Grassland Phenology Response to Drought in the Canadian Prairies

Abstract
:
1. Introduction
- Is drought a significant climatic factor for the temporal evolution of grassland phenology?
- How have changes in precipitation influenced the grassland phenology response to drought?
2. Materials and Methods
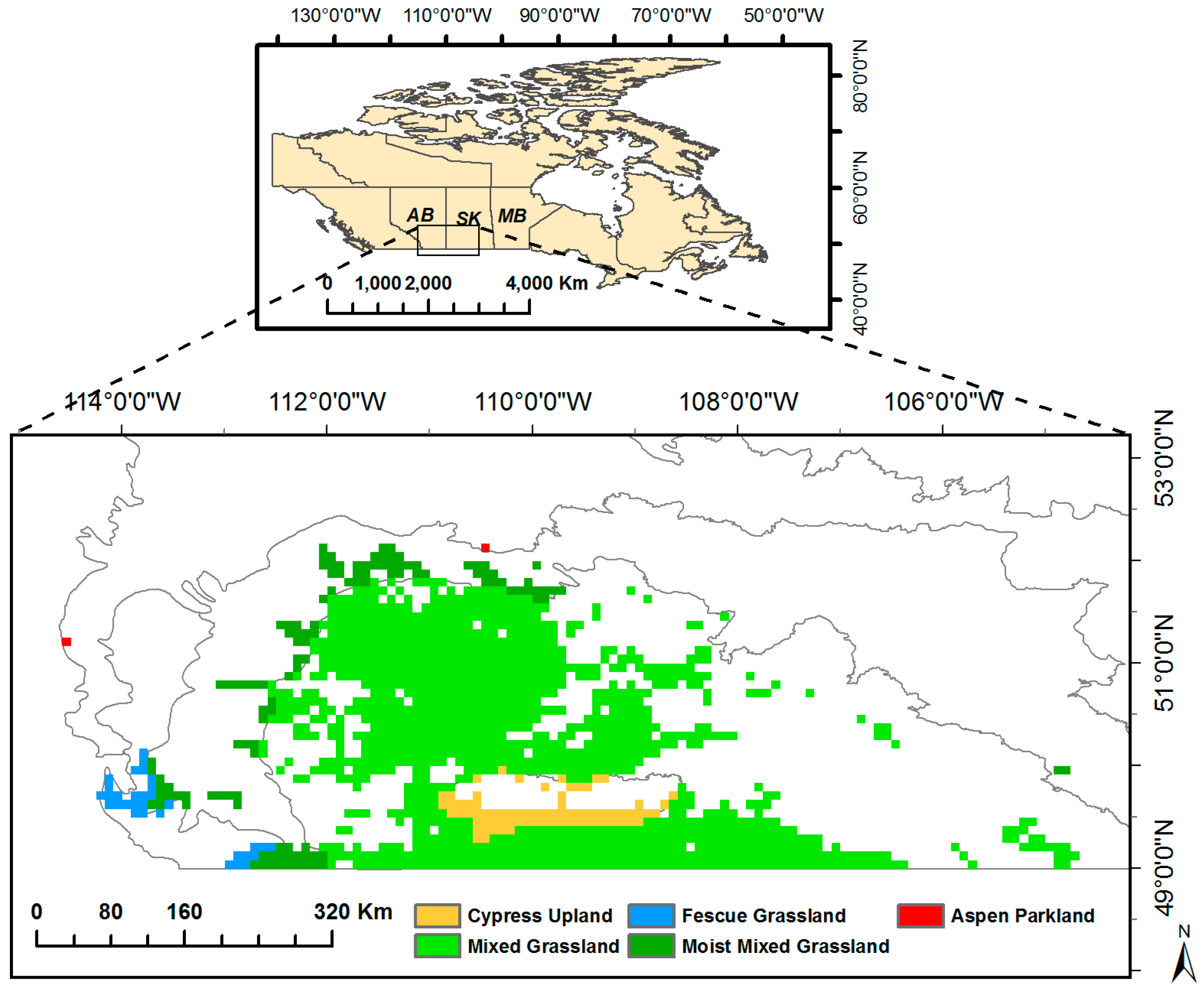
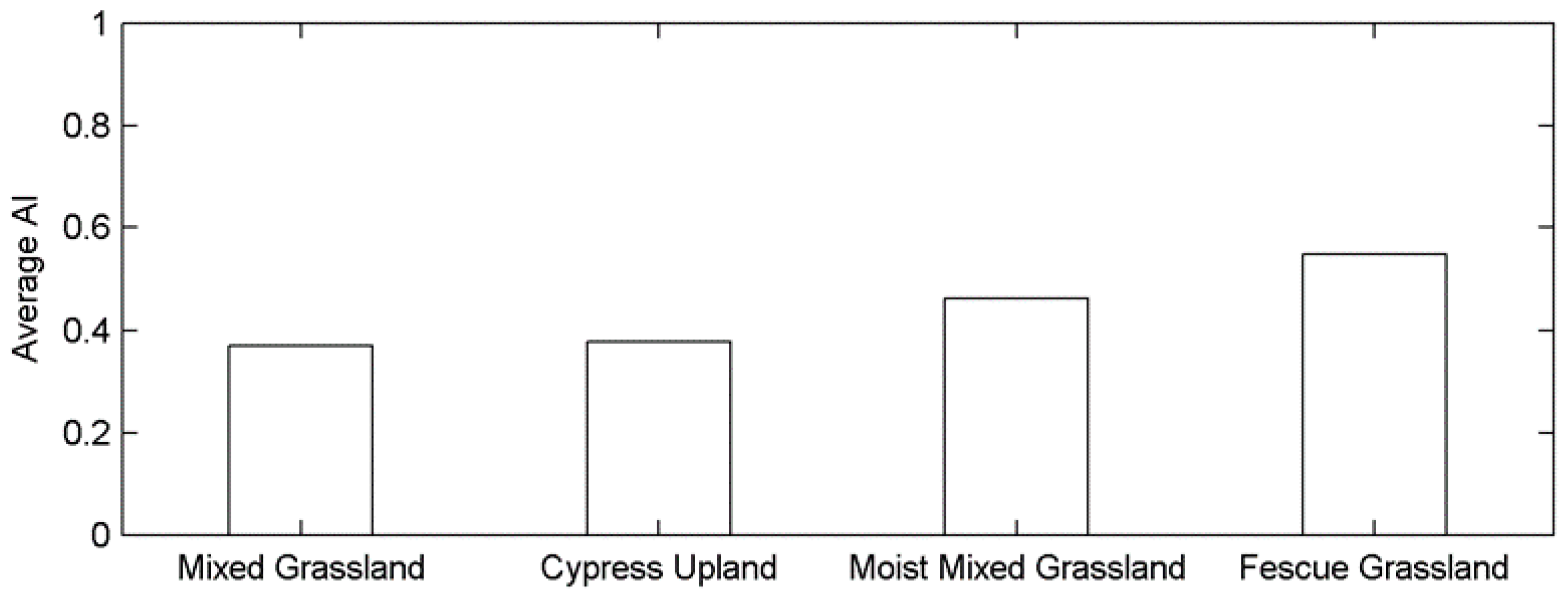
2.1. Canadian Prairie Grasslands
2.2. Grassland Phenology Extraction
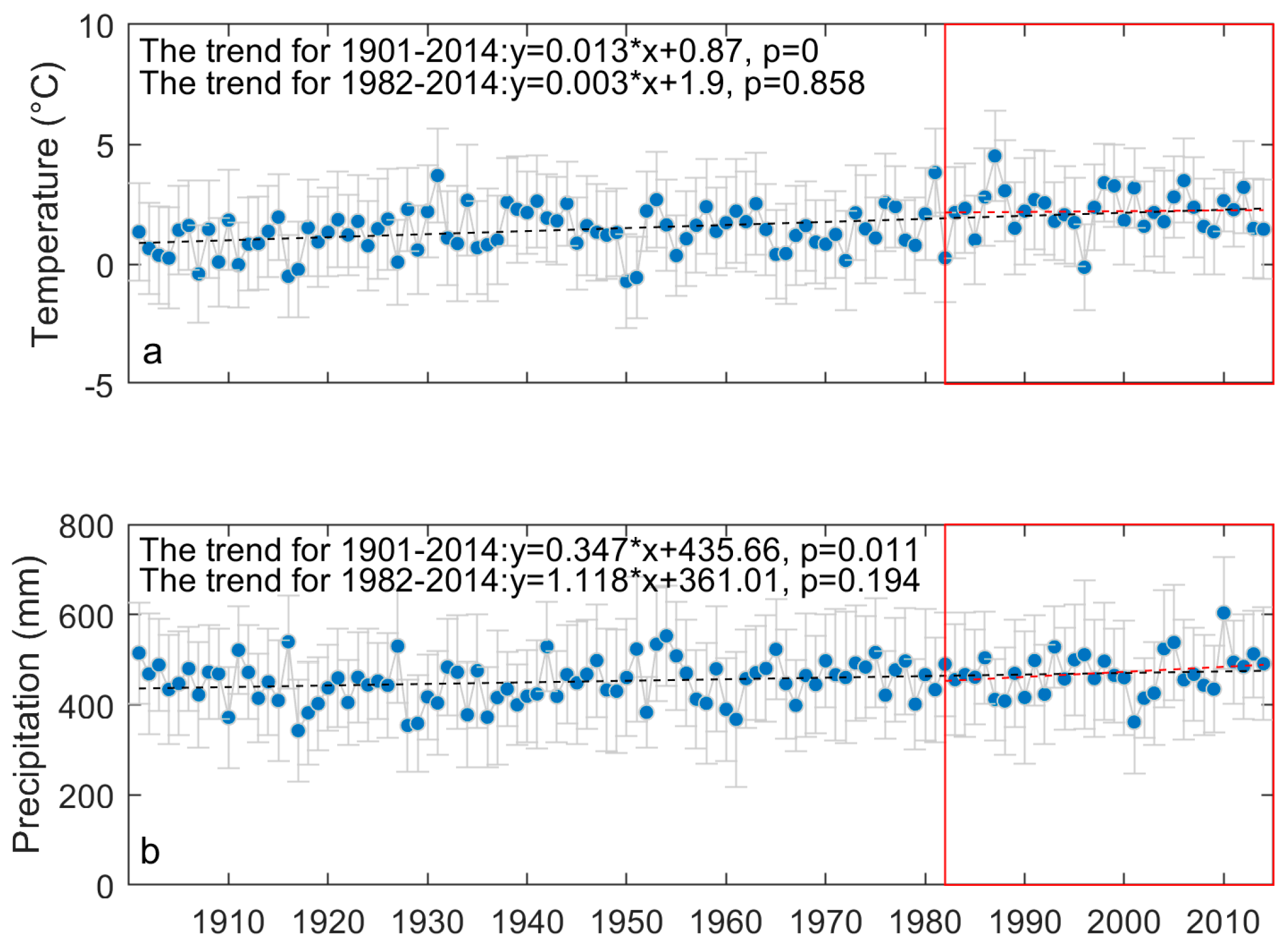
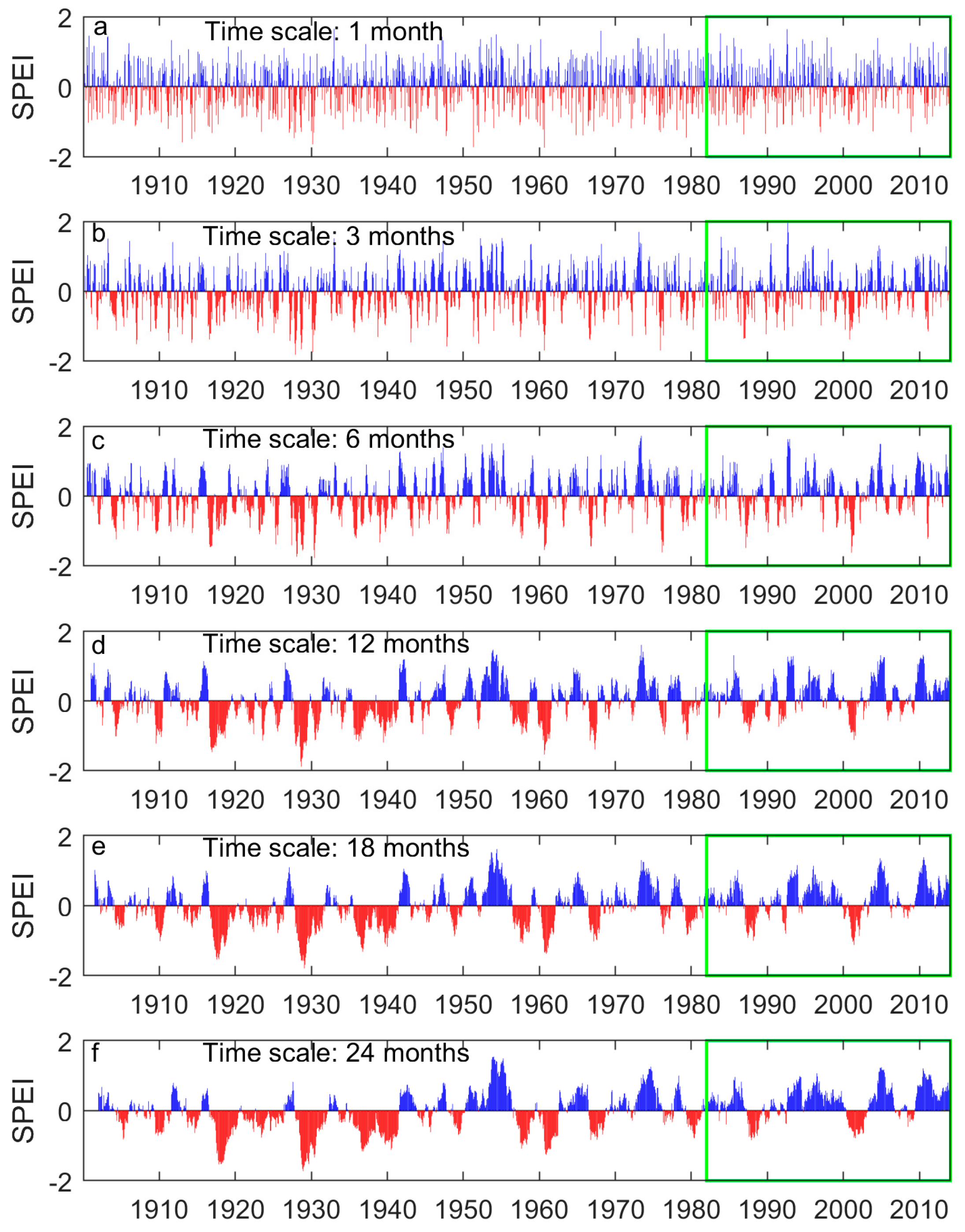
2.3. Drought and Precipitation for Grasslands in the Prairie Ecozone
2.4. Statistics
3. Results and Discussion
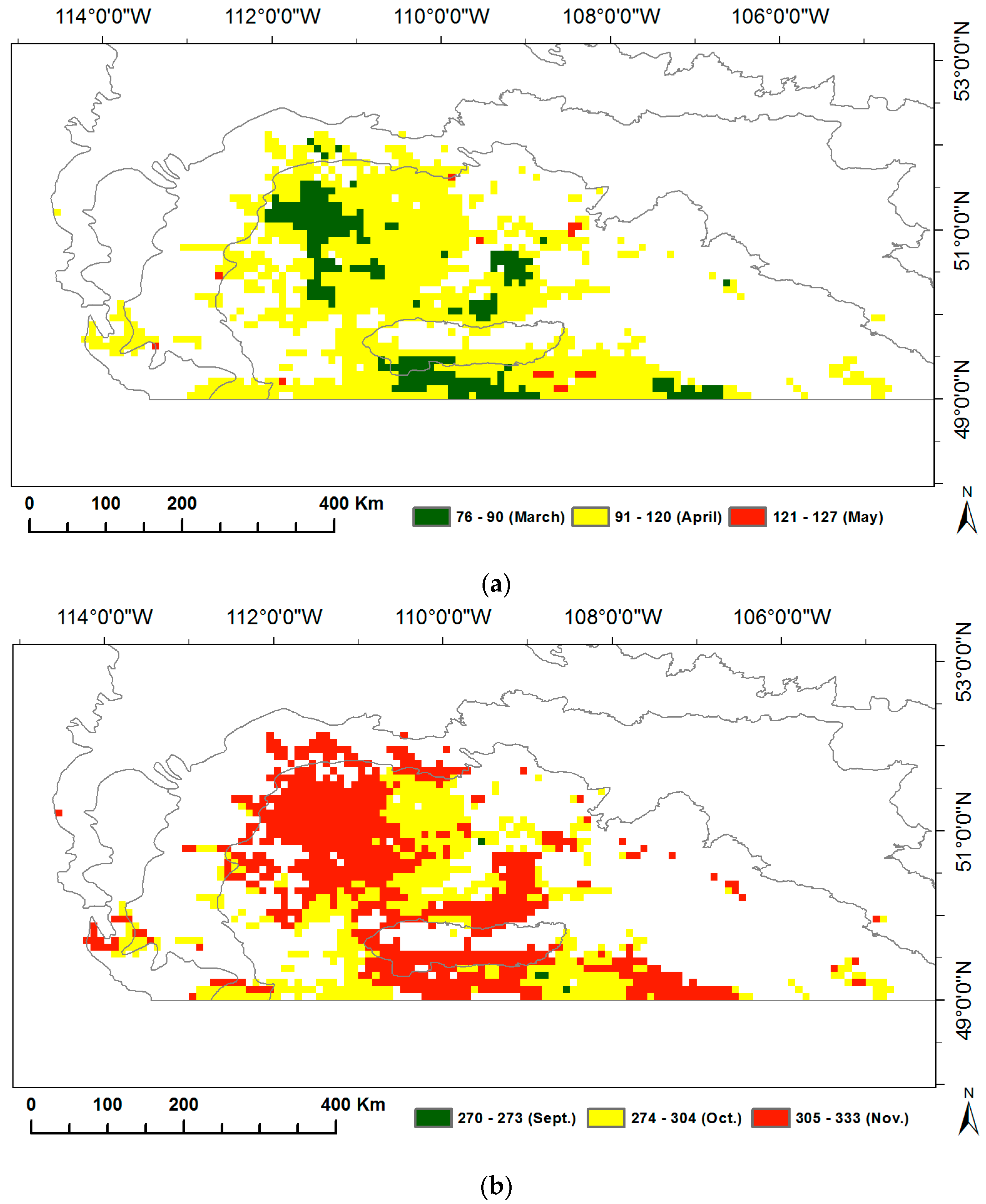
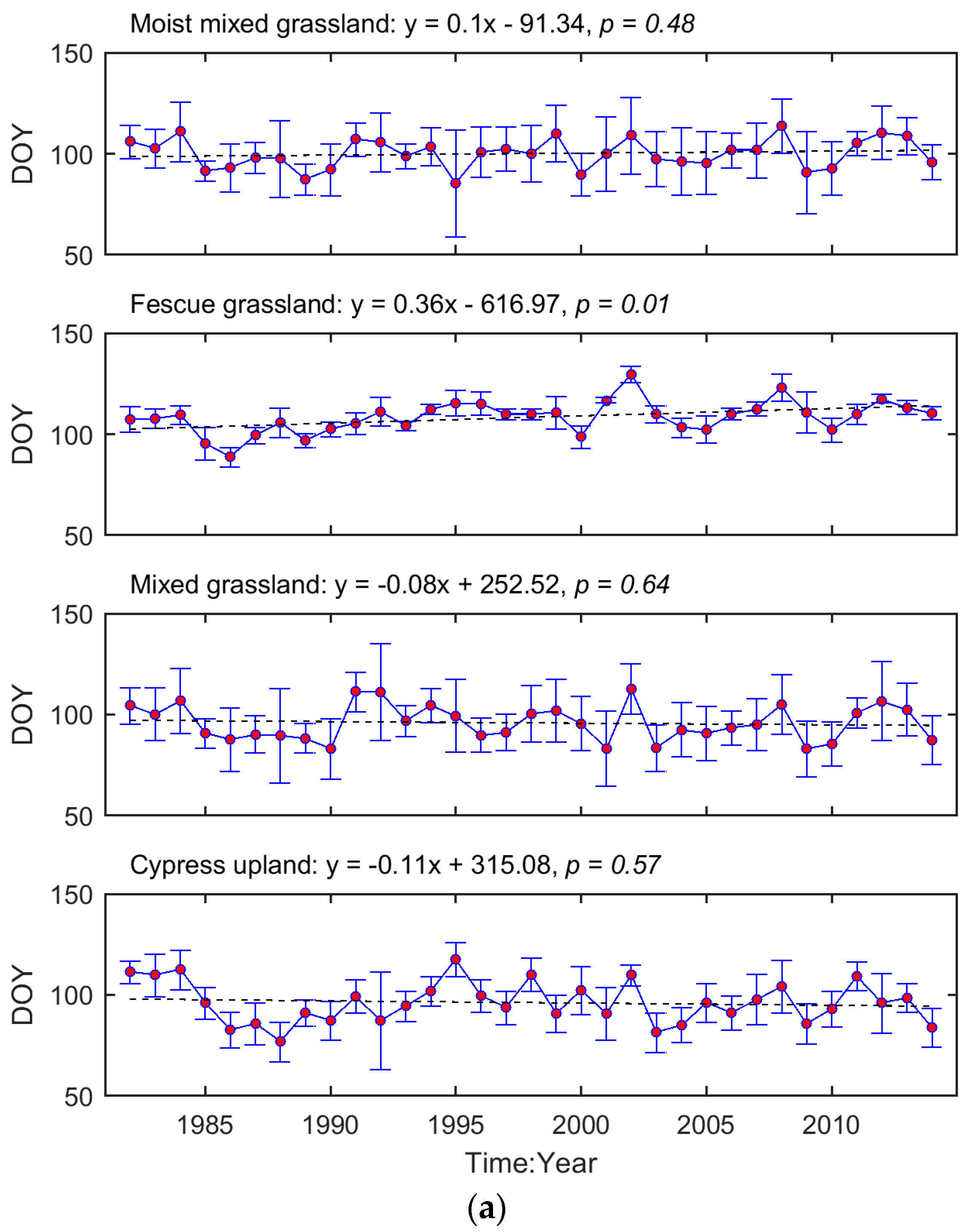
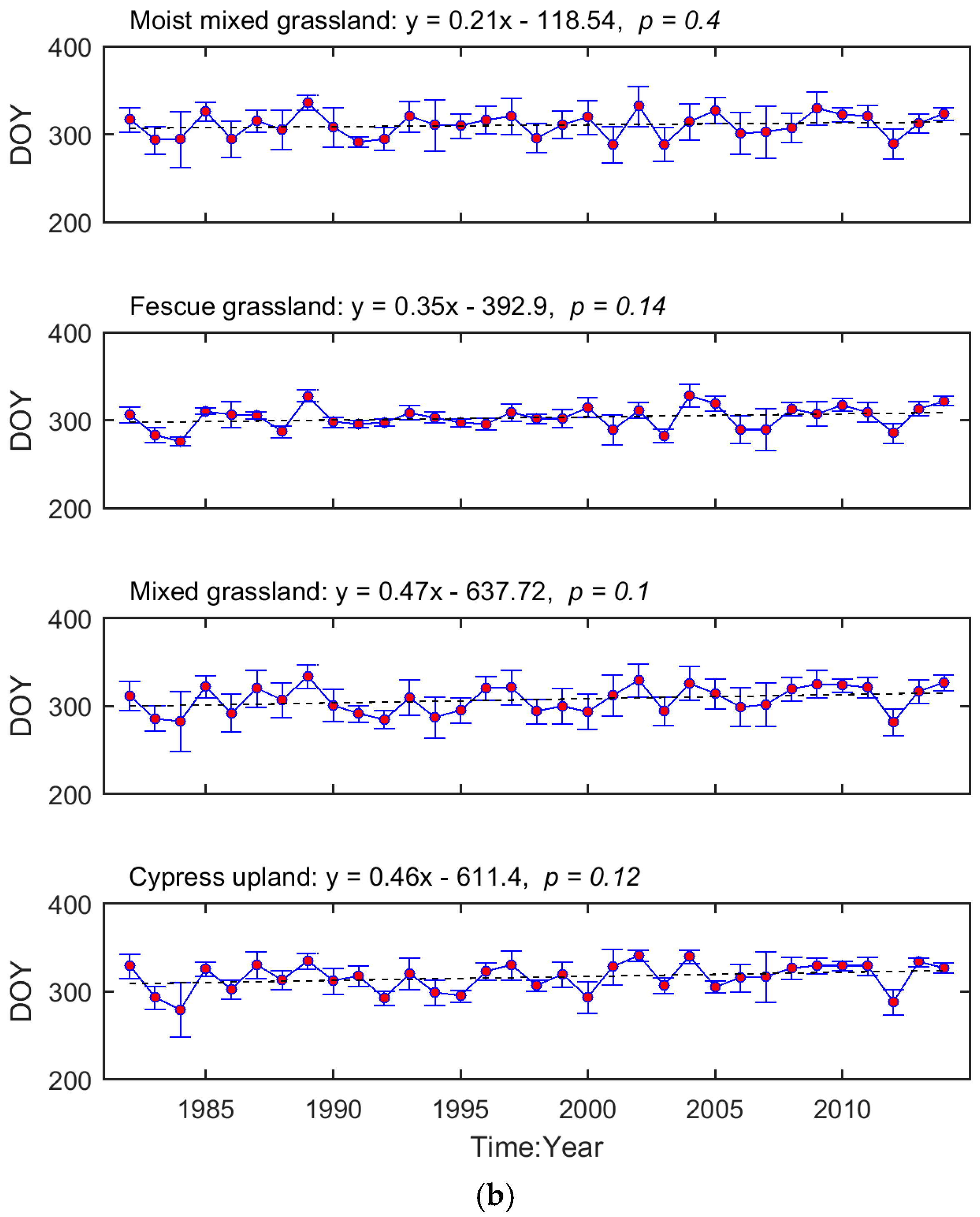
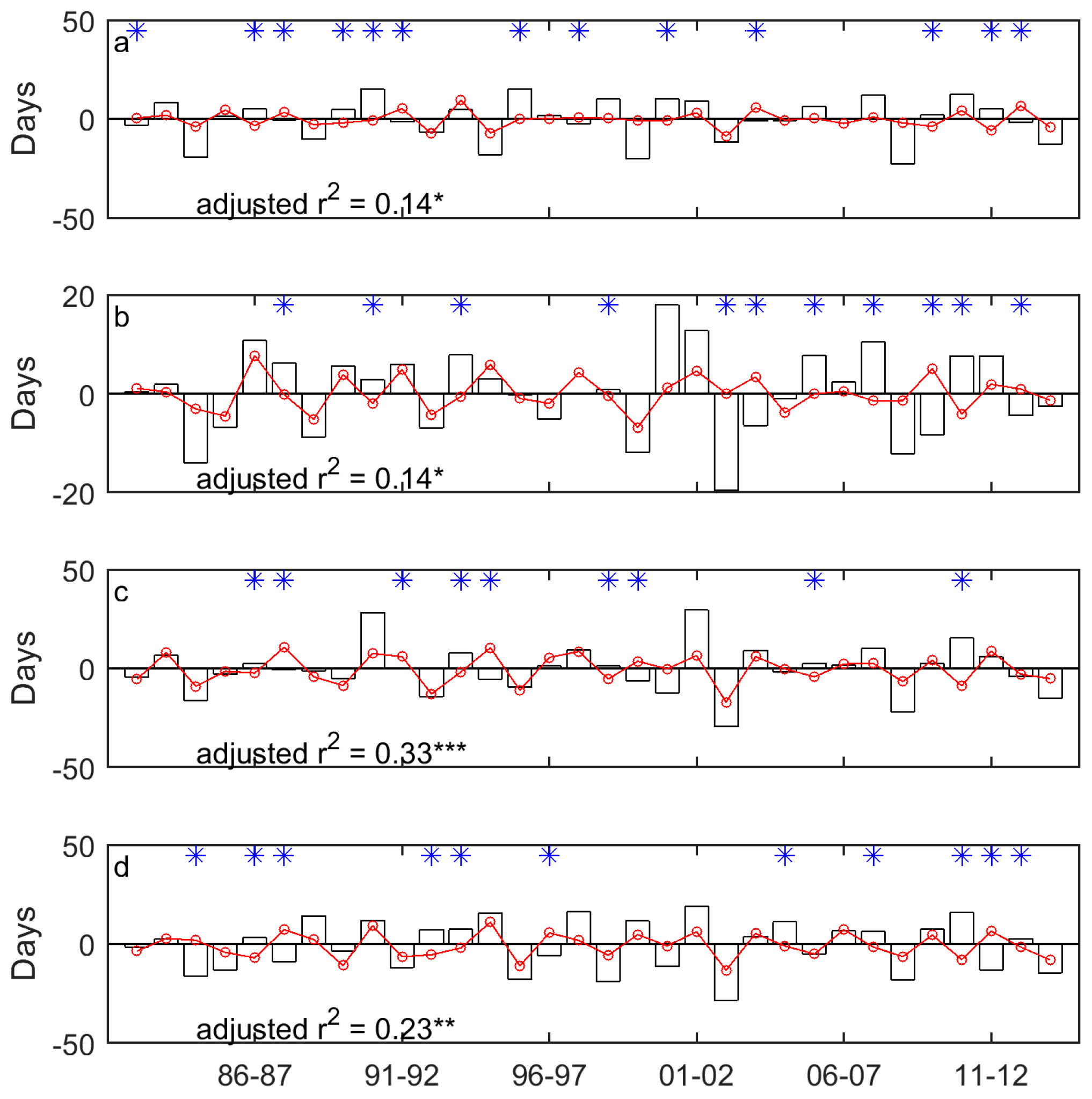
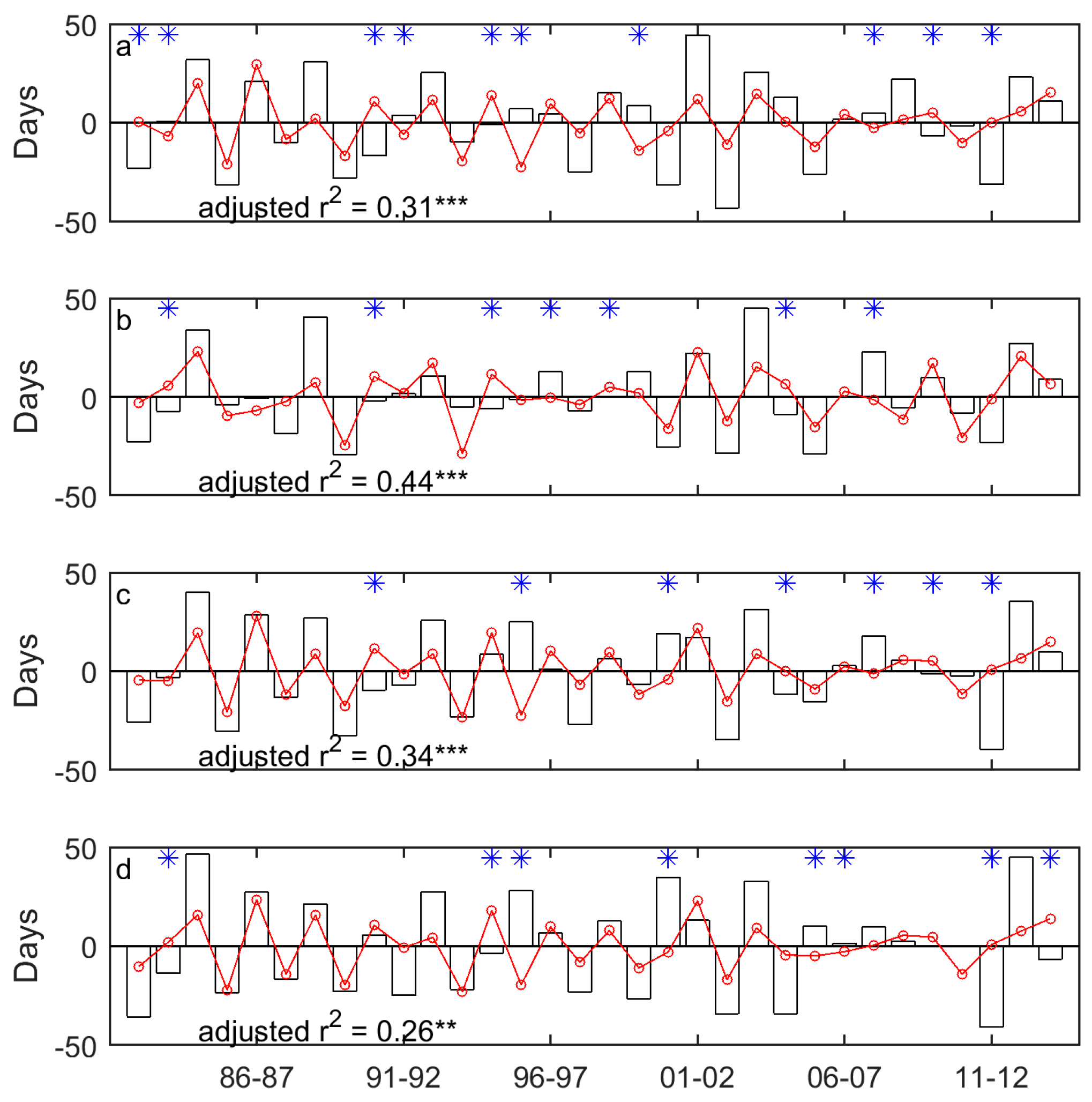
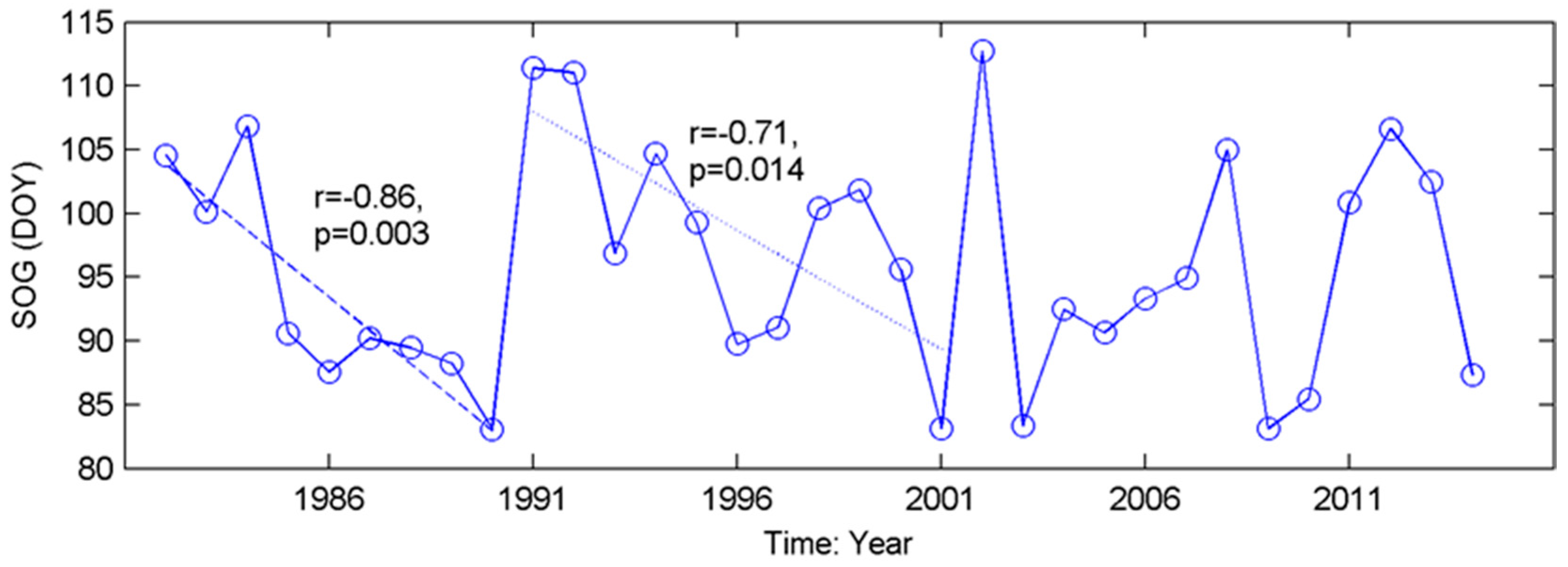
3.1. Long-Term Means and Interannual Changes of Grassland Phenology in the Prairie Ecozone from 1982–2014
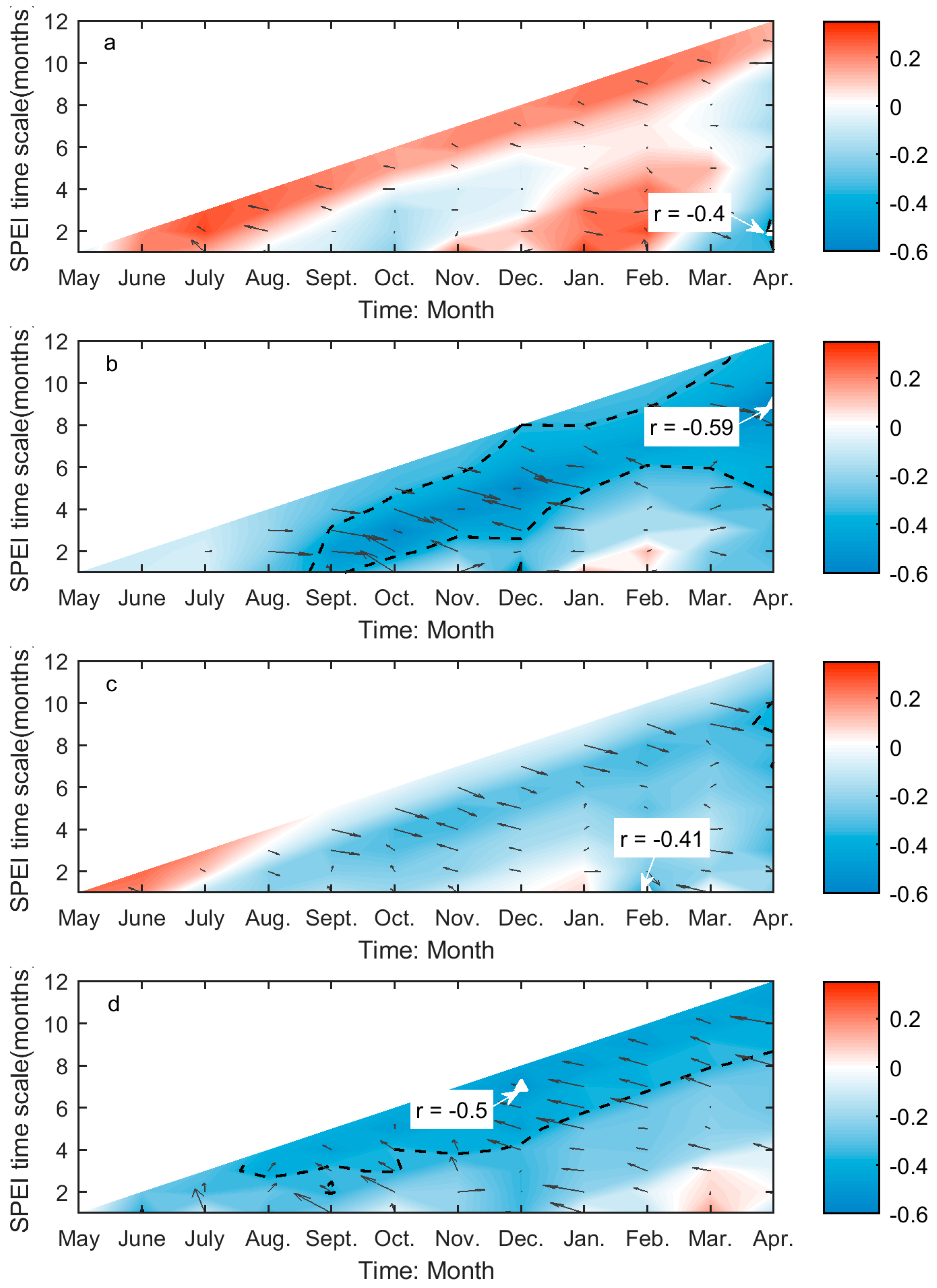
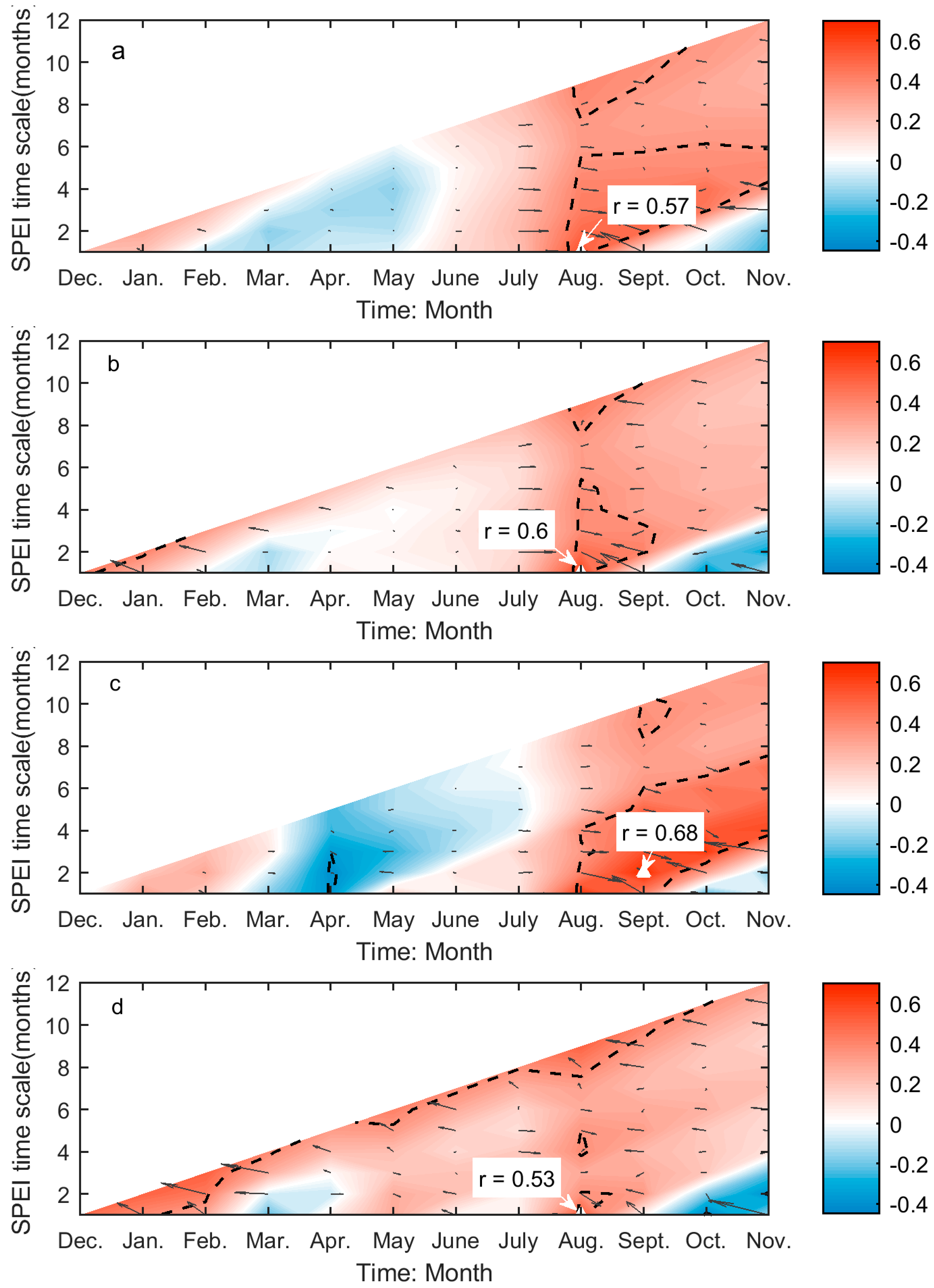
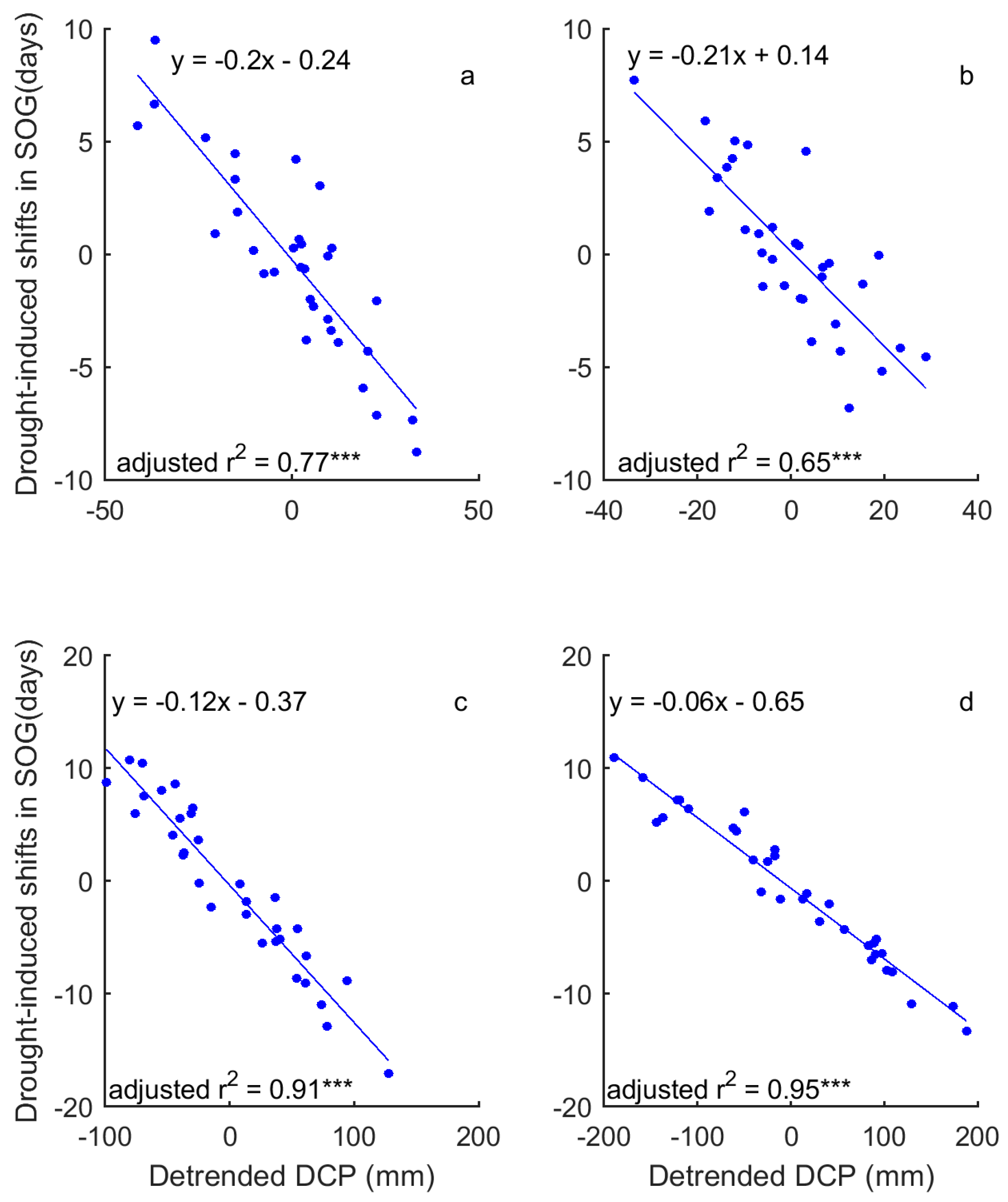
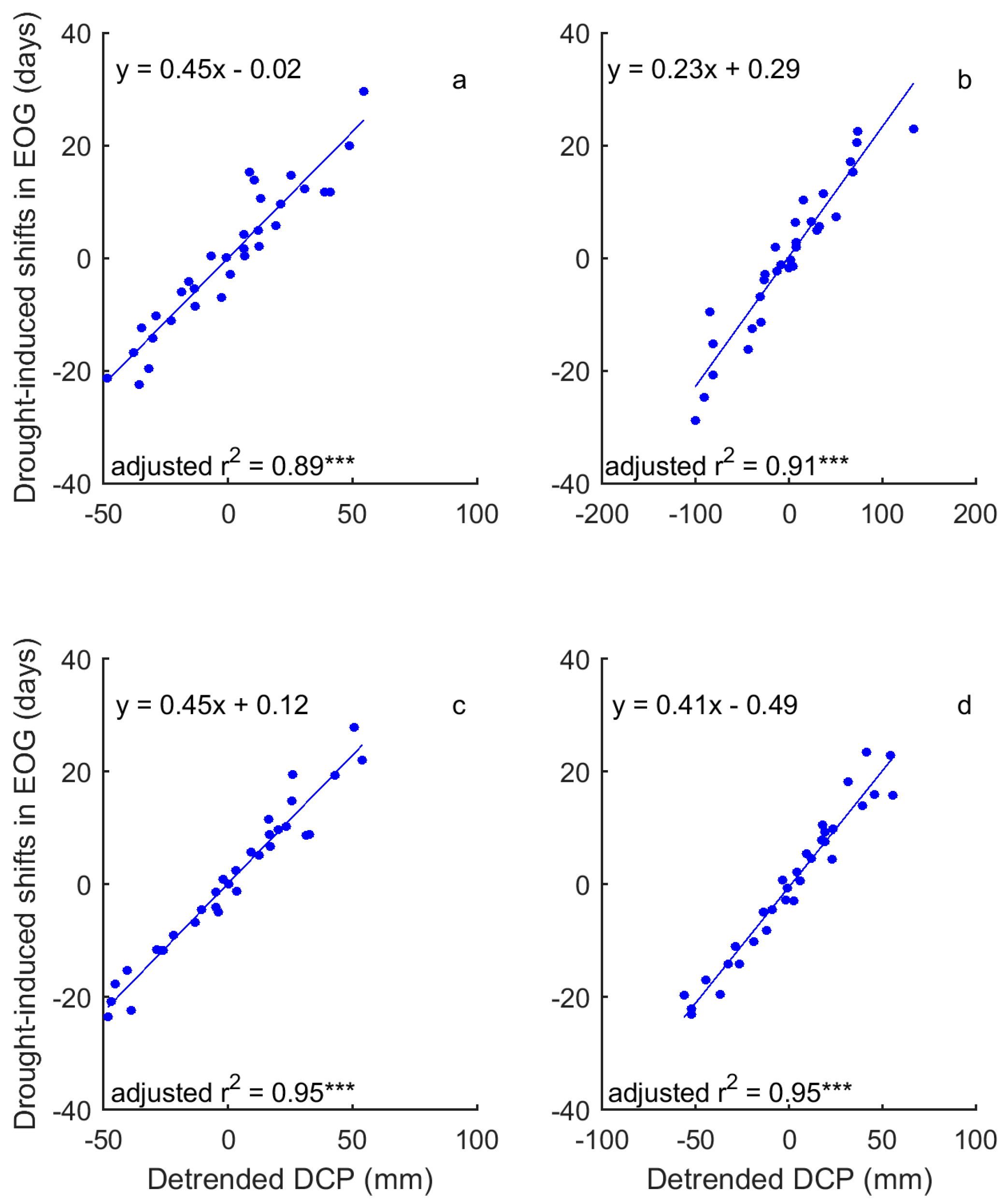
3.2. Correlations between Grassland Phenology and Preceding Drought Events
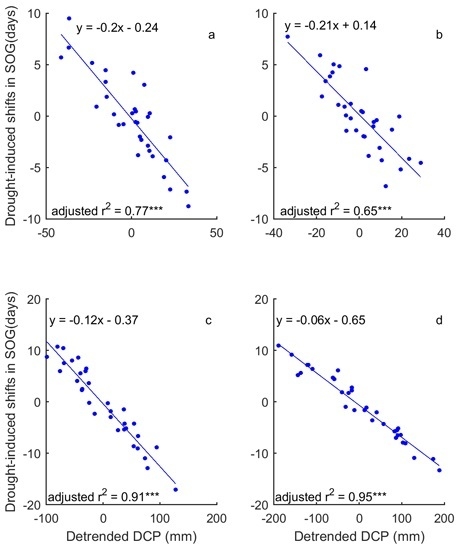
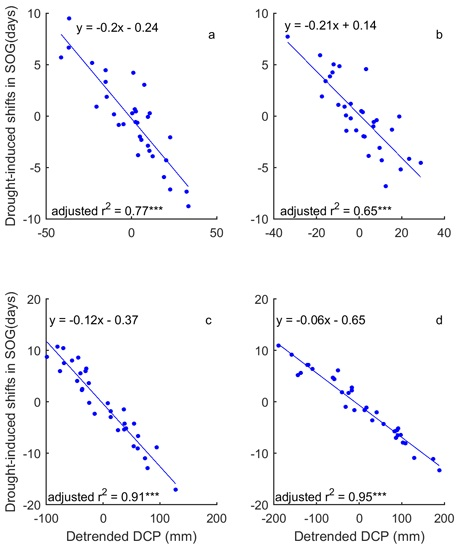
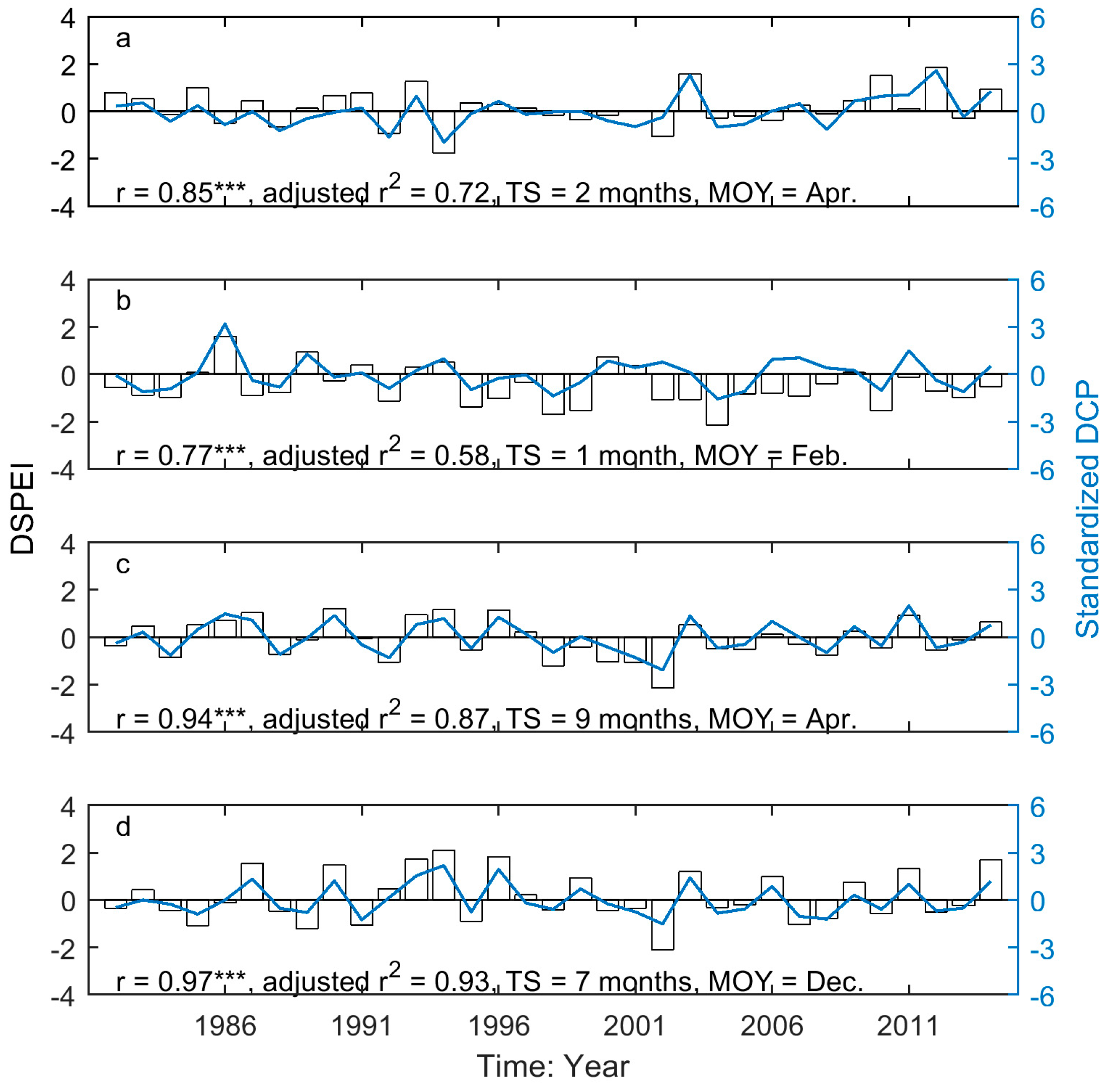
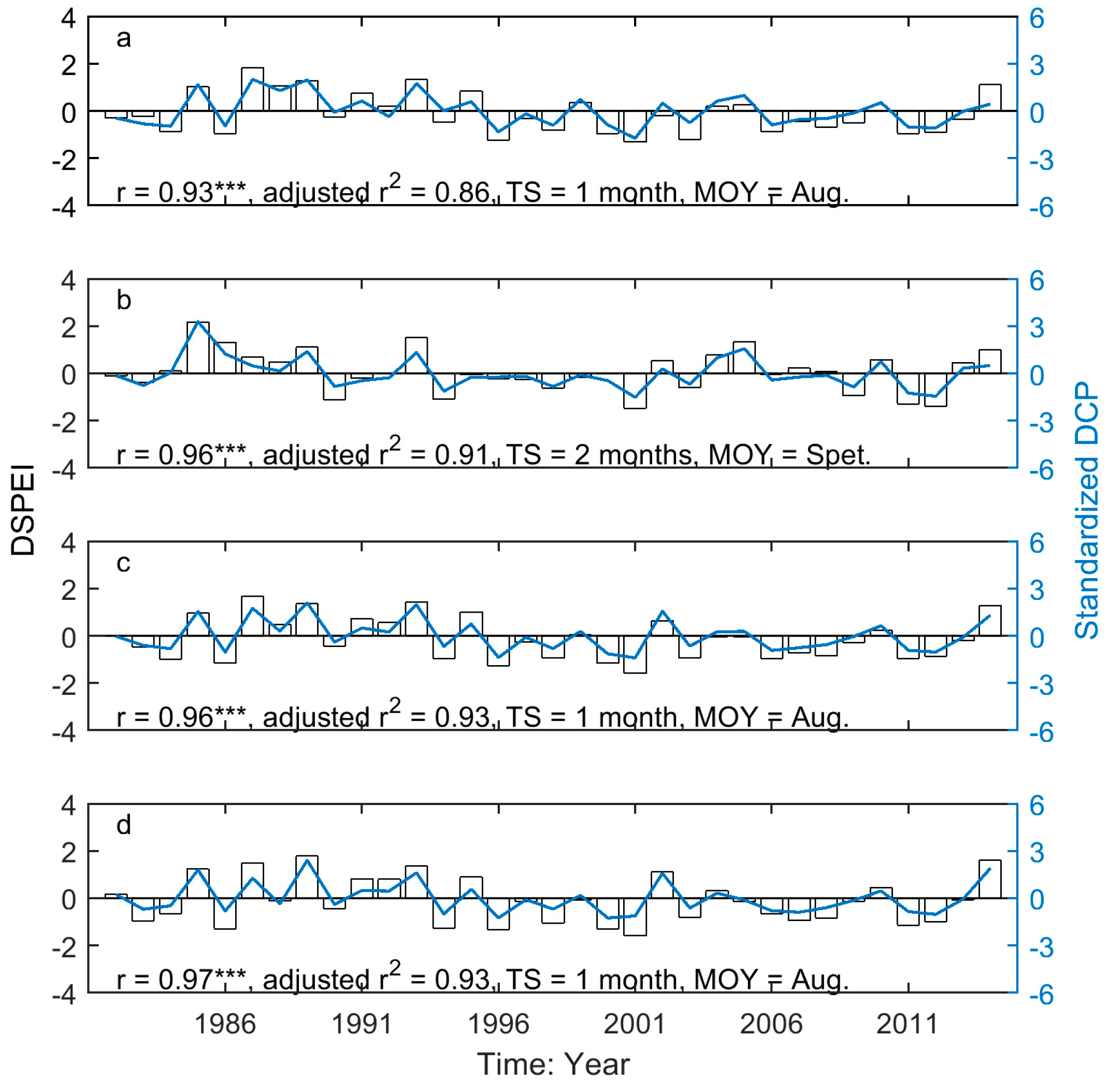
3.3. Drought-Induced Shifts in Grassland Relation to Phenology and Their Precipitation
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A




References
- Anderson, R.C. Evolution and origin of the Central Grassland of North America: Climate, fire, and mammalian grazers. J. Torrey Bot. Soc. 2006, 133, 626–647. [Google Scholar] [CrossRef]
- White, R.P.; Murray, S.; Rohweder, M.; Prince, S.; Thompson, K. Grassland Ecosystems; World Resources Institute: Washington, DC, USA, 2000. [Google Scholar]
- Parton, W.; Morgan, J.; Smith, D.; Del Grosso, S.; Prihodko, L.; Lecain, D.; Kelly, R.; Lutz, S. Impact of precipitation dynamics on net ecosystem productivity. Glob. Chang. Biol. 2012, 18, 915–927. [Google Scholar] [CrossRef]
- Scurlock, J.M.O.; Hall, D.O. The global carbon sink: A grassland perspective. Glob. Chang. Biol. 1998, 4, 229–233. [Google Scholar] [CrossRef]
- Piao, S.L.; Tan, K.; Nan, H.J.; Ciais, P.; Fang, J.Y.; Wang, T.; Vuichard, N.; Zhu, B.A. Impacts of climate and CO2 changes on the vegetation growth and carbon balance of Qinghai-Tibetan grasslands over the past five decades. Glob. Planet. Chang. 2012, 98–99, 73–80. [Google Scholar] [CrossRef]
- Puissant, J.; Mills, R.T.E.; Robroek, B.J.M.; Gavazov, K.; Perrette, Y.; De Danieli, S.; Spiegelberger, T.; Buttler, A.; Brun, J.J.; Cecillon, L. Climate change effects on the stability and chemistry of soil organic carbon pools in a subalpine grassland. Biogeochemistry 2017, 132, 123–139. [Google Scholar] [CrossRef]
- Chang, J.F.; Ciais, P.; Viovy, N.; Vuichard, N.; Herrero, M.; Havlik, P.; Wang, X.H.; Sultan, B.; Soussana, J.F. Effect of climate change, CO2 trends, nitrogen addition, and land-cover and management intensity changes on the carbon balance of European grasslands. Glob. Chang. Biol. 2016, 22, 338–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogee, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Bonsal, B.; Regier, M. Historical comparison of the 2001/2002 drought in the Canadian Prairies. Clim. Res. 2007, 33, 229–242. [Google Scholar] [CrossRef]
- Dai, A.G. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Masud, M.B.; Khaliq, M.N.; Wheater, H.S. Future changes to drought characteristics over the Canadian Prairie Provinces based on NARCCAP multi-RCM ensemble. Clim. Dyn. 2017, 48, 2685–2705. [Google Scholar] [CrossRef]
- Bloor, J.M.G.; Pichon, P.; Falcimagne, R.; Leadley, P.; Soussana, J.F. Effects of Warming, Summer Drought, and CO2 Enrichment on Aboveground Biomass Production, Flowering Phenology, and Community Structure in an Upland Grassland Ecosystem. Ecosystems 2010, 13, 888–900. [Google Scholar] [CrossRef]
- Frank, D.A. Drought effects on above- and belowground production of a grazed temperate grassland ecosystem. Oecologia 2007, 152, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Boot, C.M.; Schaeffer, S.M.; Schimel, J.P. Static osmolyte concentrations in microbial biomass during seasonal drought in a California grassland. Soil Biol. Biochem. 2013, 57, 356–361. [Google Scholar] [CrossRef]
- Manea, A.; Sloane, D.R.; Leishman, M.R. Reductions in native grass biomass associated with drought facilitates the invasion of an exotic grass into a model grassland system. Oecologia 2016, 181, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Craine, J.M.; Ocheltree, T.W.; Nippert, J.B.; Towne, E.G.; Skibbe, A.M.; Kembel, S.W.; Fargione, J.E. Global diversity of drought tolerance and grassland climate-change resilience. Nat. Clim. Chang. 2013, 3, 63–67. [Google Scholar] [CrossRef]
- White, C.S.; Moore, D.I.; Craig, J.A. Regional-scale drought increases potential soil fertility in semiarid grasslands. Biol. Fertil. Soils 2004, 40, 73–78. [Google Scholar] [CrossRef]
- De Vries, F.T.; Brown, C.; Stevens, C.J. Grassland species root response to drought: Consequences for soil carbon and nitrogen availability. Plant Soil 2016, 409, 297–312. [Google Scholar] [CrossRef]
- Sanaullah, M.; Chabbi, A.; Girardin, C.; Durand, J.L.; Poirier, M.; Rumpel, C. Effects of drought and elevated temperature on biochemical composition of forage plants and their impact on carbon storage in grassland soil. Plant Soil 2014, 374, 767–778. [Google Scholar] [CrossRef]
- Polyakov, V.O.; Nearing, M.A.; Stone, J.J.; Hamerlynck, E.P.; Nichols, M.H.; Collins, C.D.H.; Scott, R.L. Runoff and erosional responses to a drought-induced shift in a desert grassland community composition. J. Geophys. Res. Biogeosci. 2010, 115. [Google Scholar] [CrossRef]
- Stampfli, A.; Zeiter, M. Plant regeneration directs changes in grassland composition after extreme drought: A 13-year study in southern Switzerland. J. Ecol. 2004, 92, 568–576. [Google Scholar] [CrossRef]
- Zeiter, M.; Scharrer, S.; Zweifel, R.; Newbery, D.M.; Stampfli, A. Timing of extreme drought modifies reproductive output in semi-natural grassland. J. Veg. Sci. 2016, 27, 238–248. [Google Scholar] [CrossRef]
- Villarreal, M.L.; Norman, L.M.; Buckley, S.; Wallace, C.S.A.; Coe, M.A. Multi-index time series monitoring of drought and fire effects on desert grasslands. Remote Sens. Environ. 2016, 183, 186–197. [Google Scholar] [CrossRef]
- Thorpe, J. Vulnerability of Prairie Grasslands to Climate Change; Saskatchewan Research Council: Saskatoon, SK, Canada, 2011. [Google Scholar]
- White, M.A.; Thornton, P.E.; Running, S.W. A continental phenology model for monitoring vegetation responses to interannual climatic variability. Glob. Biogeochem. Cycles 1997, 11, 217–234. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Myneni, R.B.; Keeling, C.D.; Tucker, C.J.; Asrar, G.; Nemani, R.R. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature 1997, 386, 698–702. [Google Scholar] [CrossRef]
- Richardson, A.D.; Black, T.A.; Ciais, P.; Delbart, N.; Friedl, M.A.; Gobron, N.; Hollinger, D.Y.; Kutsch, W.L.; Longdoz, B.; Luyssaert, S.; et al. Influence of spring and autumn phenological transitions on forest ecosystem productivity. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3227–3246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.Y.; Hou, X.H.; Peng, D.L.; Gonsamo, A.; Xu, S.G. Land surface phenology of China’s temperate ecosystems over 1999–2013: Spatial-temporal patterns, interaction effects, covariation with climate and implications for productivity. Agric. For. Meteorol. 2016, 216, 177–187. [Google Scholar] [CrossRef]
- Mamolos, A.P.; Veresoglou, D.S.; Noitsakis, V.; Gerakis, A. Differential drought tolerance of five coexisting plant species in Mediterranean lowland grasslands. J. Arid Environ. 2001, 49, 329–341. [Google Scholar] [CrossRef]
- Wolf, S.; Eugster, W.; Ammann, C.; Hani, M.; Zielis, S.; Hiller, R.; Stieger, J.; Imer, D.; Merbold, L.; Buchmann, N. Contrasting response of grassland versus forest carbon and water fluxes to spring drought in Switzerland. Environ. Res. Lett. 2013, 8. [Google Scholar] [CrossRef]
- Glade, F.E.; Miranda, M.D.; Meza, F.J.; van Leeuwen, W.J.D. Productivity and phenological responses of natural vegetation to present and future inter-annual climate variability across semi-arid river basins in chile. Environ. Monit. Assess. 2016, 188, 676. [Google Scholar] [CrossRef] [PubMed]
- Tao, F.; Yokozawa, M.; Zhang, Z.; Hayashi, Y.; Ishigooka, Y. Land surface phenology dynamics and climate variations in the North East China Transect (NECT), 1982–2000. Int. J. Remote Sens. 2008, 29, 5461–5478. [Google Scholar] [CrossRef]
- Riley, J.L.; Brodribb, K.E.; Green, S.E. A Conservation Blueprint for Canada’s Prairies and Parklands; Nature Conservancy of Canada: Toronto, ON, Canada, 2007. [Google Scholar]
- Shorthouse, J.D. Ecoregions of Canada’s prairie grasslands. Arthropods Can. Grassl. 2010, 1, 53–81. [Google Scholar]
- Chipanshi, A.C.; Findlater, K.M.; Hadwen, T.; O’Brien, E.G. Analysis of consecutive droughts on the Canadian Prairies. Clim. Res. 2006, 30, 175–187. [Google Scholar] [CrossRef]
- McGinn, S.M. Weather and climate patterns in Canada’s prairie grasslands. Arthropods Can. Grassl. 2010, 1, 105–119. [Google Scholar]
- Zhang, X.; Brown, R.; Vincent, L.; Skinner, W.; Feng, Y.; Mekis, E. Canadian Climate Trends, 1950–2007; Canadian Biodiversity: Ecosystem Status and Trends; Technical Thematic Report No. 5; The Canadian Councils of Resource Ministers: Ottawa, ON, Canada, 2010. [Google Scholar]
- Thorpe, J.; Wolfe, S.A.; Houston, B. Potential impacts of climate change on grazing capacity of native grasslands in the Canadian prairies. Can. J. Soil Sci. 2008, 88, 595–609. [Google Scholar] [CrossRef]
- Kochy, M.; Wilson, S.D. Semiarid grassland responses to short-term variation in water availability. Plant Ecol. 2004, 174, 197–203. [Google Scholar] [CrossRef]
- Haddad, N.M.; Tilman, D.; Knops, J.M.H. Long-term oscillations in grassland productivity induced by drought. Ecol. Lett. 2002, 5, 110–120. [Google Scholar] [CrossRef]
- Adams, B.W.; Association, A.C.; Development, A.A.S.R. Beneficial Grazing Management Practices for Sage Grouse (centrocercus urophasianus) and Ecology of Silver Sagebrush (artemisia cana prush subsp. Cana) in Southeastern Alberta; Alberta Sustainable Resource Development: Edmonton, AB, Canada, 2004. [Google Scholar]
- Beaubien, E.G.; Hamann, A. Plant phenology networks of citizen scientists: Recommendations from two decades of experience in Canada. Int. J. Biometeorol. 2011, 55, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Reed, B.C. Trend analysis of time-series phenology of North America derived from satellite data. Gisci. Remote Sens. 2006, 43, 24–38. [Google Scholar] [CrossRef]
- Li, Z.; Guo, X. Detecting Climate Effects on Vegetation in Northern Mixed Prairie Using NOAA AVHRR 1-km Time-Series NDVI Data. Remote Sens. 2012, 4, 120–134. [Google Scholar] [CrossRef]
- Channan, S.; Collins, K.; Emanuel, W. Global Mosaics of the Standard MODIS Land Cover Type Data; University of Maryland and the Pacific Northwest Laboratory: College Park, MA, USA, 2014.
- Ecological Stratification Working Group (Canada); Center for Land and Biological Resources Research (Canada); Canada State of the Environment Directorate. A National Ecological Framework for Canada; Centre for Land and Biological Resources Research; State of the Environment Directorate: Hull, QC, Canada, 1996. [Google Scholar]
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R.; Vermote, E.F.; El Saleous, N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and spot vegetation NDVI data. Int. J. Remote Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- Zhu, Z.C.; Bi, J.; Pan, Y.Z.; Ganguly, S.; Anav, A.; Xu, L.; Samanta, A.; Piao, S.L.; Nemani, R.R.; Myneni, R.B. Global Data Sets of Vegetation Leaf Area Index (LAI)3g and Fraction of Photosynthetically Active Radiation (FPAR)3g Derived from Global Inventory Modeling and Mapping Studies (GIMMS) Normalized Difference Vegetation Index (NDVI3g) for the Period 1981 to 2011. Remote Sens. 2013, 5, 927–948. [Google Scholar]
- Zeng, F.W.; Collatz, G.J.; Pinzon, J.E.; Ivanoff, A. Evaluating and Quantifying the Climate-Driven Interannual Variability in Global Inventory Modeling and Mapping Studies (GIMMS) Normalized Difference Vegetation Index (NDVI3g) at Global Scales. Remote Sens. 2013, 5, 3918–3950. [Google Scholar] [CrossRef]
- Eastman, J.R.; Sangermano, F.; Machado, E.A.; Rogan, J.; Anyamba, A. Global Trends in Seasonality of Normalized Difference Vegetation Index (NDVI), 1982–2011. Remote Sens. 2013, 5, 4799–4818. [Google Scholar] [CrossRef]
- Atzberger, C.; Klisch, A.; Mattiuzzi, M.; Vuolo, F. Phenological metrics derived over the European continent from NDVI3g data and MODIS time series. Remote Sens. 2014, 6, 257–284. [Google Scholar] [CrossRef]
- Ibrahim, Y.Z.; Balzter, H.; Kaduk, J.; Tucker, C.J. Land Degradation Assessment Using Residual Trend Analysis of GIMMS NDVI3g, Soil Moisture and Rainfall in Sub-Saharan West Africa from 1982 to 2012. Remote Sens. 2015, 7, 5471–5494. [Google Scholar] [CrossRef]
- Marshall, M.; Okuto, E.; Kang, Y.; Opiyo, E.; Ahmed, M. Global assessment of Vegetation Index and Phenology Lab (VIP) and Global Inventory Modeling and Mapping Studies (GIMMS) version 3 products. Biogeosciences 2016, 13, 625–639. [Google Scholar] [CrossRef]
- Jonsson, P.; Eklundh, L. TIMESAT—A program for analyzing time-series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef]
- Julien, Y.; Sobrino, J.A. Global land surface phenology trends from GIMMS database. Int. J. Remote Sens. 2009, 30, 3495–3513. [Google Scholar] [CrossRef]
- Fisher, J.I.; Mustard, J.F. Cross-scalar satellite phenology from ground, Landsat, and MODIS data. Remote Sens. Environ. 2007, 109, 261–273. [Google Scholar] [CrossRef]
- White, M.A.; de Beurs, K.M.; Didan, K.; Inouye, D.W.; Richardson, A.D.; Jensen, O.P.; O’Keefe, J.; Zhang, G.; Nemani, R.R.; van Leeuwen, W.J.D.; et al. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982–2006. Glob. Chang. Biol. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- Cong, N.; Piao, S.; Chen, A.; Wang, X.; Lin, X.; Chen, S.; Han, S.; Zhou, G.; Zhang, X. Spring vegetation green-up date in China inferred from SPOT NDVI data: A multiple model analysis. Agric. For. Meteorol. 2012, 165, 104–113. [Google Scholar] [CrossRef]
- Zhou, D.C.; Zhao, S.Q.; Zhang, L.X.; Liu, S.G. Remotely sensed assessment of urbanization effects on vegetation phenology in China’s 32 major cities. Remote Sens. Environ. 2016, 176, 272–281. [Google Scholar] [CrossRef]
- Wang, C.; Tang, Y.H.; Chen, J. Plant phenological synchrony increases under rapid within-spring warming. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Serrano, S.M.; Begueria, S.; Lopez-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Begueria, S.; Vicente-Serrano, S.M.; Reig, F.; Latorre, B. Standardized precipitation evapotranspiration index (SPEI) revisited: Parameter fitting, evapotranspiration models, tools, datasets and drought monitoring. Int. J. Climatolog. 2014, 34, 3001–3023. [Google Scholar] [CrossRef]
- Palmer, W.C. Meteorological Drought; US Department of Commerce, Weather Bureau: Washington, DC, USA, 1965; Volume 30.
- Wells, N.; Goddard, S.; Hayes, M.J. A self-calibrating Palmer Drought Severity Index. J. Clim. 2004, 17, 2335–2351. [Google Scholar] [CrossRef]
- McKee, T.B.; Doesken, N.J.; Kleist, J. The relationship of drought frequency and duration to time scales. In Proceedings of the 8th Conference on Applied Climatology, Anaheim, CA, USA, 17–22 January 1993; American Meteorological Society: Boston, MA, USA; pp. 179–183. [Google Scholar]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations - the CRU TS3.10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Begueria, S.; Trigo, R.; Lopez-Moreno, J.I.; Azorin-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J.; et al. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, J.T.; Inouye, D.W.; McKinney, A.M.; Colautti, R.I.; Mitchell-Olds, T. Phenotypic plasticity and adaptive evolution contribute to advancing flowering phenology in response to climate change. Proc. R. Soc. B Biol. Sci. 2012, 279, 3843–3852. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.M.; Tucker, C.J.; Kaufmann, R.K.; Slayback, D.; Shabanov, N.V.; Myneni, R.B. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J. Geophys. Res. Atmos. 2001, 106, 20069–20083. [Google Scholar] [CrossRef]
- Buermann, W.; Bikash, P.R.; Jung, M.; Burn, D.H.; Reichstein, M. Earlier springs decrease peak summer productivity in North American boreal forests. Environ. Res. Lett. 2013, 8. [Google Scholar] [CrossRef]
- Garonna, I.; de Jong, R.; Schaepman, M.E. Variability and evolution of global land surface phenology over the past three decades (1982–2012). Glob. Chang. Biol. 2016, 22, 1456–1468. [Google Scholar] [CrossRef] [PubMed]
- Lesica, P.; Kittelson, P. Precipitation and temperature are associated with advanced flowering phenology in a semi-arid grassland. J. Arid Environ. 2010, 74, 1013–1017. [Google Scholar] [CrossRef]
- Zhu, L.K.; Meng, J.J. Determining the relative importance of climatic drivers on spring phenology in grassland ecosystems of semi-arid areas. Int. J. Biometeorol. 2015, 59, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Ganjurjav, H.; Gao, Q.Z.; Schwartz, M.W.; Zhu, W.Q.; Liang, Y.; Li, Y.; Wan, Y.F.; Cao, X.J.; Williamson, M.A.; Jiangcun, W.; et al. Complex responses of spring vegetation growth to climate in a moisture-limited alpine meadow. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.Y.; Xu, J.C.; Okuto, E.; Luedeling, E. Seasonal Response of Grasslands to Climate Change on the Tibetan Plateau. PLoS ONE 2012, 7, e49230. [Google Scholar] [CrossRef] [PubMed]
- Nandintsetseg, B.; Shinoda, M.; Kimura, R.; Ibaraki, Y. Relationship between Soil Moisture and Vegetation Activity in the Mongolian Steppe. Sola 2010, 6, 29–32. [Google Scholar] [CrossRef]
- Martiny, N.; Philippon, N.; Richard, Y.; Camberlin, P.; Reason, C. Predictability of NDVI in semi-arid African regions. Theor. Appl. Climatol. 2010, 100, 467–484. [Google Scholar] [CrossRef]
- Morgan, J.; Pataki, D.; Körner, C.; Clark, H.; Del Grosso, S.; Grünzweig, J.; Knapp, A.; Mosier, A.; Newton, P.; Niklaus, P.A. Water relations in grassland and desert ecosystems exposed to elevated atmospheric CO2. Oecologia 2004, 140, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.F.; Price, K.P.; Ellis, J.; Shi, P.J. Response of seasonal vegetation development to climatic variations in eastern central Asia. Remote Sens. Environ. 2003, 87, 42–54. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Barbero, R.; Wolf, J.W.; Holden, Z.A. Tracking Interannual Streamflow Variability with Drought Indices in the U.S. Pacific Northwest. J. Hydrometeorol. 2014, 15, 1900–1912. [Google Scholar] [CrossRef]
- Shook, K.; Pomeroy, J.; van der Kamp, G. The transformation of frequency distributions of winter precipitation to spring streamflow probabilities in cold regions; case studies from the Canadian Prairies. J. Hydrol. 2015, 521, 395–409. [Google Scholar] [CrossRef]
- Ma, X.L.; Huete, A.; Moran, S.; Ponce-Campos, G.; Eamus, D. Abrupt shifts in phenology and vegetation productivity under climate extremes. J. Geophys. Res. Biogeosci. 2015, 120, 2036–2052. [Google Scholar] [CrossRef]
- Middleton, N.J.; Thomas, D.S. World Atlas of Desertification; United Nations Environment Programme: Nairobi, Kenya, 1992. [Google Scholar]
- Thornthwaite, C.W. An approach toward a rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- Sun, Z.G.; Wang, Q.X.; Xiao, Q.G.; Batkhishig, O.; Watanabe, M. Diverse Responses of Remotely Sensed Grassland Phenology to Interannual Climate Variability over Frozen Ground Regions in Mongolia. Remote Sens. 2015, 7, 360–377. [Google Scholar] [CrossRef]
- Abbas, S.; Qamer, F.M.; Murthy, M.S.R.; Tripathi, N.K.; Ning, W.; Sharma, E.; Ali, G. Grassland Growth in Response to Climate Variability in the Upper Indus Basin, Pakistan. Climate 2015, 3, 697–714. [Google Scholar] [CrossRef]
- Dahlgren, J.P.; von Zeipel, H.; Ehrlen, J. Variation in vegetative and flowering phenology in a forest herb caused by environmental heterogeneity. Am. J. Bot. 2007, 94, 1570–1576. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ecoregions | Grassland Areas (km2) | Grassland Compositional Proportions (%) | Major Grass Species |
|---|---|---|---|
| Aspen parkland | 128 | 0.2 | Rough Fescue (Festuca scabrella) |
| Moist Mixed Grassland | 7680 | 8.9 | Western Porcupine Grass (Stipa cur-tiseta), Northern and Western Wheatgrass (Agropyron cristatum & Pascopyrum smithii), and Green Needlegrass (Nassella viridula) |
| Mixed Grassland | 71,744 | 83.0 | Western Wheatgrass (Pascopyrum smithii), Needle-and-Thread Grass (Hesperostipa comata), June Grass (Koeleria macrantha), Blue Grama (Bouteloua gracilis), and Northern Wheatgrass (Agropyron cristatum) |
| Fescue Grassland | 2048 | 2.3 | Rough Fescue (Festuca scabrella) |
| Cypress Upland | 4800 | 5.6 | Rough Fescue (Festuca scabrella) |
| Moist Mixed Grassland | Fescue Grassland | Mixed Grassland | Cypress Upland | |||||
|---|---|---|---|---|---|---|---|---|
| r | Adjrsq | r | Adjrsq | r | Adjrsq | r | Adjrsq | |
| SOG | −0.40 * | 0.14 | −0.41 * | 0.14 | −0.59 *** | 0.33 | −0.50 ** | 0.23 |
| EOG | 0.57 *** | 0.31 | 0.68 *** | 0.44 | 0.60 *** | 0.34 | 0.53 ** | 0.26 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, T.; Martz, L.; Guo, X. Grassland Phenology Response to Drought in the Canadian Prairies. Remote Sens. 2017, 9, 1258. https://doi.org/10.3390/rs9121258
Cui T, Martz L, Guo X. Grassland Phenology Response to Drought in the Canadian Prairies. Remote Sensing. 2017; 9(12):1258. https://doi.org/10.3390/rs9121258
Chicago/Turabian StyleCui, Tengfei, Lawrence Martz, and Xulin Guo. 2017. "Grassland Phenology Response to Drought in the Canadian Prairies" Remote Sensing 9, no. 12: 1258. https://doi.org/10.3390/rs9121258
APA StyleCui, T., Martz, L., & Guo, X. (2017). Grassland Phenology Response to Drought in the Canadian Prairies. Remote Sensing, 9(12), 1258. https://doi.org/10.3390/rs9121258