Lactobacillus rhamnosus GG Protects the Epithelial Barrier of Wistar Rats from the Pepsin-Trypsin-Digested Gliadin (PTG)-Induced Enteropathy




Abstract
:1. Introduction
2. Materials and Methods
2.1. Gliadin Digest
2.2. Animals and Experimental Design
2.3. Histologic and Morphometric Evaluation
2.4. Polyamine Analysis
2.5. ODC Activity
2.6. Real-Time PCR
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
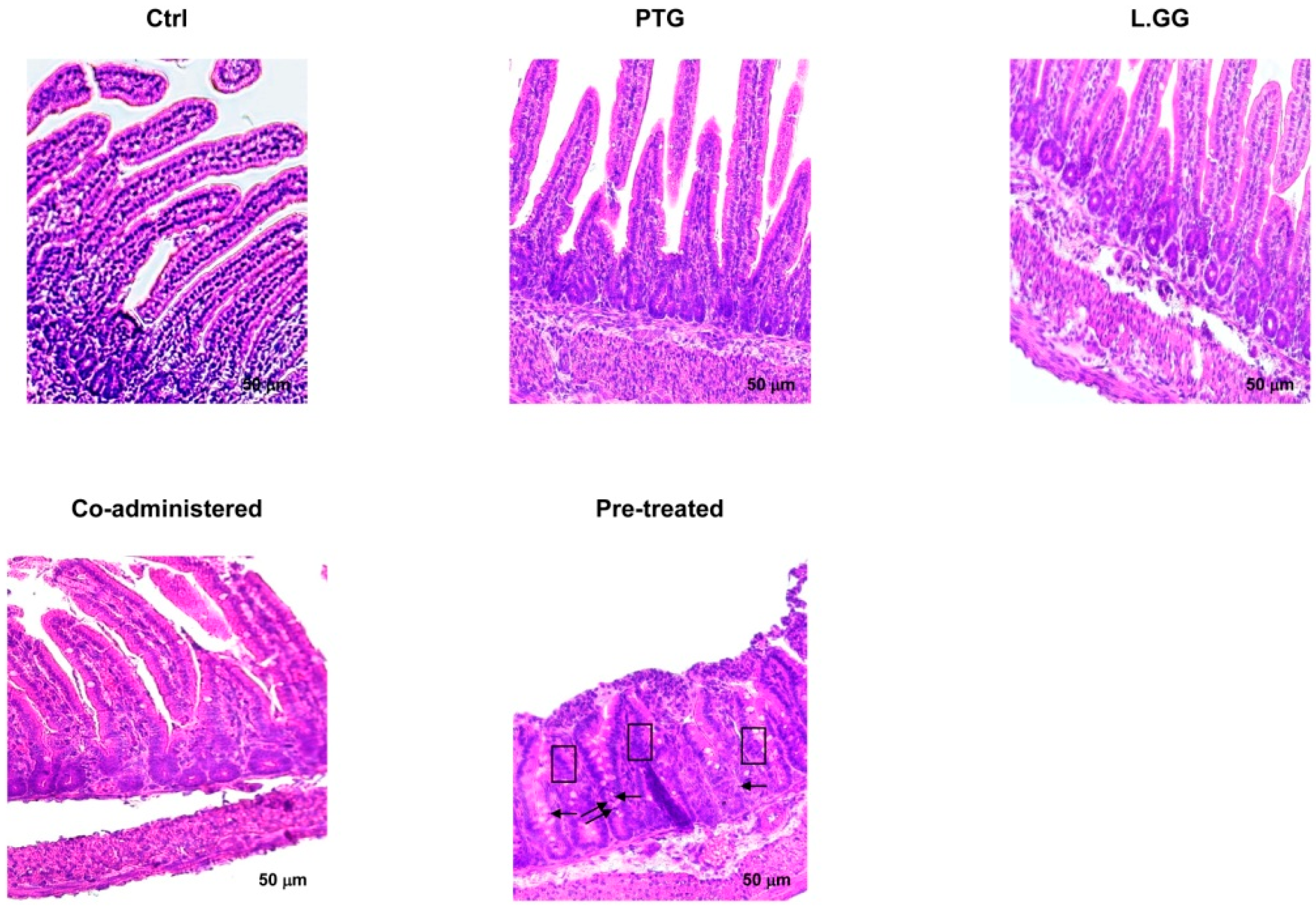
3.1. Morphometric Analysis
3.2. Polyamine Profile
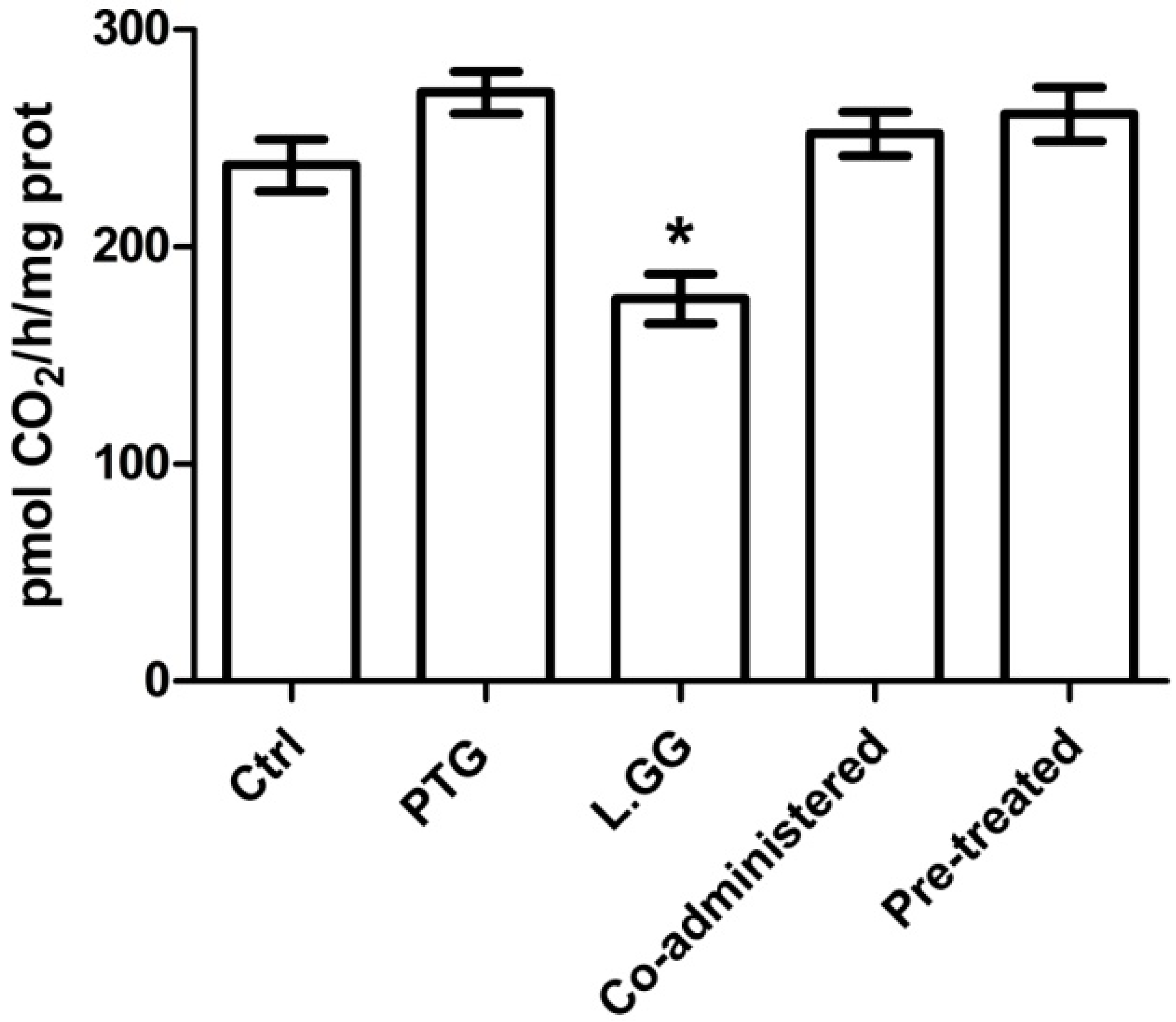
3.3. ODC Activity
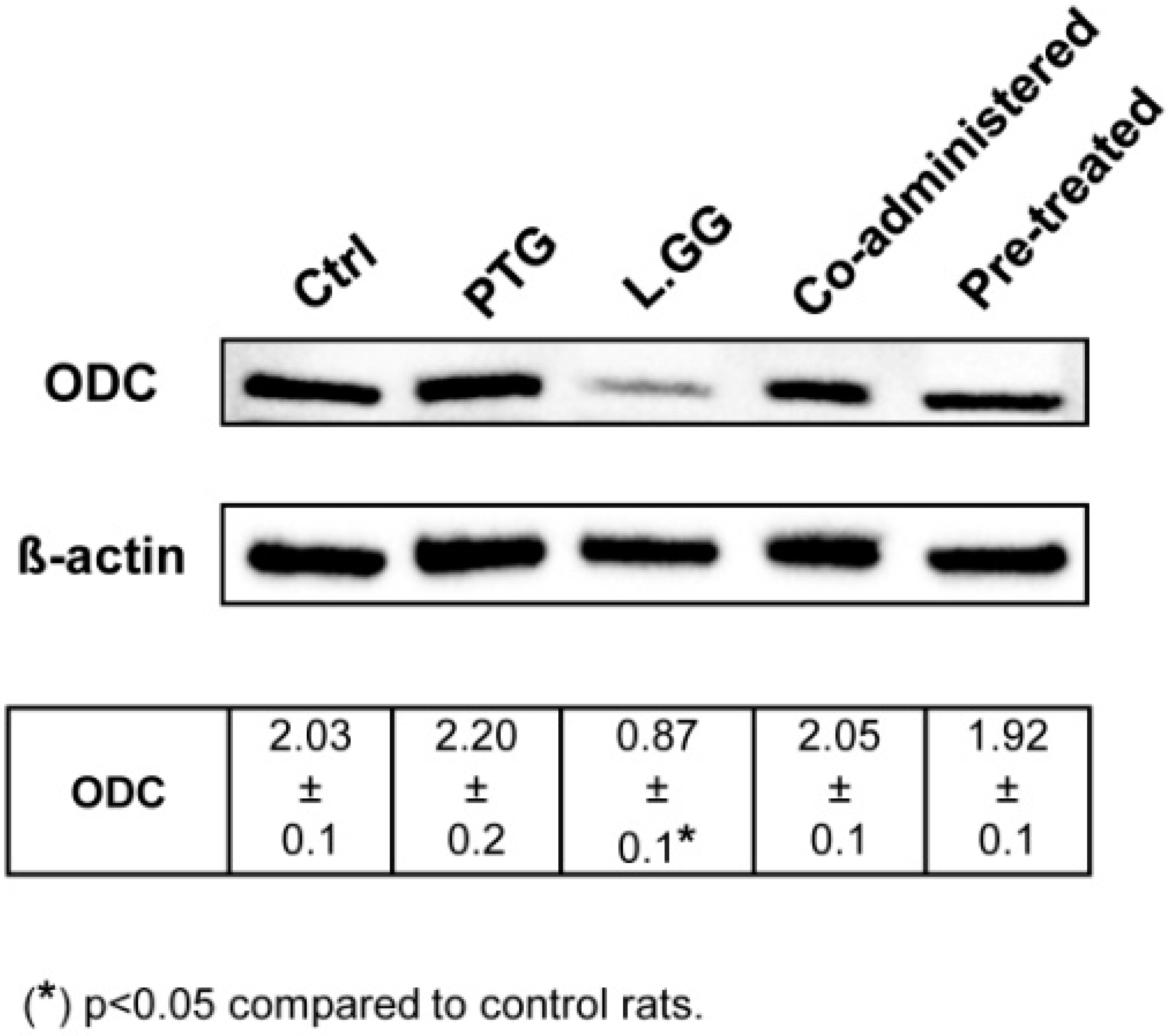
3.4. ODC Expression
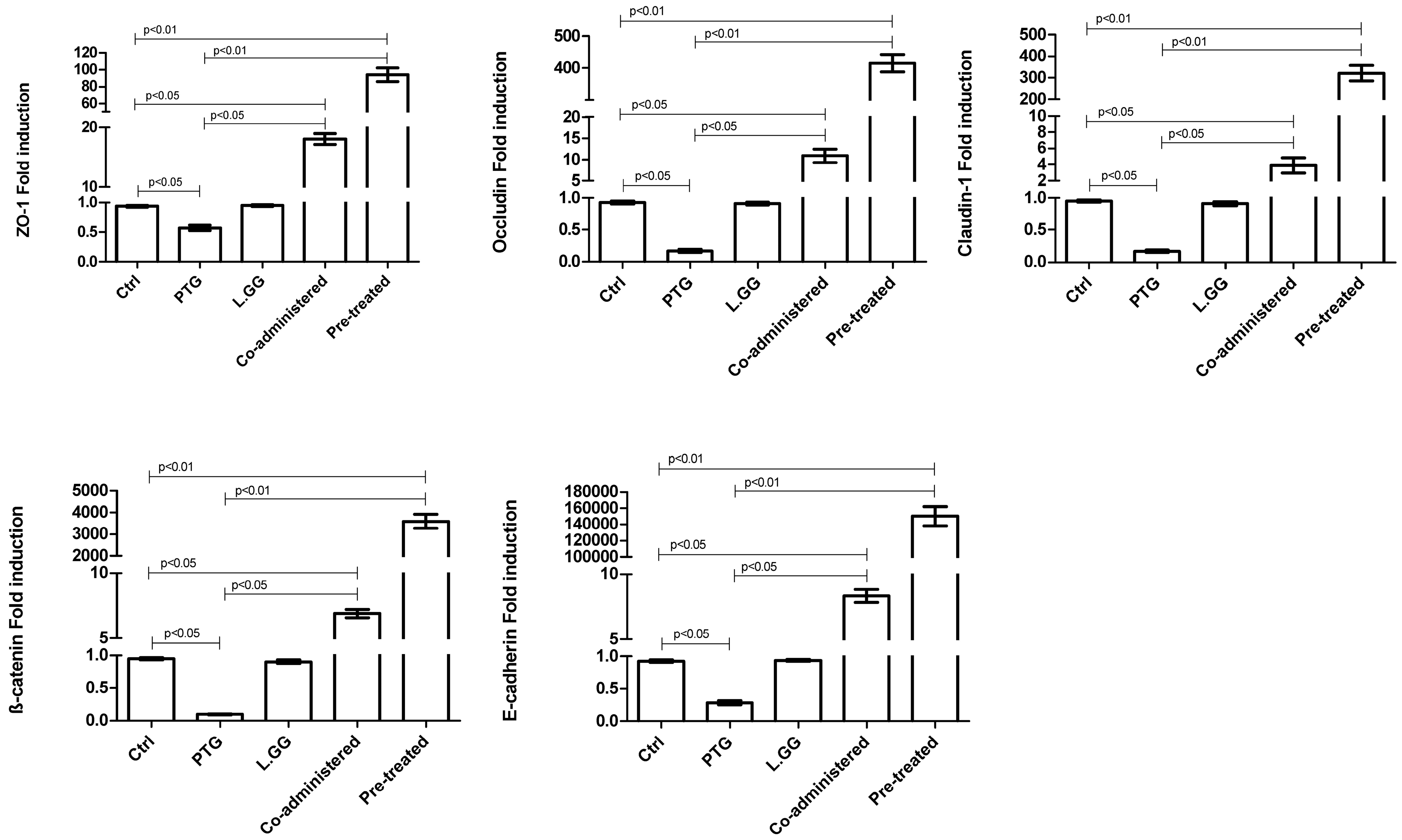
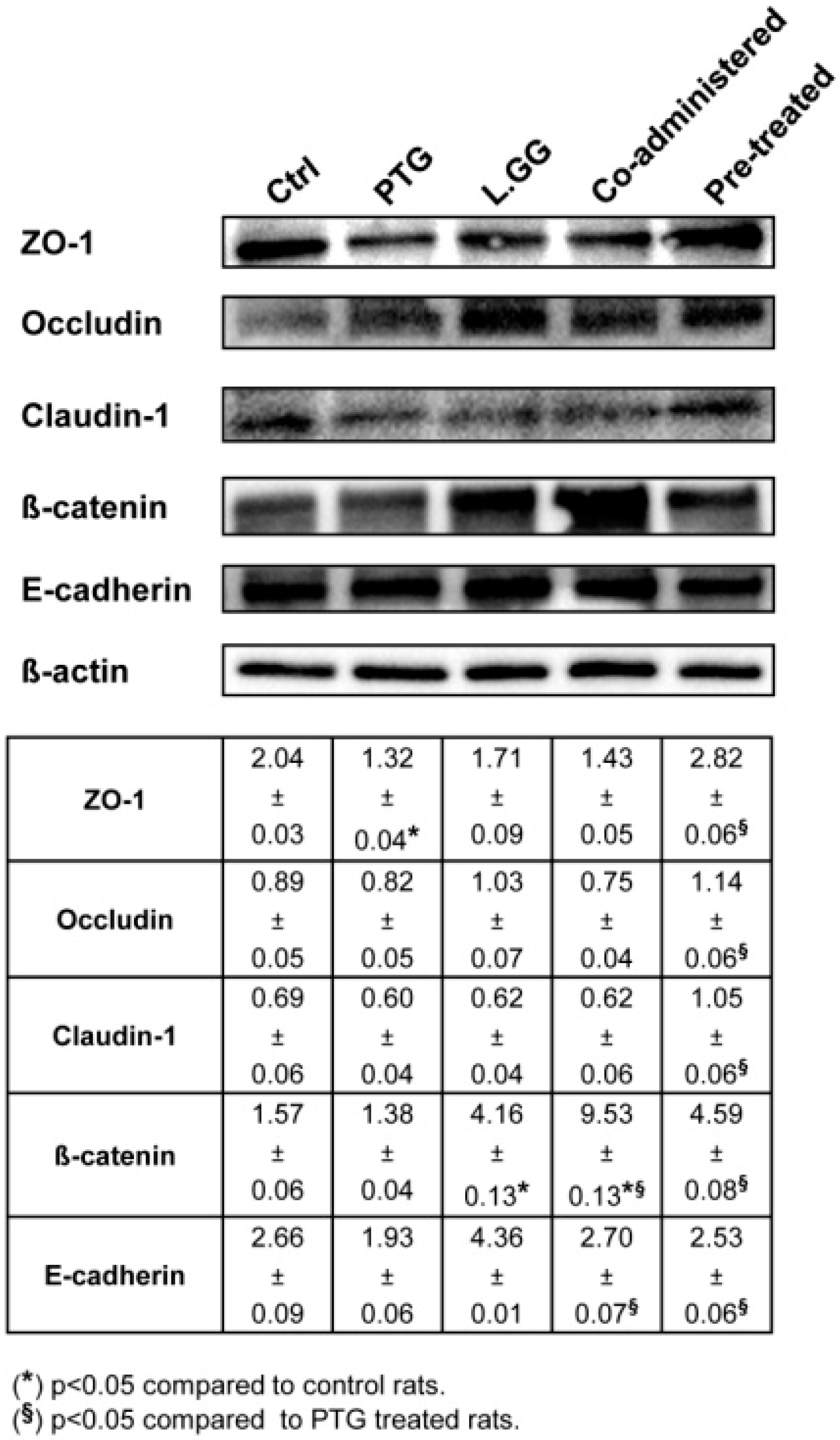
3.5. Intercellular Junction Protein Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cukrowska, B.; Sowinska, A.; Bierla, J.B.; Czarnowska, E.; Rybak, A.; Grzybowska-Chlebowczyk, U. Intestinal epithelium, intraepithelial lymphocytes and the gut microbiota—Key players in the pathogenesis of celiac disease. World J. Gastroenterol. 2017, 23, 7505–7518. [Google Scholar] [CrossRef] [PubMed]
- Cooper, B.T. Intestinal permeability in coeliac disease. Lancet 1983, 1, 658–659. [Google Scholar] [CrossRef]
- Molberg, O.; McAdam, S.N.; Korner, R.; Quarsten, H.; Kristiansen, C.; Madsen, L.; Fugger, L.; Scott, H.; Noren, O.; Roepstorff, P.; et al. Tissue transglutaminase selectively modifies gliadin peptides that are recognized by gut-derived t cells in celiac disease. Nat. Med. 1998, 4, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability—A new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef] [PubMed]
- Campbell, H.K.; Maiers, J.L.; DeMali, K.A. Interplay between tight junctions & adherens junctions. Exp. Cell Res. 2017, 358, 39–44. [Google Scholar] [PubMed]
- Van Itallie, C.M.; Anderson, J.M. Architecture of tight junctions and principles of molecular composition. Semin. Cell Dev. Biol. 2014, 36, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabor, C.W.; Tabor, H. Polyamines. Annu. Rev. Biochem. 1984, 53, 749–790. [Google Scholar] [CrossRef] [PubMed]
- Timmons, J.; Chang, E.T.; Wang, J.Y.; Rao, J.N. Polyamines and Gut Mucosal Homeostasis. J. Gastrointest. Dig. Syst. 2012, 2 (Suppl. 7), 001. [Google Scholar] [CrossRef] [PubMed]
- Madsen, K.; Cornish, A.; Soper, P.; McKaigney, C.; Jijon, H.; Yachimec, C.; Doyle, J.; Jewell, L.; De Simone, C. Probiotic bacteria enhance murine and human intestinal epithelial barrier function. Gastroenterology 2001, 121, 580–591. [Google Scholar] [CrossRef] [PubMed]
- Zareie, M.; Johnson-Henry, K.; Jury, J.; Yang, P.C.; Ngan, B.Y.; McKay, D.M.; Soderholm, J.D.; Perdue, M.H.; Sherman, P.M. Probiotics prevent bacterial translocation and improve intestinal barrier function in rats following chronic psy chological stress. Gut 2006, 55, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.J.; Wells, J.M. Regulation of human epithelial tight junction proteins by lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, E.; O’Callaghan, J.; Butto, L.F.; Hurley, G.; Melgar, S.; Tanabe, S.; Shanahan, F.; Nally, K.; O’Toole, P.W. Mechanism of protection of transepithelial barrier function by lactobacillus salivarius: Strain dependence and attenuation by bacteriocin production. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1029–G1041. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Y.; Sidhu, A.; Ma, Z.; McClain, C.; Feng, W. Lactobacillus rhamnosus GG culture supernatant ameliorates acute alcohol-induced intestinal permeability and liver injury. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G32–G41. [Google Scholar] [CrossRef] [PubMed]
- Zakostelska, Z.; Kverka, M.; Klimesova, K.; Rossmann, P.; Mrazek, J.; Kopecny, J.; Hornova, M.; Srutkova, D.; Hudcovic, T.; Ridl, J.; et al. Lysate of probiotic lactobacillus casei DN-114 001 ameliorates colitis by strengthening the gut barrier function and changing the gut microenvironment. PLoS ONE 2011, 6, e27961. [Google Scholar] [CrossRef] [PubMed]
- Wagnerberger, S.; Spruss, A.; Kanuri, G.; Stahl, C.; Schroder, M.; Vetter, W.; Bischoff, S.C.; Bergheim, I. Lactobacillus casei shirota protects from fructose-induced liver steatosis: A mouse model. J. Nutr. Biochem. 2013, 24, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Parmar, A.; Greco, D.; Venalainen, J.; Gentile, M.; Dukes, E.; Saavalainen, P. Gene expression profiling of gliadin effects on intestinal epithelial cells suggests novel non-enzymatic functions of pepsin and trypsin. PLoS ONE 2013, 8, e66307. [Google Scholar] [CrossRef] [PubMed]
- Orlando, A.; Linsalata, M.; Notarnicola, M.; Tutino, V.; Russo, F. Lactobacillus GG restoration of the gliadin induced epithelial barrier disruption: The role of cellular polyamines. BMC Microbiol. 2014, 14, 19. [Google Scholar] [CrossRef] [PubMed]
- Orlando, A.; Linsalata, M.; D’Attoma, B.; Russo, F. Changes in paracellular permeability induced by pepsin-trypsin digested gliadin (PTG): Role of polyamines in the lactobacillus rhamnosus GG protective action. J. Funct. Foods 2017, 36, 52–62. [Google Scholar] [CrossRef]
- Troncone, R.; Ferguson, A. Animal model of gluten induced enteropathy in mice. Gut 1991, 32, 871–875. [Google Scholar] [CrossRef] [PubMed]
- Stepankova, R.; Tlaskalova-Hogenova, H.; Sinkora, J.; Jodl, J.; Fric, P. Changes in jejunal mucosa after long-term feeding of germfree rats with gluten. Scand. J. Gastroenterol. 1996, 31, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Kozakova, H.; Stepankova, R.; Tuckova, L.; Sinkora, M.; Jelinkova, L.; Tlaskalova-Hogenova, H. Humoral and cellular immune responses in gluten-treated suckling or hand-fed rats. Physiol. Res. 2000, 49, 665–672. [Google Scholar] [PubMed]
- Tlaskalova-Hogenova, H.; Stepankova, R.; Farre, M.; Funda, D.P.; Rehakova, Z.; Sinkora, J.; Tuckova, L.; Horak, I.; Horakova, D.; Cukrowska, B.; et al. Autoimmune reactions induced by gliadin feeding in germ-free avn rats and athymic nude mice. Animal models for celiac disease. Ann. N. Y. Acad. Sci. 1997, 815, 503–505. [Google Scholar] [CrossRef] [PubMed]
- Black, K.E.; Murray, J.A.; David, C.S. HLA-DQ determines the response to exogenous wheat proteins: A model of gluten sensitivity in transgenic knockout mice. J. Immunol. 2002, 169, 5595–5600. [Google Scholar] [CrossRef] [PubMed]
- Verdu, E.F.; Huang, X.; Natividad, J.; Lu, J.; Blennerhassett, P.A.; David, C.S.; McKay, D.M.; Murray, J.A. Gliadin-dependent neuromuscular and epithelial secretory responses in gluten-sensitive hla-dq8 transgenic mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G217–G225. [Google Scholar] [CrossRef] [PubMed]
- Laparra, J.M.; Olivares, M.; Gallina, O.; Sanz, Y. Bifidobacterium longum cect 7347 modulates immune responses in a gliadin-induced enteropathy animal model. PLoS ONE 2012, 7, e30744. [Google Scholar] [CrossRef] [PubMed]
- Drago, S.; El Asmar, R.; Di Pierro, M.; Grazia Clemente, M.; Tripathi, A.; Sapone, A.; Thakar, M.; Iacono, G.; Carroccio, A.; D’Agate, C.; et al. Gliadin, zonulin and gut permeability: Effects on celiac and non-celiac intestinal mucosa and intestinal cell lines. Scand. J. Gastroenterol. 2006, 41, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Linsalata, M.; Russo, F.; Notarnicola, M.; Berloco, P.; Di Leo, A. Polyamine profile in human gastric mucosa infected by helicobacter pylori. Ital. J. Gastroenterol. Hepatol. 1998, 30, 484–489. [Google Scholar] [PubMed]
- Linsalata, M.; Orlando, A.; Tutino, V.; Notarnicola, M.; D’Attoma, B.; Russo, F. Inhibitory effect of vitamin K1 on growth and polyamine biosynthesis of human gastric and colon carcinoma cell lines. Int. J. Oncol. 2015, 47, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Niessen, C.M. Tight junctions/adherens junctions: Basic structure and function. J. Investig. Dermatol. 2007, 127, 2525–2532. [Google Scholar] [CrossRef] [PubMed]
- Sander, G.R.; Cummins, A.G.; Henshall, T.; Powell, B.C. Rapid disruption of intestinal barrier function by gliadin involves altered expression of apical junctional proteins. FEBS Lett. 2005, 579, 4851–4855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elli, L.; Roncoroni, L.; Doneda, L.; Ciulla, M.M.; Colombo, R.; Braidotti, P.; Bonura, A.; Bardella, M.T. Imaging analysis of the gliadin direct effect on tight junctions in an in vitro three-dimensional lovo cell line culture system. Toxicol. In Vitro 2011, 25, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Ciccocioppo, R.; Finamore, A.; Ara, C.; Di Sabatino, A.; Mengheri, E.; Corazza, G.R. Altered expression, localization, and phosphorylation of epithelial junctional proteins in celiac disease. Am. J. Clin. Pathol. 2006, 125, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Stepankova, R.; Kofronova, O.; Tuckova, L.; Kozakova, H.; Cebra, J.J.; Tlaskalova-Hogenova, H. Experimentally induced gluten enteropathy and protective effect of epidermal growth factor in artificially fed neonatal rats. J. Pediatr. Gastroenterol. Nutr. 2003, 36, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Barilli, A.; Rotoli, B.M.; Visigalli, R.; Dall’Asta, V. Gliadin activates arginase pathway in raw264.7 cells and in human monocytes. Biochim. Biophys. Acta 2014, 1842, 1364–1371. [Google Scholar] [CrossRef] [PubMed]
- Larque, E.; Sabater-Molina, M.; Zamora, S. Biological significance of dietary polyamines. Nutrition 2007, 23, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Powroznik, B.; Gharbi, M.; Dandrifosse, G.; Peulen, O. Enhancement of lysozyme stability and activity by polyamines. Biochimie 2004, 86, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Schulzke, J.D.; Bentzel, C.J.; Schulzke, I.; Riecken, E.O.; Fromm, M. Epithelial tight junction structure in the jejunum of children with acute and treated celiac sprue. Pediatr. Res. 1998, 43, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, A.; Stephen, S.; Borum, M.L.; Doman, D.B. Emerging therapeutic options for celiac disease: Potential alternatives to a gluten-free diet. Gastroenterol. Hepatol. (N. Y.) 2012, 8, 582–588. [Google Scholar]
- De Sousa Moraes, L.F.; Grzeskowiak, L.M.; de Sales Teixeira, T.F.; Gouveia Peluzio, M.D.C. Intestinal microbiota and probiotics in celiac disease. Clin. Microbiol. Rev. 2014, 27, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Vanderpool, C.; Yan, F.; Polk, D.B. Mechanisms of probiotic action: Implications for therapeutic applications in inflammatory bowel diseases. Inflamm. Bowel Dis. 2008, 14, 1585–1596. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Rizzello, C.G.; Fasano, A.; Clemente, M.G.; De Simone, C.; Silano, M.; De Vincenzi, M.; Losito, I.; Gobbetti, M. Vsl#3 probiotic preparation has the capacity to hydrolyze gliadin polypeptides responsible for celiac sprue. Biochim. Biophys. Acta 2006, 1762, 80–93. [Google Scholar] [PubMed]
- Claes, I.J.; Schoofs, G.; Regulski, K.; Courtin, P.; Chapot-Chartier, M.P.; Rolain, T.; Hols, P.; von Ossowski, I.; Reunanen, J.; de Vos, W.M.; et al. Genetic and biochemical characterization of the cell wall hydrolase activity of the major secreted protein of lactobacillus rhamnosus GG. PLoS ONE 2012, 7, e31588. [Google Scholar] [CrossRef] [PubMed]
- Sarno, M.; Lania, G.; Cuomo, M.; Nigro, F.; Passannanti, F.; Budelli, A.; Fasano, F.; Troncone, R.; Auricchio, S.; Barone, M.V.; et al. Lactobacillus paracasei CBA l74 interferes with gliadin peptides entrance in Caco-2 cells. Int. J. Food Sci. Nutr. 2014, 65, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Peuhkuri, K.; Vapaatalo, H.; Korpela, R. Even low-grade inflammation impacts on small intestinal function. World J. Gastroenterol. 2010, 16, 1057–1062. [Google Scholar] [CrossRef] [PubMed]
- Orlando, A.; Messa, C.; Linsalata, M.; Cavallini, A.; Russo, F. Effects of lactobacillus rhamnosus GG on proliferation and polyamine metabolism in HGC-27 human gastric and DLD-1 colonic cancer cell lines. Immunopharmacol. Immunotoxicol. 2009, 31, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.C.; Wang, J.T.; Wei, S.C.; Ni, Y.H. Host-microbial interactions and regulation of intestinal epithelial barrier function: From physiology to pathology. World J. Gastrointest. Pathophysiol. 2012, 3, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Mennigen, R.; Nolte, K.; Rijcken, E.; Utech, M.; Loeffler, B.; Senninger, N.; Bruewer, M. Probiotic mixture vsl#3 protects the epithelial barrier by maintaining tight junction protein expression and preventing apoptosis in a murine model of colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1140–G1149. [Google Scholar] [PubMed]
- Qin, H.; Zhang, Z.; Hang, X.; Jiang, Y.L. Plantarum prevents enteroinvasive Escherichia coli-induced tight junction proteins changes in intestinal epithelial cells. BMC Microbiol. 2009, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Resta-Lenert, S.; Barrett, K.E. Live probiotics protect intestinal epithelial cells from the effects of infection with enteroinvasive Escherichia coli (EIEC). Gut 2003, 52, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Ewaschuk, J.B.; Diaz, H.; Meddings, L.; Diederichs, B.; Dmytrash, A.; Backer, J.; Looijer-van Langen, M.; Madsen, K.L. Secreted bioactive factors from bifidobacterium infantis enhance epithelial cell barrier function. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G1025–G1034. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, K.; Barrett, K.E.; Resta-Lenert, S.C. Probiotic inhibition of the entry of enteroinvasive E. coli into, human intestinal epithelial cells involves both Rho-dependent and -independent pathways. Gastroenterology 2003, 124, A106. [Google Scholar] [CrossRef]
- De Boer, W.I.; Sharma, H.S.; Baelemans, S.M.; Hoogsteden, H.C.; Lambrecht, B.N.; Braunstahl, G.J. Altered expression of epithelial junctional proteins in atopic asthma: Possible role in inflammation. Can. J. Physiol. Pharmacol. 2008, 86, 105–112. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Villi | ||
|---|---|---|
| Length (µm) | Width (µm) | |
| Ctrl | 259.26 ± 1.98 | 50.59 ± 0.44 |
| PTG | 144.41 ± 2.84 ** | 45.02 ± 0.40 |
| L.GG | 260.11 ± 0.91 | 53.91 ± 0.26 |
| Co-administered | 221.64 ± 1.71 § | 54.92 ± 0.54 |
| Pre-treated | 159.14 ± 1.98 ** | 50.22 ± 0.25 |
| Putrescine | Spermidine | Spermine | Total Polyamines | |
|---|---|---|---|---|
| Ctrl | 2.32 ± 0.3 | 12.54 ± 1.0 | 7.72 ± 0.5 | 22.59 ± 1.8 |
| PTG | 1.89 ± 0.1 | 14.88 ± 0.5 | 8.42 ± 0.4 | 25.20 ± 0.7 |
| L.GG | 1.92 ± 0.2 | 11.05 ± 0.3 | 7.82 ± 0.1 | 20.79 ± 0.5 |
| Co-administered | 1.94 ± 0.2 | 10.68 ± 0.6 § | 8.53 ± 0.3 | 21.16 ± 1.1 § |
| Pre-treated | 1.86 ± 0.1 | 11.58 ± 0.7 | 7.57 ± 0.3 | 21.01 ± 0.6 § |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orlando, A.; Linsalata, M.; Bianco, G.; Notarnicola, M.; D’Attoma, B.; Scavo, M.P.; Tafaro, A.; Russo, F. Lactobacillus rhamnosus GG Protects the Epithelial Barrier of Wistar Rats from the Pepsin-Trypsin-Digested Gliadin (PTG)-Induced Enteropathy. Nutrients 2018, 10, 1698. https://doi.org/10.3390/nu10111698
Orlando A, Linsalata M, Bianco G, Notarnicola M, D’Attoma B, Scavo MP, Tafaro A, Russo F. Lactobacillus rhamnosus GG Protects the Epithelial Barrier of Wistar Rats from the Pepsin-Trypsin-Digested Gliadin (PTG)-Induced Enteropathy. Nutrients. 2018; 10(11):1698. https://doi.org/10.3390/nu10111698
Chicago/Turabian StyleOrlando, Antonella, Michele Linsalata, Giusy Bianco, Maria Notarnicola, Benedetta D’Attoma, Maria Principia Scavo, Angela Tafaro, and Francesco Russo. 2018. "Lactobacillus rhamnosus GG Protects the Epithelial Barrier of Wistar Rats from the Pepsin-Trypsin-Digested Gliadin (PTG)-Induced Enteropathy" Nutrients 10, no. 11: 1698. https://doi.org/10.3390/nu10111698
APA StyleOrlando, A., Linsalata, M., Bianco, G., Notarnicola, M., D’Attoma, B., Scavo, M. P., Tafaro, A., & Russo, F. (2018). Lactobacillus rhamnosus GG Protects the Epithelial Barrier of Wistar Rats from the Pepsin-Trypsin-Digested Gliadin (PTG)-Induced Enteropathy. Nutrients, 10(11), 1698. https://doi.org/10.3390/nu10111698