habitat Relationships


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Field Site and Bird Breeding Data
2.2. Airborne Lidar Data Acquisition and Pre-Processing

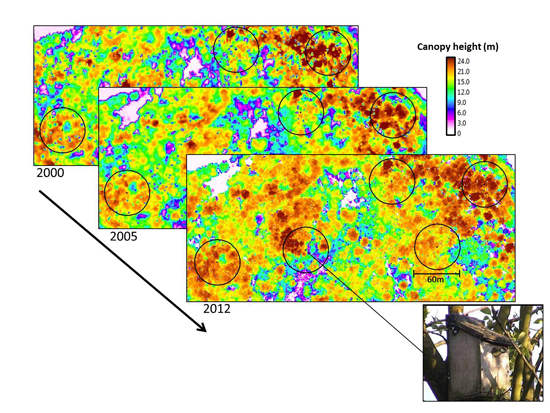{kind=link}
{kind=link}
| Acquisition Parameter | 2000 | 2005 | 2012 |
|---|---|---|---|
| Scanner | Optech ALTM-2010 | Optech ALTM-3033 | Leica ALS50-II |
| Wavelength | 1047 nm | 1064 nm | 1064 nm |
| Flying altitude | ca. 1000 m | ca. 2100 m | ca. 1600 m |
| Flying date | 10/06/2000 | 26/06/2005 | 15/09/2012 |
| Pulse repetition freq. | 10 kHz | 33 kHz | 144 kHz |
| Scan half angle | 10° | 20° | 10° |
| Max. no. of returns per pulse | 2 (first & last) | 2 (first & last) | 4 (first, second, third & last) |
| Post spacing | ca. 1 per 5 m2 | ca. 1 per 2 m2 | ca. 7.5 per 1 m2 |
| Footprint size | ca. 25 cm | ca. 45 cm | ca. 35 cm |
2.3. Data Extraction and Analysis
| Metric Name | Metric Description |
|---|---|
| Hmax | Maximum height |
| Hmean | Average height |
| Hstd | Standard deviation of height |
| H5, H10 ….H90, H95 | Height percentiles (H50 is median height) |
| Vegetation cover | Vegetation returns (>0.5 m) as a proportion of total returns |
| Canopy permeability | Proportion of laser pulses for which there are multiple returns |
| Canopy closure | Percentage of returns above a canopy height threshold of 2 m |
| Pgroundlayer | Percentage of returns in the ground layer (i.e., 0.5–2 m) |
| Punderstorey | Percentage of returns in the understorey layer (i.e., 2–8 m) |
| Poverstorey | Percentage of returns in the overstorey layer (i.e., >8 m) |
| Hmean > 2 m | Mean height of returns >2 m (i.e., mean height of the understorey & overstorey layers combined) |
| Hmean 2–8 m | Mean height of returns in the range 2–8 m (i.e., the understorey layer) |
| Hmean > 8 m | Mean height of returns >8 m (i.e., the overstorey layer) |
| Foliage height diversity (FHD) | Foliage height diversity calculated with the Shannon index as the proportion of returns in the ground layer, understorey and overstorey layers |
| Vegetation distribution ratio (VDR) | Vegetation distribution ratio (Hmax-H50/Hmax) |
3. Results
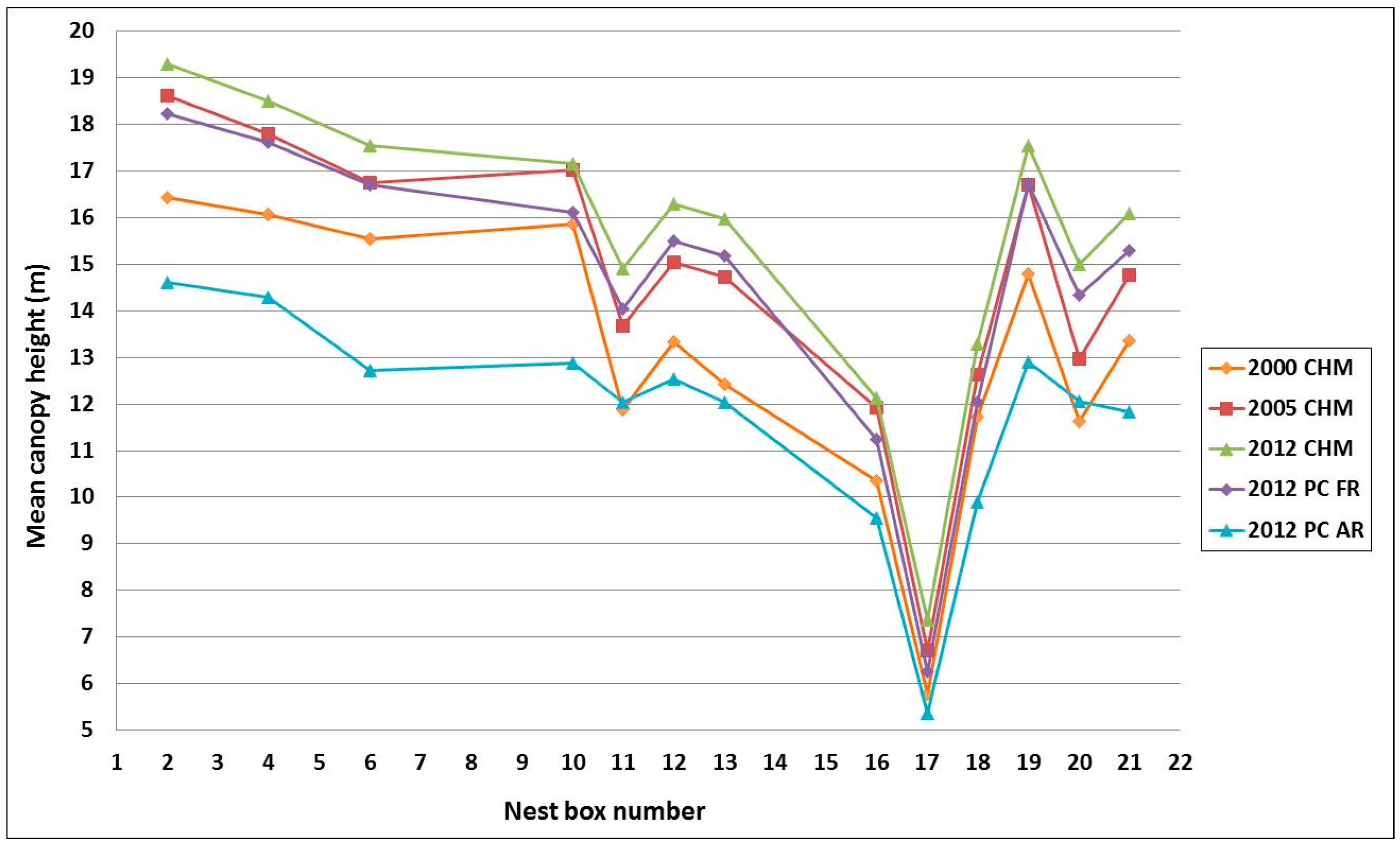
3.1. Comparison of Woodland Structure between the Different Lidar Datasets

3.2. Organism-Habitat Relationships Using Mean Height from 2000, 2005 and 2012 Chms
| Dataset | 1997 Great Tit Data | 2001 Great Tit Data | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hmean | Hmean > 2 m | Hmean > 8 m | Hmean | Hmean > 2 m | Hmean > 8 m | |||||||
| R2 | p | R2 | p | R2 | p | R2 | p | R2 | p | R2 | p | |
| 2000 CHM | 0.409 | 0.088 | 0.405 | 0.090 | 0.396 | 0.094 | 0.838 | <0.001 | 0.856 | <0.001 | 0.810 | <0.001 |
| 2005 CHM | 0.421 | 0.082 | 0.442 | 0.072 | 0.499 | 0.051 | 0.816 | <0.001 | 0.821 | <0.001 | 0.797 | <0.001 |
| 2012 CHM | 0.319 | 0.145 | 0.356 | 0.118 | 0.433 | 0.076 | 0.740 | <0.001 | 0.757 | <0.001 | 0.754 | <0.001 |
3.3. Organism–Habitat Relationships Using Mean Height from 2012 CHM and Point Cloud Data
| Dataset | 1997 Great Tit Data | 2001 Great Tit Data | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hmean | Hmean > 2 m | Hmean > 8 m | Hmean | Hmean > 2 m | Hmean > 8 m | |||||||
| R2 | p | R2 | p | R2 | p | R2 | p | R2 | p | R2 | p | |
| CHM | 0.319 | 0.145 | 0.356 | 0.118 | 0.433 | 0.076 | 0.740 | <0.001 | 0.757 | <0.001 | 0.754 | <0.001 |
| Point cloud | 0.280 | 0.177 | 0.327 | 0.139 | 0.400 | 0.093 | 0.718 | <0.001 | 0.744 | <0.001 | 0.748 | <0.001 |
3.4. Organism-Habitat Relationships Using Mean Height from 2012 Point Cloud Data Systematically Reducing the Point Density
3.5. Organism-Habitat Relationships Using 33 Structure Metrics from 2012 Point Cloud Data
| Metric Name | 1997 Great Tit Data | 2001 Great Tit Data | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| All Returns | First Return Only | All Returns | First Return Only | |||||||||
| Trend | R2 | p | Trend | R2 | p | Trend | R2 | p | Trend | R2 | p | |
| Veg. cover | − | 0.093 | 0.464 | − | 0.179 | 0.296 | − | 0.427 | 0.029 | − | 0.469 | 0.020 |
| Canopy perm. | + | 0.185 | 0.288 | + | 0.197 | 0.271 | − | 0.491 | 0.016 | − | 0.488 | 0.017 |
| Canopy closure | + | 0.222 | 0.234 | + | 0.002 | 0.921 | − | 0.440 | 0.026 | − | 0.473 | 0.019 |
| H5 | + | 0.000 | 0.985 | + | 0.000 | 0.990 | − | 0.111 | 0.316 | − | 0.511 | 0.013 |
| H10 | + | 0.001 | 0.952 | + | 0.025 | 0.708 | − | 0.113 | 0.311 | − | 0.643 | 0.003 |
| H15 | + | 0.002 | 0.919 | + | 0.105 | 0.433 | − | 0.261 | 0.108 | − | 0.684 | 0.002 |
| H20 | + | 0.016 | 0.760 | + | 0.174 | 0.304 | − | 0.386 | 0.041 | − | 0.705 | 0.001 |
| H25 | + | 0.042 | 0.627 | + | 0.222 | 0.239 | − | 0.484 | 0.018 | − | 0.723 | 0.001 |
| H30 | + | 0.108 | 0.471 | + | 0.264 | 0.192 | − | 0.580 | 0.006 | − | 0.731 | 0.001 |
| H35 | + | 0.136 | 0.370 | + | 0.312 | 0.150 | − | 0.635 | 0.003 | − | 0.736 | 0.001 |
| H40 | + | 0.166 | 0.317 | + | 0.403 | 0.091 | − | 0.679 | 0.002 | − | 0.744 | 0.001 |
| H45 | + | 0.203 | 0.261 | + | 0.453 | 0.067 | − | 0.714 | 0.001 | − | 0.747 | 0.001 |
| H50 (Hmedian) | + | 0.251 | 0.206 | + | 0.469 | 0.061 | − | 0.735 | 0.001 | − | 0.726 | 0.001 |
| H55 | + | 0.293 | 0.166 | + | 0.474 | 0.058 | − | 0.745 | 0.001 | − | 0.712 | 0.001 |
| H60 | + | 0.397 | 0.094 | + | 0.472 | 0.060 | − | 0.746 | 0.001 | − | 0.697 | 0.001 |
| H65 | + | 0.445 | 0.071 | + | 0.465 | 0.063 | − | 0.726 | 0.001 | − | 0.682 | 0.002 |
| H70 | + | 0.458 | 0.065 | + | 0.457 | 0.066 | − | 0.703 | 0.001 | − | 0.675 | 0.002 |
| H75 | + | 0.450 | 0.096 | + | 0.448 | 0.069 | − | 0.683 | 0.002 | − | 0.672 | 0.002 |
| H80 | + | 0.439 | 0.073 | + | 0.417 | 0.083 | − | 0.674 | 0.002 | − | 0.665 | 0.002 |
| H85 | + | 0.412 | 0.087 | + | 0.358 | 0.117 | − | 0.665 | 0.002 | − | 0.646 | 0.003 |
| H90 | + | 0.321 | 0.143 | + | 0.270 | 0.187 | − | 0.638 | 0.003 | − | 0.620 | 0.004 |
| H95 | + | 0.191 | 0.279 | + | 0.151 | 0.341 | − | 0.585 | 0.006 | − | 0.554 | 0.009 |
| H100 (Hmax) | + | 0.031 | 0.679 | + | 0.031 | 0.678 | − | 0.397 | 0.038 | − | 0.397 | 0.038 |
| Hmean | + | 0.284 | 0.174 | + | 0.280 | 0.177 | − | 0.661 | 0.002 | − | 0.718 | 0.001 |
| Hstd | + | 0.486 | 0.055 | + | 0.050 | 0.596 | − | 0.769 | 0.001 | + | 0.125 | 0.286 |
| Hmean >2m | + | 0.340 | 0.129 | + | 0.327 | 0.139 | − | 0.719 | 0.001 | − | 0.744 | 0.001 |
| Hmean 2-8m | − | 0.001 | 0.976 | − | 0.036 | 0.651 | + | 0.005 | 0.862 | + | 0.109 | 0.322 |
| Hmean >8m | + | 0.403 | 0.091 | + | 0.400 | 0.093 | − | 0.735 | 0.001 | − | 0.748 | 0.001 |
| Pgroundlayer | − | 0.153 | 0.338 | − | 0.118 | 0.404 | + | 0.416 | 0.032 | + | 0.483 | 0.018 |
| Punderstorey | − | 0.166 | 0.316 | − | 0.189 | 0.282 | + | 0.601 | 0.005 | + | 0.637 | 0.003 |
| Poverstorey | + | 0.221 | 0.240 | + | 0.136 | 0.368 | − | 0.528 | 0.011 | − | 0.577 | 0.007 |
| FHD | − | 0.045 | 0.612 | − | 0.082 | 0.491 | + | 0.502 | 0.015 | + | 0.589 | 0.006 |
| VDR | − | 0.149 | 0.345 | − | 0.344 | 0.126 | + | 0.674 | 0.002 | + | 0.713 | 0.001 |
4. Discussion
4.1. Great Tit Breeding Habitat Requirements
4.2. Assessment of Results against Study Aims
4.3. Applicability of Results to Other Ecological Systems
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- MacArthur, R.H.; MacArthur, J. On bird species diversity. Ecology 1961, 42, 594–598. [Google Scholar] [CrossRef]
- Turner, W.; Spector, S.; Gardiner, N.; Fladeland, M.; Sterling, E.; Steininger, M. Remote sensing for biodiversity science and conservation. Trends Ecol. Evol. 2003, 18, 306–314. [Google Scholar] [CrossRef]
- Bergen, K.M.; Goetz, S.J.; Dubayah, R.O.; Henebry, G.M.; Hunsaker, C.T.; Imhoff, M.L.; Nelson, R.F.; Parker, G.G.; Radeloff, V.C. Remote sensing of vegetation 3-D structure for biodiversity and habitat: Review and implications for lidar and radar spaceborne missions. J. Geophys. Res.: Biogeosci. 2009, 114. [Google Scholar] [CrossRef]
- Nelson, R.; Keller, C.; Ratnaswamy, M. Locating and estimating the extent of Delmarva fox squirrel habitat using an airborne LiDAR profiler. Remote Sens. Environ. 2005, 96, 292–301. [Google Scholar] [CrossRef]
- Coops, N.C.; Duffe, J.; Koot, C. Assessing the utility of lidar remote sensing technology to identify mule deer winter habitat. Can. J. Remote Sens. 2010, 36, 81–88. [Google Scholar] [CrossRef]
- Garabedian, J.E.; McGaughey, R.J.; Reutebuch, S.E.; Parresol, B.R.; Kilgo, J.C.; Moorman, C.E.; Peterson, M.N. Quantitative analysis of woodpecker habitat using high-resolution airborne LiDAR estimates of forest structure and composition. Remote Sens. Environ. 2014, 145, 68–80. [Google Scholar] [CrossRef]
- Vogeler, J.C.; Hudak, A.T.; Vierling, L.A.; Vierling, K.T. Lidar-derived canopy architecture predicts brown creeper occupancy of two western coniferous forests. Condor 2013, 115, 614–622. [Google Scholar] [CrossRef]
- Graf, R.F.; Mathys, L.; Bollmann, K. Habitat assessment for forest dwelling species using LiDAR remote sensing: Capercaillie in the Alps. For. Ecol. Manag. 2009, 257, 160–167. [Google Scholar] [CrossRef]
- Hinsley, S.A.; Hill, R.A.; Fuller, R.J.; Bellamy, P.E.; Rothery, P. Bird species distributions across woodland canopy structure gradients. Community Ecol. 2009, 10, 99–110. [Google Scholar] [CrossRef]
- Eldegard, K.; Dirksen, J.W.; Orka, H.O.; Halvorsen, R.; Naesset, E.; Gobakken, T.; Ohlson, M. Modelling bird richness and bird species presence in a boreal forest reserve using airborne laser-scanning and aerial images. Bird Study 2014, 61, 204–219. [Google Scholar] [CrossRef]
- Farrell, S.L.; Collier, B.A.; Skow, K.L.; Long, A.M.; Campomizzi, A.J.; Morrison, M.L.; Hays, K.B.; Wilkins, R.N. Using LiDAR-derived vegetation metrics for high-resolution, species distribution models for conservation planning. Ecosphere 2013, 4. [Google Scholar] [CrossRef]
- Zellweger, F.; Braunisch, V.; Baltensweiller, A.; Bollmann, K. Remotely sensed forest structural complexity predicts multi species occurrence at the landscape scale. For. Ecol. Manag. 2013, 307, 303–312. [Google Scholar] [CrossRef]
- Vierling, K.T.; Vierling, L.A.; Gould, W.A.; Martinuzzi, S.; Clawges, R.M. Lidar: Shedding new light on habitat characterization and modelling. Front. Ecol. Environ. 2008, 6, 90–98. [Google Scholar] [CrossRef]
- Bellamy, P.E.; Hill, R.A.; Rothery, P.; Hinsley, S.A.; Fuller, R.J.; Broughton, R.K. Willow warbler Phylloscopus trochilus habitat in woods with different structure and management in southern England. Bird Study 2009, 56, 338–348. [Google Scholar] [CrossRef]
- Clawges, R.; Vierling, K.; Vierling, L.; Rowell, E. The use of airborne lidar to assess avian species diversity, density, and occurrence in a pine/aspen forest. Remote Sens. Environ. 2008, 112, 2064–2073. [Google Scholar] [CrossRef]
- Swatantran, A.; Dubayah, R.; Goetz, S.; Hofton, M.; Betts, M.G.; Sun, M.; Simard, M.; Holmes, R. Mapping migratory bird prevalence using remote sensing data fusion. PLoS One 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Vierling, L.A.; Vierling, K.T.; Adam, P.; Hudak, A.T. Using satellite and airborne lidar to model woodpecker habitat occupancy at the landscape scale. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Palminteri, S.; Powell, G.V.N.; Asner, G.P.; Peres, C.C. LiDAR measurements of canopy structure predict spatial distribution of a tropical mature forest primate. Remote Sens. Environ. 2012, 127, 98–105. [Google Scholar] [CrossRef]
- Zhao, F.; Sweitzer, R.A.; Guo, Q.; Kelly, M. Characterizing habitats associated with fisher den structures in the Southern Sierra Nevada, California using discrete return lidar. For. Ecol. Manag. 2012, 280, 112–119. [Google Scholar] [CrossRef]
- Melin, M.; Packalen, P.; Matala, J.; Mehtalalo, L.; Pusenius, J. Assessing and modeling moose (Alces alces) habitats with airborne laser scanning data. Int. J. Appl. Earth Obs. 2013, 23, 389–396. [Google Scholar] [CrossRef]
- Flaherty, S.; Lurz, P.W.W.; Patenaude, G. Use of LiDAR in the conservation management of the endangered red squirrel (Sciurus vulgaris L.). J. Appl. Remote Sens. 2014, 8. [Google Scholar] [CrossRef]
- Jung, K.; Kaiser, S.; Bohm, S.; Nieschulze, J.; Kalko, E.K.V. Moving in three dimensions: Effects of structural complexity on occurrence and activity of insectivorous bats in managed forest stands. J. Appl. Ecol. 2012, 49, 523–531. [Google Scholar] [CrossRef]
- Müller, J.; Mehr, M.; Bässler, C.; Fenton, M.B.; Hothorn, T.; Pretzsch, H.; Klemmt, H.J.; Brandl, R. Aggregative response in bats: Prey abundance versus habitat. Oecologia 2012, 169, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Lone, K.; Loe, L.E.; Gobakken, T.; Linnell, J.D.C.; Odden, J.; Remmen, J.; Mysterud, A. Living and dying in a multi-predator landscape of fear: Roe deer are squeezed by contrasting pattern of predation risk imposed by lynx and humans. Oikos 2014, 123, 641–651. [Google Scholar] [CrossRef]
- Ewald, M.; Dupke, C.; Heurich, M.; Muller, J.; Reineking, B. LiDAR remote sensing of forest structure and GPS telemetry data provide insights on winter habitat selection of European roe deer. Forests 2014, 5, 1374–1390. [Google Scholar] [CrossRef]
- Melin, M.; Matala, J.; Mehtatalo, L.; Tiilikainen, R.; Tikkanen, O.P.; Maltamo, M.; Pusenius, J.; Packalen, P. Moose (Alces alces) reacts to high summer temperatures by utilizing thermal shelters in boreal forests—An analysis based on airborne laser scanning of the canopy structure at moose locations. Glob. Change Biol. 2014, 20, 1115–1125. [Google Scholar] [CrossRef]
- Hinsley, S.A.; Hill, R.A.; Gaveau, D.L.A.; Bellamy, P.E. Quantifying woodland structure and habitat quality for birds using airborne laser scanning. Funct. Ecol. 2002, 16, 851–857. [Google Scholar] [CrossRef]
- Hill, R.A.; Hinsley, S.A.; Gaveau, D.L.A.; Bellamy, P.E. Predicting habitat quality for Great Tits (Parus major) with airborne laser scanning data. Int. J. Remote Sens. 2004, 25, 4851–4855. [Google Scholar] [CrossRef]
- Hinsley, S.A.; Hill, R.A.; Bellamy, P.E.; Balzter, H. The application of lidar in woodland bird ecology: Climate, canopy structure, and habitat quality. Photogramm. Eng. Rem. Sens. 2006, 72, 1399–1406. [Google Scholar] [CrossRef]
- Seavy, N.E.; Viers, J.H.; Wood, J.K. Riparian bird response to vegetation structure: A multiscale analysis using LiDAR measurements of canopy height. Ecol. Appl. 2009, 19, 1848–1857. [Google Scholar] [CrossRef] [PubMed]
- Smart, L.S.; Swenson, J.J.; Christensen, N.L.; Sexton, J.O. Three-dimensional characterization of pine forest type and red-cockaded woodpecker habitat by small-footprint, discrete-return lidar. For. Ecol. Manag. 2012, 281, 100–110. [Google Scholar] [CrossRef]
- Goetz, S.J.; Steinberg, D.; Betts, M.G.; Holmes, R.T.; Doran, P.J.; Dubayah, R.; Hofton, M. Lidar remote sensing variables predict breeding habitat of a neotropical migrant bird. Ecology 2010, 91, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Vierling, K.T.; Swift, C.E.; Hudak, A.T.; Vogeler, J.C.; Vierling, L.A. Does the time lag between wildlife field data collection and LiDAR data acquisition matter for studies of wildlife distributions? A case study using bird communities. Remote Sens. Lett. 2014, 5, 185–193. [Google Scholar] [CrossRef]
- Patenaude, G.; Hill, R.A.; Milne, R.; Gaveau, D.L.A.; Briggs, B.B.J.; Dawson, T.P. Quantifying forest above ground carbon content using LiDAR remote sensing. Remote Sens. Environ. 2004, 93, 368–380. [Google Scholar] [CrossRef]
- Hill, R.A.; Thomson, A.G. Mapping woodland species composition and structure using airborne spectral and LiDAR data. Int. J. Rem. Sens. 2005, 26, 3763–3779. [Google Scholar] [CrossRef]
- Massey, M.E.; Welch, R.C. Monks Wood National Nature Reserve: The Experience of 40 Years 1953–1993; English Nature: Peterborough, UK, 1993. [Google Scholar]
- Hill, R.A.; Wilson, A.K.; George, M.; Hinsley, S.A. Mapping tree species in temperate deciduous woodland using time-series multi-spectral data. Appl. Veg. Sci. 2010, 13, 86–99. [Google Scholar] [CrossRef]
- Hill, R.A.; Broughton, R.K. Mapping the understorey of deciduous woodland from leaf-on and leaf-off airborne LiDAR data: A case study in lowland Britain. ISPRS J. Photogramm. 2009, 64, 223–233. [Google Scholar] [CrossRef]
- Hinsley, S.A.; Rothery, P.; Bellamy, P.E. Influence of woodland area on breeding success in Great Tits (Parus major) and Blue Tits (Parus caeruleus). J. Avian Biol. 1999, 3, 271–281. [Google Scholar] [CrossRef]
- Przybylo, R.; Wiggins, D.A.; Merila, J. Breeding success in Blue Tits: Good territories or good parents? J. Avian Biol. 2001, 32, 214–218. [Google Scholar] [CrossRef]
- Gaveau, D.L.A.; Hill, R.A. Quantifying canopy height underestimation by laser pulse penetration in small-footprint airborne laser scanning data. Can. J. Remote Sens. 2003, 29, 650–657. [Google Scholar] [CrossRef]
- Zhang, K.Q.; Chen, S.C.; Whitman, D.; Shyu, M.L.; Yan, J.H.; Zhang, C.C. A progressive morphological filter for removing non-ground measurements from airborne lidar data. IEEE Trans. Geosci. Remote Sens. 2003, 41, 872–882. [Google Scholar] [CrossRef]
- Moran, M.D. Arguments for rejecting the sequential Bonferroni in ecological studies. Oikos 2003, 100, 403–405. [Google Scholar] [CrossRef]
- Nakagawa, S. A farewell to Bonferroni: The problems of low statistical power and publication bias. Behav. Ecol. 2004, 15, 1044–1045. [Google Scholar] [CrossRef]
- Lack, D. Ecological Isolation in Birds; Blackwell Scientific Publications: Oxford, UK, 1971. [Google Scholar]
- Perrins, C.M. British Tits; Collins: London, UK, 1979. [Google Scholar]
- Gosler, A. The Great Tit; Hamlyn Limited: London, UK, 1993. [Google Scholar]
- Naef-Daenzer, B.; Keller, L.F. The foraging performance of great and blue tits (Parus major and P. caeruleus) in relation to caterpillar development and its consequences for nestling growth and fledging weight. J. Anim. Ecol. 1999, 57, 607–697. [Google Scholar]
- Minot, E.O. Effects of interspecific competition for food in breeding blue and great tits. J. Anim. Ecol. 1981, 50, 375–385. [Google Scholar] [CrossRef]
- Matthysen, E.; Adriaensen, F.; Dhondt, A.A. Multiple responses to increasing spring temperatures in the breeding cycle of blue and great tits (Cyanistes caeruleus, Parus. major). Glob. Change Biol. 2011, 17, 1–16. [Google Scholar] [CrossRef]
- Whitehouse, M.J.; Harrison, N.M.; Mackenzie, J.A.; Hinsley, S.A. Preferred habitat of breeding birds may be compromised by climate change: unexpected effects of an exceptionally cold, wet spring. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Wilsey, C.B.; Lawler, J.J.; Cimprich, D.A. Performance of habitat suitability models for the endangered black-capped vireo built with remotely-sensed data. Remote Sens. Environ. 2012, 119, 35–42. [Google Scholar] [CrossRef]
- Broughton, R.K.; Hill, R.A.; Freeman, S.N.; Hinsley, S.A. Describing habitat occupation by wood land birds with territory mapping and remotely sensed data: An example using the Marsh Tit (Poecile palustris). Condor 2012, 114, 812–822. [Google Scholar] [CrossRef] [Green Version]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hill, R.A.; Hinsley, S.A. Airborne Lidar for Woodland Habitat Quality Monitoring: Exploring the Significance of Lidar Data Characteristics when Modelling Organism-Habitat Relationships. Remote Sens. 2015, 7, 3446-3466. https://doi.org/10.3390/rs70403446
Hill RA, Hinsley SA. Airborne Lidar for Woodland Habitat Quality Monitoring: Exploring the Significance of Lidar Data Characteristics when Modelling Organism-Habitat Relationships. Remote Sensing. 2015; 7(4):3446-3466. https://doi.org/10.3390/rs70403446
Chicago/Turabian StyleHill, Ross A., and Shelley A. Hinsley. 2015. "Airborne Lidar for Woodland Habitat Quality Monitoring: Exploring the Significance of Lidar Data Characteristics when Modelling Organism-Habitat Relationships" Remote Sensing 7, no. 4: 3446-3466. https://doi.org/10.3390/rs70403446
APA StyleHill, R. A., & Hinsley, S. A. (2015). Airborne Lidar for Woodland Habitat Quality Monitoring: Exploring the Significance of Lidar Data Characteristics when Modelling Organism-Habitat Relationships. Remote Sensing, 7(4), 3446-3466. https://doi.org/10.3390/rs70403446