A Combination of Coffee Compounds Shows Insulin-Sensitizing and Hepatoprotective Effects in a Rat Model of Diet-Induced Metabolic Syndrome

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. Diet
2.3. Intervention
2.4. Chronic Study Design
2.5. Acute Study Design
2.6. Immunoassays and Reagents
2.7. Histopathological Studies
2.8. Hyperpolarized-MR Spectroscopy Examination
2.9. Liquid Chromatography Tandem Mass Spectrometry (LC-MS/MS)
2.10. Statistical Methods
3. Results
3.1. The Chronic Study
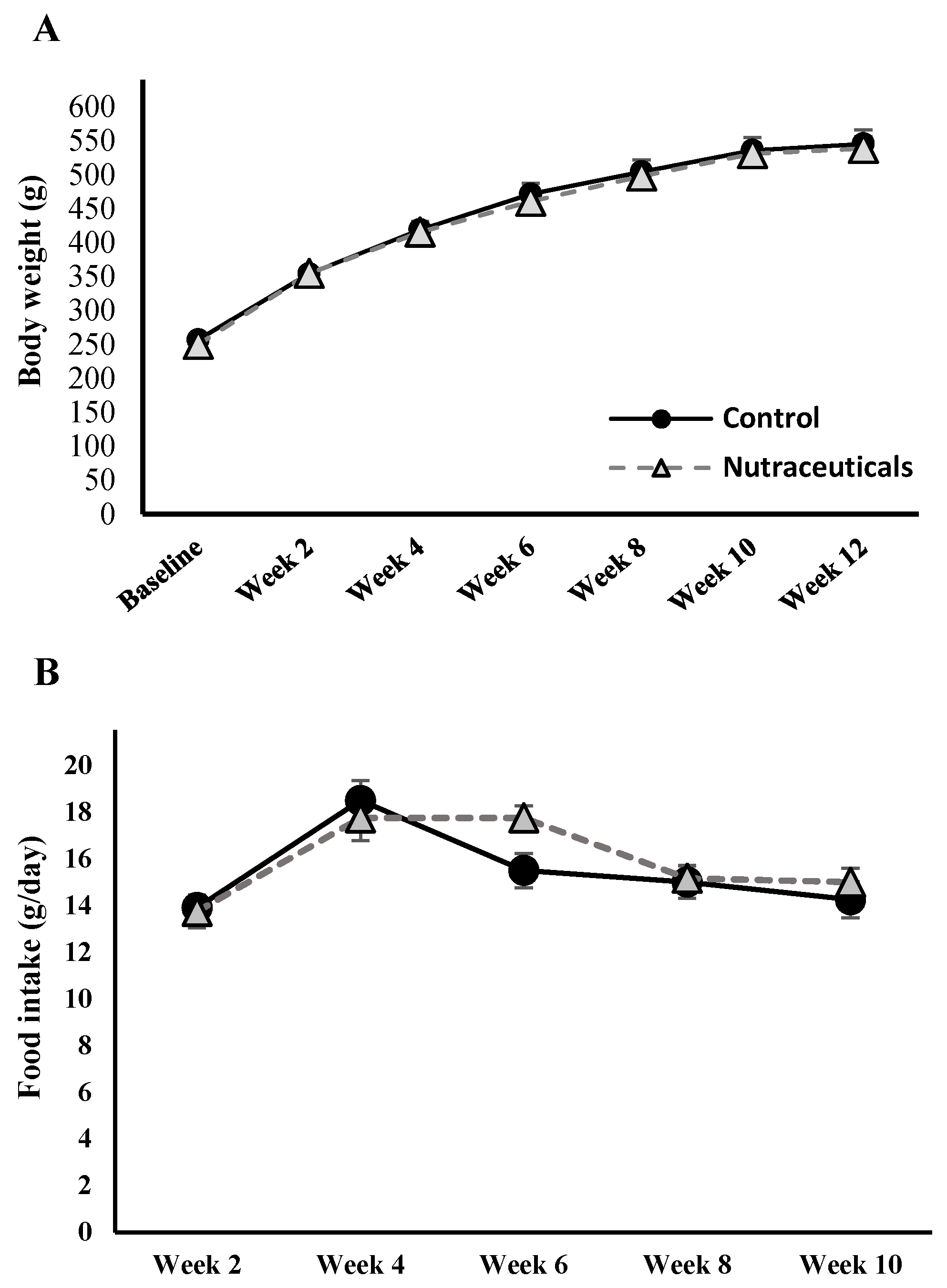
3.1.1. Energy Homeostasis
3.1.2. Blood Glucose and IR
3.1.3. Plasma Lipids and Cytokines
3.1.4. Liver Steatosis
3.1.5. Hyperpolarized-[1-13C]Pyruvate MR Spectroscopy
3.1.6. Plasma Levels of Caffeic acid Metabolites
3.2. Acute Study
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Obesity—Situation and Trends. Available online: http://www.who.int/gho/ncd/risk_factors/obesity_text/en/ (accessed on 15 September 2017).
- Virtue, S.; Vidal-Puig, A. Adipose tissue expandability, lipotoxicity and the metabolic syndrome—An allostatic perspective. Biochim. Biophys. Acta 2010, 1801, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, P.; Boulanger, M.C.; Despres, J.P. Ectopic visceral fat: A clinical and molecular perspective on the cardiometabolic risk. Rev. Endocr. Metab. Disord. 2014, 15, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Heymsfield, S.B.; Wadden, T.A. Mechanisms, pathophysiology, and management of obesity. N. Engl. J. Med. 2017, 376, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Godos, J.; Galvano, F.; Giovannucci, E.L. Coffee, caffeine, and health outcomes: An umbrella review. Annu. Rev. Nutr. 2017, 37, 131–156. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Micek, A.; Godos, J.; Sciacca, S.; Pajak, A.; Martinez-Gonzalez, M.A.; Giovannucci, E.L.; Galvano, F. Coffee consumption and risk of all-cause, cardiovascular, and cancer mortality in smokers and non-smokers: A dose-response meta-analysis. Eur. J. Epidemiol. 2016, 31, 1191–1205. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.M.; Lima, D.R. Coffee consumption, obesity and type 2 diabetes: A mini-review. Eur. J. Nutr. 2016, 55, 1345–1358. [Google Scholar] [CrossRef] [PubMed]
- Marventano, S.; Salomone, F.; Godos, J.; Pluchinotta, F.; Del Rio, D.; Mistretta, A.; Grosso, G. Coffee and tea consumption in relation with non-alcoholic fatty liver and metabolic syndrome: A systematic review and meta-analysis of observational studies. Clin. Nutr. 2016, 35, 1269–1281. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Bhupathiraju, S.N.; Chen, M.; van Dam, R.M.; Hu, F.B. Caffeinated and decaffeinated coffee consumption and risk of type 2 diabetes: A systematic review and a dose-response meta-analysis. Diabetes Care 2014, 37, 569–586. [Google Scholar] [CrossRef] [PubMed]
- Song, S.J.; Choi, S.; Park, T. Decaffeinated green coffee bean extract attenuates diet-induced obesity and insulin resistance in mice. Evid.-Based Complement. Altern. Med. 2014, 2014, 718379. [Google Scholar] [CrossRef] [PubMed]
- Tajik, N.; Tajik, M.; Mack, I.; Enck, P. The potential effects of chlorogenic acid, the main phenolic components in coffee, on health: A comprehensive review of the literature. Eur. J. Nutr. 2017. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.M.P.; Rathinasabapathy, T.; Esposito, D.; Komarnytsky, S. Structural constraints and importance of caffeic acid moiety for anti-hyperglycemic effects of caffeoylquinic acids from chicory. Mol. Nutr. Food Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Lafay, S.; Morand, C.; Manach, C.; Besson, C.; Scalbert, A. Absorption and metabolism of caffeic acid and chlorogenic acid in the small intestine of rats. Br. J. Nutr. 2006, 96, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Mellbye, F.B.; Jeppesen, P.B.; Hermansen, K.; Gregersen, S. Cafestol, a bioactive substance in coffee, stimulates insulin secretion and increases glucose uptake in muscle cells: Studies in vitro. J. Nat. Prod. 2015, 78, 2447–2451. [Google Scholar] [CrossRef] [PubMed]
- Mellbye, F.B.; Jeppesen, P.B.; Shokouh, P.; Laustsen, C.; Hermansen, K.; Gregersen, S. Cafestol, a bioactive substance in coffee, has antidiabetic properties in KKAy mice. J. Nat. Prod. 2017. [Google Scholar] [CrossRef] [PubMed]
- Lozano, I.; Van der Werf, R.; Bietiger, W.; Seyfritz, E.; Peronet, C.; Pinget, M.; Jeandidier, N.; Maillard, E.; Marchioni, E.; Sigrist, S.; et al. High-fructose and high-fat diet-induced disorders in rats: Impact on diabetes risk, hepatic and vascular complications. Nutr. Metab. 2016, 13, 15. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Jouihan, H. Measurement of liver triglyceride content. Bio-Protocol 2012, 2. [Google Scholar] [CrossRef]
- Laustsen, C.; Nielsen, P.M.; Norlinger, T.S.; Qi, H.; Pedersen, U.K.; Bertelsen, L.B.; Ostergaard, J.A.; Flyvbjerg, A.; Ardenkjaer-Larsen, J.H.; Palm, F.; et al. Antioxidant treatment attenuates lactate production in diabetic nephropathy. Am. J. Physiol. Ren. Physiol. 2017, 312, F192–F199. [Google Scholar] [CrossRef] [PubMed]
- Norskov, N.P.; Hedemann, M.S.; Theil, P.K.; Fomsgaard, I.S.; Laursen, B.B.; Knudsen, K.E. Phenolic acids from wheat show different absorption profiles in plasma: A model experiment with catheterized pigs. J. Agric. Food Chem. 2013, 61, 8842–8850. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.J.; Zhu, Q.; Zhong, Y.L.; Xu, S.H.; Wang, Z. Chlorogenic acid maintains glucose homeostasis through modulating the expression of SGLT-1, GLUT-2, and PLG in different intestinal segments of sprague-dawley rats fed a high-fat diet. Biomed. Environ. Sci. BES 2015, 28, 894–903. [Google Scholar] [PubMed]
- Mubarak, A.; Hodgson, J.M.; Considine, M.J.; Croft, K.D.; Matthews, V.B. Supplementation of a high-fat diet with chlorogenic acid is associated with insulin resistance and hepatic lipid accumulation in mice. J. Agric. Food Chem. 2013, 61, 4371–4378. [Google Scholar] [CrossRef] [PubMed]
- Li Kwok Cheong, J.D.; Croft, K.D.; Henry, P.D.; Matthews, V.; Hodgson, J.M.; Ward, N.C. Green coffee polyphenols do not attenuate features of the metabolic syndrome and improve endothelial function in mice fed a high fat diet. Arch. Biochem. Biophys. 2014, 559, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.; Varghese, M.; Wang, J.; Zhao, W.; Chen, F.; Knable, L.A.; Ferruzzi, M.; Pasinetti, G.M. Dietary supplementation with decaffeinated green coffee improves diet-induced insulin resistance and brain energy metabolism in mice. Nutr. Neurosci. 2012, 15, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Aw, W.; Egashira, K.; Takahashi, S.; Aoyama, S.; Saito, K.; Kishimoto, Y.; Kato, H. Coffee intake mitigated inflammation and obesity-induced insulin resistance in skeletal muscle of high-fat diet-induced obese mice. Genes Nutr. 2014, 9, 389. [Google Scholar] [CrossRef] [PubMed]
- Shearer, J.; Sellars, E.A.; Farah, A.; Graham, T.E.; Wasserman, D.H. Effects of chronic coffee consumption on glucose kinetics in the conscious rat. Can. J. Physiol. Pharmacol. 2007, 85, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Ippoushi, K.; Nakayama, M.; Ito, H.; Higashio, H.; Terao, J. Absorption of chlorogenic acid and caffeic acid in rats after oral administration. J. Agric. Food Chem. 2000, 48, 5496–5500. [Google Scholar] [CrossRef] [PubMed]
- Achari, A.E.; Jain, S.K. Adiponectin, a therapeutic target for obesity, diabetes, and endothelial dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Sasaki, S.; Uenishi, K. Serum adiponectin concentration in relation to macronutrient and food intake in young Japanese women. Nutrition 2013, 29, 1315–1320. [Google Scholar] [CrossRef] [PubMed]
- Rudich, A.; Tirosh, A.; Potashnik, R.; Hemi, R.; Kanety, H.; Bashan, N. Prolonged oxidative stress impairs insulin-induced GLUT4 translocation in 3T3-L1 adipocytes. Diabetes 1998, 47, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Gulcin, I. Antioxidant activity of caffeic acid (3,4-dihydroxycinnamic acid). Toxicology 2006, 217, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Tunnicliffe, J.M.; Eller, L.K.; Reimer, R.A.; Hittel, D.S.; Shearer, J. Chlorogenic acid differentially affects postprandial glucose and glucose-dependent insulinotropic polypeptide response in rats. Appl. Physiol. Nutr. Metab. 2011, 36, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Hsu, F.L.; Chen, Y.C.; Cheng, J.T. Caffeic acid as active principle from the fruit of xanthium strumarium to lower plasma glucose in diabetic rats. Planta Med. 2000, 66, 228–230. [Google Scholar] [CrossRef] [PubMed]
- Murase, T.; Misawa, K.; Minegishi, Y.; Aoki, M.; Ominami, H.; Suzuki, Y.; Shibuya, Y.; Hase, T. Coffee polyphenols suppress diet-induced body fat accumulation by downregulating SREBP-1c and related molecules in C57BL/6J mice. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E122–E133. [Google Scholar] [CrossRef] [PubMed]
- Beynen, A.C.; Weusten-Van der Wouw, M.P.; de Roos, B.; Katan, M.B. Boiled coffee fails to raise serum cholesterol in hamsters and rats. Br. J. Nutr. 1996, 76, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Vitaglione, P.; Morisco, F.; Mazzone, G.; Amoruso, D.C.; Ribecco, M.T.; Romano, A.; Fogliano, V.; Caporaso, N.; D’Argenio, G. Coffee reduces liver damage in a rat model of steatohepatitis: The underlying mechanisms and the role of polyphenols and melanoidins. Hepatology 2010, 52, 1652–1661. [Google Scholar] [CrossRef] [PubMed]
- Ardenkjaer-Larsen, J.H.; Fridlund, B.; Gram, A.; Hansson, G.; Hansson, L.; Lerche, M.H.; Servin, R.; Thaning, M.; Golman, K. Increase in signal-to-noise ratio of >10,000 times in liquid-state NMR. Proc. Natl. Acad. Sci. USA 2003, 100, 10158–10163. [Google Scholar] [CrossRef] [PubMed]
- Moon, C.M.; Oh, C.H.; Ahn, K.Y.; Yang, J.S.; Kim, J.Y.; Shin, S.S.; Lim, H.S.; Heo, S.H.; Seon, H.J.; Kim, J.W.; et al. Metabolic biomarkers for non-alcoholic fatty liver disease induced by high-fat diet: In vivo magnetic resonance spectroscopy of hyperpolarized [1-13C] pyruvate. Biochem. Biophys. Res. Commun. 2017, 482, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.J.; Miller, J.J.; McCallum, C.; Rider, O.J.; Neubauer, S.; Heather, L.C.; Tyler, D.J. Assessment of metformin-induced changes in cardiac and hepatic redox state using hyperpolarized[1-13C]pyruvate. Diabetes 2016, 65, 3544–3551. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Chen, A.P.; Zierhut, M.L.; Bok, R.; Yen, Y.F.; Schroeder, M.A.; Hurd, R.E.; Nelson, S.J.; Kurhanewicz, J.; Vigneron, D.B. In vivo carbon-13 dynamic MRS and MRSI of normal and fasted rat liver with hyperpolarized 13C-pyruvate. Mol. Imaging Biol. MIB 2009, 11, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Keijzers, G.B.; De Galan, B.E.; Tack, C.J.; Smits, P. Caffeine can decrease insulin sensitivity in humans. Diabetes Care 2002, 25, 364–369. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Group | Nutraceuticals Group | p-Value of Change/Difference | |||
|---|---|---|---|---|---|
| Baseline | 12 Weeks | Baseline | 12 Weeks | ||
| Plasma lipids (mmol/L) | |||||
| Total cholesterol | 1.87 (0.08) | 2.00 (0.08) | 2.02 (0.10) | 2.33 (0.16) | 0.428 |
| HDL-C | 1.42 (0.05) | 1.58 (0.06) | 1.55 (0.07) | 1.83 (0.11) | 0.492 |
| LDL-C | 0.48 (0.03) | 0.36 (0.03) | 0.47 (0.03) | 0.49 (0.07) | 0.097 |
| Triglyceride | 0.36 (0.02) | 0.42 (0.05) | 0.37 (0.01) | 0.39 (0.04) | 0.675 |
| NEFAs | 1.01 (0.08) | 0.31 (0.03) | 0.91 (0.05) | 0.31 (0.02) | 0.268 |
| HbA1C (mmol/mol) | - | 23.08 (0.29) | - | 23.55 (0.55) | 0.451 |
| Plasma adiponectin (µg/mL) | - | 4.40 (0.28) | - | 5.39 (0.39) * | 0.048 |
| Plasma IL1b (pg/mL) | - | 13.65 (1.88) | - | 8.92 (2.46) | 0.139 |
| Plasma ALT (U/L) | 31.20 (0.73) | 27.72 (1.39) | 32.78 (0.71) | 22.97 (1.09) * | 0.004 |
| Liver triglycerides content (mmol/L) | - | 2.00 (0.26) | - | 1.62 (0.17) | 0.223 |
| Model Species | Intervention Duration (Weeks) | Intervention | Dosage (mg/Day) | Human Equivalent Dosage | |
|---|---|---|---|---|---|
| Present study | Sprague-Dawley rat | 12 | Chemicals combination | 51 | 1428 |
| CA | 30 | 840 | |||
| Song et al., 2014 [10] | C57BL/6N mouse | 11 | DCBE | 3 | 507 |
| 9 | 1521 | ||||
| 27 | 4563 | ||||
| CGA | 4.5 | 761 | |||
| Peng et al., 2015 [21] | Sprague-Dawley rat | 12 | CGA | 7 | 196 |
| 32 | 896 | ||||
| Mubarak et al., 2013 [22] | C57BL6 mouse | 12 | CGA | 0.3 | 51 |
| Li Kwok Cheong et al., 2014 [23] | C57BL6 mouse | 12 | DCBE | 15 | 2535 |
| Ho et al., 2012 [24] | C57B6SJL mouse | 20 | DCBE | 2.8 | 473 |
| Jia et al., 2014 [25] | C57BL/6J mouse | 9 | DCBE | 60 | 10,140 |
| Shearer et al., 2007 [26] | Sprague-Dawley rat | 4 | DCBE | 1600 | 44,800 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shokouh, P.; Jeppesen, P.B.; Hermansen, K.; Nørskov, N.P.; Laustsen, C.; Jacques Hamilton-Dutoit, S.; Qi, H.; Stødkilde-Jørgensen, H.; Gregersen, S. A Combination of Coffee Compounds Shows Insulin-Sensitizing and Hepatoprotective Effects in a Rat Model of Diet-Induced Metabolic Syndrome. Nutrients 2018, 10, 6. https://doi.org/10.3390/nu10010006
Shokouh P, Jeppesen PB, Hermansen K, Nørskov NP, Laustsen C, Jacques Hamilton-Dutoit S, Qi H, Stødkilde-Jørgensen H, Gregersen S. A Combination of Coffee Compounds Shows Insulin-Sensitizing and Hepatoprotective Effects in a Rat Model of Diet-Induced Metabolic Syndrome. Nutrients. 2018; 10(1):6. https://doi.org/10.3390/nu10010006
Chicago/Turabian StyleShokouh, Pedram, Per Bendix Jeppesen, Kjeld Hermansen, Natalja P. Nørskov, Christoffer Laustsen, Stephen Jacques Hamilton-Dutoit, Haiyun Qi, Hans Stødkilde-Jørgensen, and Søren Gregersen. 2018. "A Combination of Coffee Compounds Shows Insulin-Sensitizing and Hepatoprotective Effects in a Rat Model of Diet-Induced Metabolic Syndrome" Nutrients 10, no. 1: 6. https://doi.org/10.3390/nu10010006
APA StyleShokouh, P., Jeppesen, P. B., Hermansen, K., Nørskov, N. P., Laustsen, C., Jacques Hamilton-Dutoit, S., Qi, H., Stødkilde-Jørgensen, H., & Gregersen, S. (2018). A Combination of Coffee Compounds Shows Insulin-Sensitizing and Hepatoprotective Effects in a Rat Model of Diet-Induced Metabolic Syndrome. Nutrients, 10(1), 6. https://doi.org/10.3390/nu10010006