
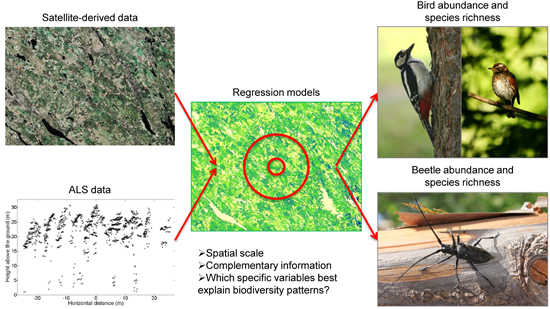
Can Airborne Laser Scanning (ALS) and Forest Estimates Derived from Satellite Images Be Used to Predict Abundance and Species Richness of Birds and Beetles in Boreal Forest?


Abstract
:
1. Introduction
2. Material and Methods
2.1. Study Area and Design

2.2. Beetle Sampling
2.3. Bird Sampling
2.4. Forest Estimates Derived from Satellite Images

{kind=link}
{kind=link}
| Variable | Description | Initial Raster Cell Size (m × m) |
|---|---|---|
| kNN-Based Variables | ||
| kNN_Age | Mean estimated forest age | 25 × 25 |
| kNN_Height | Mean estimated tree height | 25 × 25 |
| kNN_Pine | Mean estimated proportion of Scots pine stem volume | 25 × 25 |
| kNN_Spruce | Mean estimated proportion of Norway spruce stem volume | 25 × 25 |
| kNN_Deciduous | Mean estimated proportion of deciduous (i.e., broadleaved) tree stem volume | 25 × 25 |
| kNN_Volume | Mean estimated total stem volume | 25 × 25 |
| ALS-Based Variables | ||
| ALS_95Height | Mean of the 95th percentile of height above the ground | 10 × 10 |
| ALS_HighVeg | Mean of the fraction of returns ≥ 3 m above the ground of all returns | 10 × 10 |
| ALS_LowVeg | Mean of the fraction of returns ≥ 0.5 m above the ground of all returns ≤ 3 m above the ground | 10 × 10 |
| ALS_ShanH | Mean of Shannon’s diversity index for height | 10 × 10 |
| ALS_MaxH | Mean of the maximum height | 1 × 1 |
| ALS_MaxHsd | Standard deviation of the maximum height | 1 × 1 |
2.5. ALS Data
- The 95th percentile of vegetation height above the ground (95Height). This variable depicts a general measure of the canopy height.
- The fraction of returns ≥ 3 m above the ground of all returns (HighVeg). This represents a general measure of higher-level foliage density, i.e., excluding vegetation below 3 m.
- The fraction of returns ≥ 0.5 m above the ground of all returns ≤ 3 m above the ground (LowVeg). This represents a general measure of lower-level foliage density below 3.0 m.
- Shannon’s diversity index for the proportion of returns in height intervals 0.5–3 m, 3–10 m and 10–35 m above the ground within each raster cell (ShanH). This provides an index of foliage height diversity (sensu [63]).
2.6. Regression Models
| 50-m Radius | 200-m Radius |
|---|---|
| ALS_ShanH 50 | ALS_ShanH 200 |
| ALS_LowVeg 50 | ALS_LowVeg 200 |
| ALS_MaxH 50 or kNN_Volume 50 | ALS_MaxH 200 or kNN_Volume 200 |
| kNN_Deciduous 50 | kNN_Height 200 |
| kNN_Pine 50 | kNN_Deciduous 200 |
| kNN_Pine 200 |
3. Results
| 50 m Radius | 200 m Radius | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Regression Model | Adjusted R2 | AICc | RMSE (Cross-Validation) | Bias (Cross-Validation) | Regression Model | Adjusted R2 | AICc | RMSE (Cross-Validation) | Bias (Cross-Validation) | |
| Bird abundance | +ALS_LowVeg50 *** +ALS_MaxH50 *** | 0.42 | 33.6 | 35.8% | −16.7% | +ALS_LowVeg200 * +ALS_MaxH200 *** | 0.21 | 48.0 | 37.9% | −16.7% |
| Bird species richness | +ALS_LowVeg50 ** +ALS_MaxH50 *** | 0.41 | 29.6 | 34.8% | −17.4% | +ALS_LowVeg200 * +ALS_MaxH200 ** | 0.20 | 44.1 | 36.8% | −17.5% |
| Flying beetle abundance | −kNN_Pine50 * +ALS_MaxH50 *** | 0.53 | 33.1 | 39.6% | −0.2% | −kNN_Pine200 * + ALS_MaxH200 ** | 0.38 | 42.6 | 45.7% | 0.0% |
| Flying beetle species richness | −kNN_Pine50 * +ALS_MaxH50 *** | 0.47 | 6.2 | 23.5% | −0.6% | −kNN_Pine200 ns +ALS_MaxH200 ** | 0.28 | 16.2 | 26.9% | −0.7% |
| Epigaeic beetle abundance | +ALS_MaxH50 *** | 0.53 | 73.9 | 77.5% | −0.5% | +kNN_Volume200 *** | 0.45 | 78.9 | 83.0% | −0.5% |
| Epigaeic beetle species richness | −kNN_Pine50 * +ALS_MaxH50 *** | 0.59 | 29.5 | 32.9% | −1.2% | −kNN_Pine200 ** + ALS_MaxH200 *** | 0.57 | 30.3 | 36.5% | −0.8% |
| kNN_Deciduous50 | kNN_Pine50 | ALS_MaxH50 | ALS_LowVeg50 | ALS_ShanH50 | |
|---|---|---|---|---|---|
| Bird abundance | 0.26 (−) | 0.27 (−) | 1.00 (+) | 0.99 (+) | 0.22 (−) |
| Bird species richness | 0.27 (−) | 0.23 (−) | 1.00 (+) | 0.98 (+) | 0.22 (−) |
| Flying beetle abundance | 0.23 (+) | 0.89 (−) | 0.99 (+) | 0.43 (−) | 0.23 (+) |
| Flying beetle species richness | 0.21 (+) | 0.72 (−) | 0.99 (+) | 0.24 (−) | 0.22 (−) |
| Epigaeic beetle abundance | 0.26 (+) | 0.34 (−) | 1.00 (+) | 0.25 (+) | 0.23 (−) |
| Epigaeic beetle species richness | 0.22 (+) | 0.91 (−) | 0.99 (+) | 0.22 (+) | 0.70 (−) |
| kNN_Deciduous50 | kNN_Pine50 | kNN_Volume50 | ALS_LowVeg50 | ALS_ShanH50 | |
|---|---|---|---|---|---|
| Bird abundance | 0.30 (−) | 0.23 (−) | 1.00 (+) | 0.73 (+) | 0.23 (+) |
| Bird species richness | 0.31 (−) | 0.22 (+) | 1.00 (+) | 0.68 (+) | 0.23 (−) |
| Flying beetle abundance | 0.21 (+) | 0.71 (−) | 0.92 (+) | 0.84 (−) | 0.27 (+) |
| Flying beetle species richness | 0.23 (−) | 0.55 (−) | 0.84 (+) | 0.54 (−) | 0.32 (−) |
| Epigaeic beetle abundance | 0.20 (+) | 0.22 (−) | 1.00 (+) | 0.25 (−) | 0.21 (−) |
| Epigaeic beetle species richness | 0.20 (+) | 0.46 (−) | 0.98 (+) | 0.24 (−) | 0.31 (−) |
4. Discussion
4.1. Can ALS and Satellite-Derived Data Products Be Used to Identify Species Richness and Abundance Hotspots for Beetles and Birds in Managed Boreal Forest?
4.2. Do the Models Perform Better When the Explanatory Variables Are Derived at the Scale of Homogenous Forest Stands or at a Scale Including Also Parts of Adjacent Stands?
4.3. Do ALS and Satellite-Derived Data Products Provide Complementary Types of Information for Predicting Biodiversity Patterns?
4.4. Which Specific Variables Derived from These Two Remote Sensing Sources Can Best Explain Biodiversity Patterns for Beetle and Bird Species in Managed Boreal Forests?
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Harrison, S.; Bruna, E. Habitat fragmentation and large-scale conservation: What do we know for sure? Ecography 1999, 22, 225–232. [Google Scholar] [CrossRef]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344. [Google Scholar] [CrossRef]
- Esseen, P.-A.; Ehnström, B.; Ericson, L.; Sjöberg, K. Boreal forests. In Ecological Bulletins, Boreal Ecosystems and Landscapes: Structures, Processes and Conservation of Biodiversity; Hansson, L., Ed.; Oikos Editorial Office: Lund, Sweden, 1997; Volume 46, pp. 16–47. [Google Scholar]
- Linder, P.; Östlund, L. Structural changes in three mid-boreal Swedish forest landscapes, 1885–1996. Biol. Conserv. 1998, 85, 9–19. [Google Scholar] [CrossRef]
- Siitonen, J. Forest management, coarse woody debris and saproxylic organisms: Fennoscandian boreal forest as an example. Ecol. Bull. 2001, 49, 11–41. [Google Scholar]
- Gibb, H.; Ball, J.P.; Johansson, T.; Atlegrim, O.; Hjältén, J.; Danell, K. Effects of management on coarse woody debris volume and composition in boreal forests in northern Sweden. Scand. J. For. Res. 2005, 20, 213–222. [Google Scholar] [CrossRef]
- Grove, S.J. Saproxylic insect ecology and the sustainable management of forests. In Annual Review of Ecology and Systematics; Futuyma, D.J., Ed.; Annual Reviews: Palo Alto, CA, USA, 2002; Volume 33, pp. 1–23. [Google Scholar]
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 1986, 15, 133–302. [Google Scholar]
- Niklasson, M.; Granström, A. Numbers and sizes of fires: Long-term spatially explicit fire history in a Swedish boreal landscape. Ecology 2000, 81, 1484–1499. [Google Scholar] [CrossRef]
- Larsson, S.; Danell, K. Science and the management of boreal forest biodiversity—Preface. Scand. J. For. Res. 2001, 16, 5–9. [Google Scholar] [CrossRef]
- Angelstam, P.; Roberge, J.M.; Ek, T.; Laestadius, L. Data and tools for conservation, management, and restoration of northern forest ecosystems at multiple scales. In Restoration of Boreal and Temperate Forests; Stanturf, J., Madsen, P., Eds.; CRC Press: Boca Raton, FL, USA, 2005; Volume 3, pp. 269–283. [Google Scholar]
- Lindenmayer, D.B.; Franklin, J.F.; Fischer, J. General management principles and a checklist of strategies to guide forest biodiversity conservation. Biol. Conserv. 2006, 131, 433–445. [Google Scholar] [CrossRef]
- Johansson, T.; Hjältén, J.; de Jong, J.; von Stedingk, H. Environmental considerations from legislation and certification in managed forest stands: A review of their importance for biodiversity. For. Ecol. Manag. 2013, 303, 98–112. [Google Scholar] [CrossRef]
- Gibb, H.; Hjältén, J.; Ball, J.P.; Atlegrim, O.; Pettersson, R.B.; Hilszczanski, J.; Johansson, T.; Danell, K. Effects of landscape composition and substrate availability on saproxylic beetles in boreal forests: A study using experimental logs for monitoring assemblages. Ecography 2006, 29, 191–204. [Google Scholar] [CrossRef]
- Paillet, Y.; Berges, L.; Hjältén, J.; Odor, P.; Avon, C.; Bernhardt-Roemermann, M.; Bijlsma, R.-J.; de Bruyn, L.; Fuhr, M.; Grandin, U.; et al. Biodiversity differences between managed and unmanaged forests: Meta-analysis of species richness in Europe. Conserv. Biol. 2010, 24, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Stighäll, K.; Roberge, J.-M.; Andersson, K.; Angelstam, P. Usefulness of biophysical proxy data for modelling habitat of an endangered forest species: The white-backed woodpecker dendrocopos leucotos. Scand. J. For. Res. 2011, 26, 576–585. [Google Scholar] [CrossRef]
- Reese, H.; Nilsson, M.; Pahen, T.G.; Hagner, O.; Joyce, S.; Tingelof, U.; Egberth, M.; Olsson, H. Countrywide estimates of forest variables using satellite data and field data from the national forest inventory. Ambio 2003, 32, 542–548. [Google Scholar] [PubMed]
- Martinuzzi, S.; Vierling, L.A.; Gould, W.A.; Vierling, K.T. Improving the characterization and mapping of wildlife habitats with lidar data: Measurement priorities for the inland northwest, USA. Gap Anal. Bull. 2009, 16, 1–8. [Google Scholar]
- Davies, A.B.; Asner, G.P. Advances in animal ecology from 3D-LiDAR ecosystem mapping. Trends Ecol. Evolut. 2014, 29, 681–691. [Google Scholar] [CrossRef]
- Shan, J.; Toth, C. Topographic Laser Ranging and Scanning: Principles and Processing; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2009. [Google Scholar]
- Wulder, M.A.; White, J.C.; Nelson, R.F.; Næsset, E.; Orka, H.O.; Coops, N.C.; Hilker, T.; Bater, C.W.; Gobakken, T. LiDAR sampling for large-area forest characterization: A review. Remote Sens. Environ. 2012, 121, 196–209. [Google Scholar] [CrossRef]
- Ussyshkin, V.; Theriault, L. Airborne LiDAR: Advances in discrete return technology for 3D vegetation mapping. Remote Sens. 2011, 3, 416–434. [Google Scholar] [CrossRef]
- Kraus, K.; Pfeifer, N. Determination of terrain models in wooded areas with airborne laser scanner data. ISPRS J. Photogram. Remote Sens. 1998, 53, 193–203. [Google Scholar] [CrossRef]
- Martinuzzi, S.; Vierling, L.A.; Gould, W.A.; Falkowski, M.J.; Evans, J.S.; Hudak, A.T.; Vierling, K.T. Mapping snags and understory shrubs for a LiDAR-based assessment of wildlife habitat suitability. Remote Sens. Environ. 2009, 113, 2533–2546. [Google Scholar] [CrossRef]
- Hyyppä, J.; Hyyppä, H.; Leckie, D.; Gougeon, F.; Yu, X.; Maltamo, M. Review of methods of small-footprint airborne laser scanning for extracting forest inventory data in boreal forests. Int. J. Remote Sens. 2008, 29, 1339–1366. [Google Scholar] [CrossRef]
- Hudak, A.T.; Evans, J.S.; Smith, A.M.S. LiDAR utility for natural resource managers. Remote Sens. 2009, 1, 934–951. [Google Scholar] [CrossRef]
- Stilla, U.; Jutzi, B. Waveform Analysis for Small-Footprint Pulsed Laser Systems. In Topographic Laser Ranging and Scanning: Principles and Processing; Shan, J., Toth, C., Eds.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2009. [Google Scholar]
- Reitberger, J.; Schnorr, C.; Krzystek, P.; Stilla, U. 3D segmentation of single trees exploiting full waveform lidar data. ISPRS J. Photogram. Remote Sens. 2009, 64, 561–574. [Google Scholar] [CrossRef]
- Lindberg, E.; Eysn, L.; Hollaus, M.; Holmgren, J.; Pfeifer, N. Delineation of tree crowns and tree species classification from full-waveform airborne laser scanning data using 3-D ellipsoidal clustering. IEEE J. Sel. Top. Appl. Earth Observ. Remote Sens. 2014, 7, 3174–3181. [Google Scholar] [CrossRef]
- Polewski, P.; Yao, W.; Heurich, M.; Krzystek, P.; Stilla, U. Detection of Fallen Trees in als Point Clouds by Learning the Normalized Cut Similarity Function from Simulated Samples. ISPRS Ann. Photogramm. Remote Sens. Spatial Inf. Sci. 2014, II-3, 111–118. [Google Scholar] [CrossRef]
- Whitehurst, A.S.; Swatantran, A.; Blair, J.B.; Hofton, M.A.; Dubayah, R. Characterization of canopy layering in forested ecosystems using full waveform LiDAR. Remote Sens. 2013, 5, 2014–2036. [Google Scholar] [CrossRef]
- Camprodon, J.; Brotons, L. Effects of undergrowth clearing on the bird communities of the northwestern mediterranean coppice holm oak forests. For. Ecol. Manag. 2006, 221, 72–82. [Google Scholar] [CrossRef]
- Eggers, S.; Low, M. Differential demographic responses of sympatric parids to vegetation management in boreal forest. For. Ecol. Manag. 2014, 319, 169–175. [Google Scholar] [CrossRef]
- Stenbacka, F.; Hjältén, J.; Hilszczanski, J.; Dynesius, M. Saproxylic and non-saproxylic beetle assemblages in boreal spruce forests of different age and forestry intensity. Ecol. Appl. 2010, 20, 2310–2321. [Google Scholar] [CrossRef] [PubMed]
- Niemelä, J.; Haila, Y.; Punttila, P. The importance of small-scale heterogeneity in boreal forests: Variation in diversity in forest-floor invertebrates across the succession gradient. Ecography 1996, 19, 352–368. [Google Scholar] [CrossRef]
- Seavy, N.E.; Viers, J.H.; Wood, J.K. Riparian bird response to vegetation structure: A multiscale analysis using LiDAR measurements of canopy height. Ecol. Appl. 2009, 19, 1848–1857. [Google Scholar] [CrossRef] [PubMed]
- Zellweger, F.; Braunisch, V.; Baltensweiler, A.; Bollmann, K. Remotely sensed forest structural complexity predicts multi species occurrence at the landscape scale. For. Ecol. Manag. 2013, 307, 303–312. [Google Scholar] [CrossRef]
- Coops, N.C.; Waring, R.H.; Wulder, M.A.; Pidgeon, A.M.; Radeloff, V.C. Bird diversity: A predictable function of satellite-derived estimates of seasonal variation in canopy light absorbance across the United States. J. Biogeogr. 2009, 36, 905–918. [Google Scholar] [CrossRef]
- Coops, N.C.; Wulder, M.A.; Iwanicka, D. Exploring the relative importance of satellite-derived descriptors of production, topography and land cover for predicting breeding bird species richness over Ontario, Canada. Remote Sens. Environ. 2009, 113, 668–679. [Google Scholar] [CrossRef]
- Goetz, S.J.; Sun, M.; Zolkos, S.; Hansen, A.; Dubayah, R. The relative importance of climate and vegetation properties on patterns of north American breeding bird species richness. Environ. Res. Lett. 2014, 9. [Google Scholar] [CrossRef]
- Sheeren, D.; Bonthoux, S.; Balent, G. Modeling bird communities using unclassified remote sensing imagery: Effects of the spatial resolution and data period. Ecol. Indic. 2014, 43, 69–82. [Google Scholar] [CrossRef]
- Shirley, S.M.; Yang, Z.; Hutchinson, R.A.; Alexander, J.D.; McGarigal, K.; Betts, M.G. Species distribution modelling for the people: Unclassified landsat tm imagery predicts bird occurrence at fine resolutions. Divers. Distrib. 2013, 19, 855–866. [Google Scholar] [CrossRef]
- Culbert, P.D.; Radeloff, V.C.; St-Louis, V.; Flather, C.H.; Rittenhouse, C.D.; Albright, T.P.; Pidgeon, A.M. Modeling broad-scale patterns of avian species richness across the midwestern United States with measures of satellite image texture. Remote Sens. Environ. 2012, 118, 140–150. [Google Scholar] [CrossRef]
- St-Louis, V.; Pidgeon, A.M.; Kuemmerle, T.; Sonnenschein, R.; Radeloff, V.C.; Clayton, M.K.; Locke, B.A.; Bash, D.; Hostert, P. Modelling avian biodiversity using raw, unclassified satellite imagery. Philos. Trans. R. Soc. B-Biol. Sci. 2014, 369. [Google Scholar] [CrossRef]
- Wood, E.M.; Pidgeon, A.M.; Radeloff, V.C.; Keuler, N.S. Image texture predicts avian density and species richness. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Goetz, S.; Steinberg, D.; Dubayah, R.; Blair, B. Laser remote sensing of canopy habitat heterogeneity as a predictor of bird species richness in an eastern temperate forest, USA. Remote Sens. Environ. 2007, 108, 254–263. [Google Scholar] [CrossRef]
- Simonson, W.D.; Allen, H.D.; Coomes, D.A. Applications of airborne lidar for the assessment of animal species diversity. Methods Ecol. Evol. 2014, 5, 719–729. [Google Scholar] [CrossRef]
- Hill, R.A.; Hinsley, S.A.; Broughton, R.K. Assessing habitats and organism-habitat relationships by airborne laser scanning. In Forestry Applications of Airborne Laser Scanning: Concepts and Case Studies; Maltamo, M., Naesset, E., Vauhkonen, J., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 27, pp. 335–356. [Google Scholar]
- Müller, J.; Vierling, K. Assessing biodiversity by airborne laser scanning. In Forestry Applications of Airborne Laser Scanning: Concepts and Case Studies; Maltamo, M., Naesset, E., Vauhkonen, J., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 27, pp. 357–374. [Google Scholar]
- Weisberg, P.J.; Dilts, T.E.; Becker, M.E.; Young, J.S.; Wong-Kone, D.C.; Newton, W.E.; Ammon, E.M. Guild-specific responses of avian species richness to lidar-derived habitat heterogeneity. Acta Oecol. 2014, 59, 72–83. [Google Scholar] [CrossRef]
- Vogeler, J.C.; Hudak, A.T.; Vierling, L.A.; Evans, J.; Green, P.; Vierling, K.I.T. Terrain and vegetation structural influences on local avian species richness in two mixed-conifer forests. Remote Sens. Environ. 2014, 147, 13–22. [Google Scholar] [CrossRef]
- Eldegard, K.; Dirksen, J.W.; Ørka, H.O.; Halvorsen, R.; Næsset, E.; Gobakken, T.; Ohlson, M. Modelling bird richness and bird species presence in a boreal forest reserve using airborne laser-scanning and aerial images. Bird Study 2014, 61, 204–219. [Google Scholar] [CrossRef]
- Müller, J.; Moning, C.; Bässler, C.; Heurich, M.; Brandl, R. Using airborne laser scanning to model potential abundance and assemblages of forest passerines. Basic Appl. Ecol. 2009, 10, 671–681. [Google Scholar] [CrossRef]
- Culbert, P.D.; Radeloff, V.C.; Flather, C.H.; Kellndorfer, J.M.; Rittenhouse, C.D.; Pidgeon, A.M. The influence of vertical and horizontal habitat structure on nationwide patterns of avian biodiversity. Auk 2013, 130, 656–665. [Google Scholar] [CrossRef]
- Muukkonen, P.; Angervuori, A.; Virtanen, T.; Kuparinen, A.; Merila, J. Loss and fragmentation of siberian jay (perisoreus infaustus) habitats. Boreal Environ. Res. 2012, 17, 59–71. [Google Scholar]
- Sirkia, S.; Helle, P.; Linden, H.; Nikula, A.; Norrdahl, K.; Suorsa, P.; Valkeajarvi, P. Persistence of capercaillie (tetrao urogallus) lekking areas depends on forest cover and fine-grain fragmentation of boreal forest landscapes. Ornis Fenn. 2011, 88, 14–29. [Google Scholar]
- Huang, Q.; Swatantran, A.; Dubayah, R.; Goetz, S.J. The influence of vegetation height heterogeneity on forest and woodland bird species richness across the United States. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Ahti, T.; Hämet-Ahti, L.; Jalas, J. Vegetation zones and their sections in northwestern Europe. Ann. Bot. Fenn. 1968, 5, 169–211. [Google Scholar]
- Arnborg, T. Forest types of northern Sweden. Vegetatio 1990, 90, 1–13. [Google Scholar] [CrossRef]
- Johansson, T.; Andersson, J.; Hjältén, J.; Dynesius, M.; Ecke, F. Short-term responses of beetle assemblages to wildfire in a region with more than 100 years of fire suppression. Insect Conserv. Divers. 2011, 4, 142–151. [Google Scholar] [CrossRef]
- Silfverberg, H. Enumeratio nova Coleopterorum Fennoscandiae, Daniae et Baltiae. Sahlbergia 2004, 9, 1–111, in English. [Google Scholar]
- Egberth, M.; Nilsson, M.; Axensten, P. kNN-Sweden—Current map data on forest land. Available online: http://skogskarta.slu.se/index.cfm?eng=1 (accessed on 21 January 2014).
- MacArthur, R.; MacArthur, J.W. On bird species diversity. Ecology 1961, 42, 594–598. [Google Scholar] [CrossRef]
- Bellamy, P.E.; Hill, R.A.; Rothery, P.; Hinsley, S.A.; Fuller, R.J.; Broughton, R.K. Willow warbler phylloscopus trochilus habitat in woods with different structure and management in southern England. Bird Study 2009, 56, 338–348. [Google Scholar] [CrossRef]
- Hinsley, S.A.; Hill, R.A.; Bellamy, P.E.; Harrison, N.M.; Speakman, J.R.; Wilson, A.K.; Ferns, P.N. Effects of structural and functional habitat gaps on breeding woodland birds: Working harder for less. Landsc. Ecol. 2008, 23, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.; Brandl, R. Assessing biodiversity by remote sensing in mountainous terrain: The potential of lidar to predict forest beetle assemblages. J. Appl. Ecol. 2009, 46, 897–905. [Google Scholar] [CrossRef]
- Müller, J.; Stadler, J.; Brandl, R. Composition versus physiognomy of vegetation as predictors of bird assemblages: The role of lidar. Remote Sens. Environ. 2010, 114, 490–495. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002; p. 488. [Google Scholar]
- Geisser, S. Predictive Inference: An Introduction; Chapman & Hall: New York, NY, USA, 1993. [Google Scholar]
- Clawges, R.; Vierling, K.; Vierling, L.; Rowell, E. The use of airborne lidar to assess avian species diversity, density, and occurrence in a pine/aspen forest. Remote Sens. Environ. 2008, 112, 2064–2073. [Google Scholar] [CrossRef]
- Ranius, T.; Martikainen, P.; Kouki, J. Colonisation of ephemeral forest habitats by specialised species: Beetles and bugs associated with recently dead aspen wood. Biodivers. Conserv. 2011, 20, 2903–2915. [Google Scholar] [CrossRef]
- Sverdrup-Thygeson, A.; Gustafsson, L.; Kouki, J. Spatial and temporal scales relevant for conservation of dead-wood associated species: Current status and perspectives. Biodivers. Conserv. 2014, 23, 513–535. [Google Scholar] [CrossRef]
- Wells, K.; Boehm, S.M.; Boch, S.; Fischer, M.; Kalko, E.K.V. Local and landscape-scale forest attributes differ in their impact on bird assemblages across years in forest production landscapes. Basic Appl. Ecol. 2011, 12, 97–106. [Google Scholar] [CrossRef]
- Andersson, J.; Hjältén, J.; Dynesius, M. Long-term effects of stump harvesting and landscape composition on beetle assemblages in the hemiboreal forest of Sweden. For. Ecol. Manag. 2012, 271, 75–80. [Google Scholar] [CrossRef]
- Azeria, E.T.; Fortin, D.; Hebert, C.; Peres-Neto, P.; Pothier, D.; Ruel, J.-C. Using null model analysis of species co-occurrences to deconstruct biodiversity patterns and select indicator species. Divers. Distrib. 2009, 15, 958–971. [Google Scholar] [CrossRef]
- Törmä, M. Estimation of tree Species Proportions of Forest Stands Using Laser Scanning. ISPRS Int. Arch. Photogramm. Remote Sens. Spatial Inf. Sci. 2000, XXXIII((Part B7)), 1524–1531. [Google Scholar]
- Packalén, P.; Maltamo, M. The k-msn method for the prediction of species-specific stand attributes using airborne laser scanning and aerial photographs. Remote Sens. Environ. 2007, 109, 328–341. [Google Scholar] [CrossRef]
- Elo, M.; Roberge, J.-M.; Rajasarkka, A.; Monkkonen, M. Energy density and its variation in space limit species richness of boreal forest birds. J. Biogeogr. 2012, 39, 1462–1472. [Google Scholar] [CrossRef]
- Gibb, H.; Johansson, T.; Stenbacka, F.; Hjältén, J. Functional roles affect diversity-succession relationships for boreal beetles. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Hjältén, J.; Stenbacka, F.; Pettersson, R.B.; Gibb, H.; Johansson, T.; Danell, K.; Ball, J.P.; Hilszczanski, J. Micro and macro-habitat associations in saproxylic beetles: Implications for biodiversity management. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Roberge, J.-M.; Angelstam, P.; Villard, M.-A. Specialised woodpeckers and naturalness in hemiboreal forests—Deriving quantitative targets for conservation planning. Biol. Conserv. 2008, 141, 997–1012. [Google Scholar] [CrossRef]
- Janssen, P.; Fortin, D.; Hebert, C. Beetle diversity in a matrix of old-growth boreal forest: Influence of habitat heterogeneity at multiple scales. Ecography 2009, 32, 423–432. [Google Scholar] [CrossRef]
- Helle, P. Effects of forest regeneration on the structure of bird communities in northern Finland. Holarct. Ecol. 1985, 8, 120–132. [Google Scholar]
- Lesak, A.A.; Radeloff, V.C.; Hawbaker, T.J.; Pidgeon, A.M.; Gobakken, T.; Contrucci, K. Modeling forest songbird species richness using LiDAR-derived measures of forest structure. Remote Sens. Environ. 2011, 115, 2823–2835. [Google Scholar] [CrossRef]
- Jones, T.G.; Arcese, P.; Sharma, T.; Coops, N.C. Describing avifaunal richness with functional and structural bioindicators derived from advanced airborne remotely sensed data. Int. J. Remote Sens. 2013, 34, 2689–2713. [Google Scholar] [CrossRef]
- Dahlberg, A.; Stokland, J.N. Vedlevande Arters Krav på Substrat—en Sammanställning Och Analys av 3600 Arter (in Swedish with English summary: Substrate Requirements of Wood-Inhabiting Species—A Synthesis and Analysis of 3600 Species); Skogsstyrelsen: Jönköping, Sweden, 2004. [Google Scholar]
- Bernes, C. Biologisk Mångfald i Sverige. Monitor 22 (Biodiversity in Sweden. Monitor 22); Naturvårdsverket/The Swedish Environment Protection Agency: Stockholm, Sweden, 2011. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lindberg, E.; Roberge, J.-M.; Johansson, T.; Hjältén, J. Can Airborne Laser Scanning (ALS) and Forest Estimates Derived from Satellite Images Be Used to Predict Abundance and Species Richness of Birds and Beetles in Boreal Forest? Remote Sens. 2015, 7, 4233-4252. https://doi.org/10.3390/rs70404233
Lindberg E, Roberge J-M, Johansson T, Hjältén J. Can Airborne Laser Scanning (ALS) and Forest Estimates Derived from Satellite Images Be Used to Predict Abundance and Species Richness of Birds and Beetles in Boreal Forest? Remote Sensing. 2015; 7(4):4233-4252. https://doi.org/10.3390/rs70404233
Chicago/Turabian StyleLindberg, Eva, Jean-Michel Roberge, Therese Johansson, and Joakim Hjältén. 2015. "Can Airborne Laser Scanning (ALS) and Forest Estimates Derived from Satellite Images Be Used to Predict Abundance and Species Richness of Birds and Beetles in Boreal Forest?" Remote Sensing 7, no. 4: 4233-4252. https://doi.org/10.3390/rs70404233
APA StyleLindberg, E., Roberge, J. -M., Johansson, T., & Hjältén, J. (2015). Can Airborne Laser Scanning (ALS) and Forest Estimates Derived from Satellite Images Be Used to Predict Abundance and Species Richness of Birds and Beetles in Boreal Forest? Remote Sensing, 7(4), 4233-4252. https://doi.org/10.3390/rs70404233