Associations of Brain Reactivity to Food Cues with Weight Loss, Protein Intake and Dietary Restraint during the PREVIEW Intervention


 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Measurements
2.3. Study Design
2.4. Brain Imaging Paradigm and Acquisition
2.5. Image Processing
2.6. Body Weight and Composition
2.7. Blood Samples and Urinary Nitrogen Excretion
2.8. Physical Activity
2.9. Eating Behaviour
2.10. Statistical Analysis
3. Results
3.1. No Differences in Brain Activity Between MP and HP Groups
3.2. Whole Brain Analysis-Relations Between Brain Activation and BMI, Body-Fat Percentage, HOMA-IR, Protein Intake, Activity or Eating Behaviour
3.2.1. Food > Non-Food Brain Response
3.2.2. High Calorie > Low Calorie Brain Response
3.3. ROI Analysis-Relations between Brain Activation and BMI, Fat Mass, HOMA-IR, Protein Intake, Activity or Eating Behaviour
3.3.1. Food > Non-Food Brain Response
3.3.2. High Calorie > Low-Calorie Brain Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vazquez, G.; Duval, S.; Jacobs, D.R., Jr.; Silventoinen, K. Comparison of body mass index, waist circumference and waist/hip ratio in predicting incident diabetes: A meta-analysis. Epidemiol. Rev. 2007, 29, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Eckel, R.H.; Kahn, S.E.; Ferrannini, E.; Goldfine, A.B.; Nathan, D.M.; Schwartz, M.W.; Smith, R.J.; Smith, S.R. Obesity and type 2 diabetes: What can be unified and what needs to be individualized? Diabetes Care 2011, 34, 1424–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stice, E.; Spoor, S.; Bohon, C.; Veldhuizen, M.G.; Small, D.M. Relation of reward from food intake and anticipated food intake to obesity: A functional magnetic resonance imaging study. J. Abnorm. Psychol. 2008, 117, 924–935. [Google Scholar] [CrossRef] [PubMed]
- Martens, M.J.; Born, J.M.; Lemmens, S.G.; Karhunen, L.; Heinecke, A.; Goebel, R.; Adam, T.C.; Westerterp-Plantenga, M.S. Increased sensitivity to food cues in the fasted state and decreased inhibitory control in the satiated state in the overweight. Am. J. Clin. Nutr. 2013, 97, 471–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chechlacz, M.; Rotshtein, P.; Klamer, S.; Porubska, K.; Higgs, S.; Booth, D.; Fritsche, A.; Preissl, H.; Abele, H.; Birbaumer, N.; et al. Diabetes dietary management alters responses to food pictures in brain regions associated with motivation and emotion: A functional magnetic resonance imaging study. Diabetologia 2009, 52, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Carnell, S.; Gibson, C.; Benson, L.; Ochner, C.N.; Geliebter, A. Neuroimaging and obesity: Current knowledge and future directions. Obes. Rev. 2012, 13, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Vidarsdottir, S.; Smeets, P.A.; Eichelsheim, D.L.; van Osch, M.J.; Viergever, M.A.; Romijn, J.A.; van der Grond, J.; Pijl, H. Glucose ingestion fails to inhibit hypothalamic neuronal activity in patients with type 2 diabetes. Diabetes 2007, 56, 2547–2550. [Google Scholar] [CrossRef] [PubMed]
- Kullmann, S.; Heni, M.; Veit, R.; Scheffler, K.; Machann, J.; Haring, H.U.; Fritsche, A.; Preissl, H. Selective insulin resistance in homeostatic and cognitive control brain areas in overweight and obese adults. Diabetes Care 2015, 38, 1044–1050. [Google Scholar] [CrossRef] [PubMed]
- Adam, T.C.; Tsao, S.; Page, K.A.; Hu, H.; Hasson, R.E.; Goran, M.I. Insulin sensitivity and brain reward activation in overweight Hispanic girls: A pilot study. Pediatr. Obes. 2015, 10, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Jastreboff, A.M.; Sinha, R.; Lacadie, C.; Small, D.M.; Sherwin, R.S.; Potenza, M.N. Neural correlates of stress- and food cue-induced food craving in obesity: Association with insulin levels. Diabetes Care 2013, 36, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Drummen, M.; Dorenbos, E.; Vreugdenhil, A.C.; Raben, A.; Westerterp-Plantenga, M.S.; Adam, T.C. Insulin resistance, weight and behavioural variables as determinants of brain reactivity to food cues—A preview study. Am. J. Clin. Nutr. 2018, in press. [Google Scholar]
- Figlewicz, D.P.; Evans, S.B.; Murphy, J.; Hoen, M.; Baskin, D.G. Expression of receptors for insulin and leptin in the ventral tegmental area/substantia nigra (VTA/SN) of the rat. Brain Res. 2003, 964, 107–115. [Google Scholar] [CrossRef]
- Behary, P.; Miras, A.D. Brain responses to food and weight loss. Exp. Physiol. 2014, 99, 1121–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, S.; Heinze, J.M.; Fritsche, A.; Linder, K.; von Feilitzsch, M.; Konigsrainer, A.; Häring, H.-U.; Veit, R.; Preissl, H. Neuronal Food Reward Activity in Patients with type 2 diabetes with improved glycaemic control after Bariatric Surgery. Diabetes Care 2016, 39, 1311–1317. [Google Scholar] [CrossRef] [PubMed]
- Journel, M.; Chaumontet, C.; Darcel, N.; Fromentin, G.; Tome, D. Brain responses to high-protein diets. Adv. Nutr. 2012, 3, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Min, D.K.; Tuor, U.I.; Koopmans, H.S.; Chelikani, P.K. Changes in differential functional magnetic resonance signals in the rodent brain elicited by mixed-nutrient or protein-enriched meals. Gastroenterology 2011, 141, 1832–1841. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; An, R.; Zhang, Y.; Li, X.; Wang, S. Correlations of macronutrient-induced functional magnetic resonance imaging signal changes in human brain and gut hormone responses. Am. J. Clin. Nutr. 2012, 96, 275–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leidy, H.J.; Ortinau, L.C.; Douglas, S.M.; Hoertel, H.A. Beneficial effects of a higher-protein breakfast on the appetitive, hormonal and neural signals controlling energy intake regulation in overweight/obese, “breakfast-skipping,” late-adolescent girls. Am. J. Clin. Nutr. 2013, 97, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Griffioen-Roose, S.; Smeets, P.A.; van den Heuvel, E.; Boesveldt, S.; Finlayson, G.; de Graaf, C. Human protein status modulates brain reward responses to food cues. Am. J. Clin. Nutr. 2014, 100, 113–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogelholm, M.; Larsen, T.M.; Westerterp-Plantenga, M.; Macdonald, I.; Martinez, J.A.; Boyadjieva, N.; Poppitt, S.; Schlicht, W.; Stratton, G.; Sundvall, J.; et al. PREVIEW: Prevention of Diabetes through Lifestyle Intervention and Population Studies in Europe and around the World. Design, Methods and Baseline Participant Description of an Adult Cohort Enrolled into a Three-Year Randomised Clinical Trial. Nutrients 2017, 9, 632. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.J. The emotion probe. Studies of motivation and attention. Am. Psychol. 1995, 50, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Plasqui, G.; Soenen, S.; Westerterp-Plantenga, M.S.; Westerterp, K.R. Measurement of longitudinal changes in body composition during weight loss and maintenance in overweight and obese subjects using air-displacement plethysmography in comparison with the deuterium dilution technique. Int. J. Obes. 2011, 35, 1124–1130. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Stunkard, A.J.; Messick, S. The three-factor eating questionnaire to measure dietary restraint, disinhibition and hunger. J. Psychosom. Res. 1985, 29, 71–83. [Google Scholar] [CrossRef]
- Institute of Medicine Committee on Military Nutrition Research. The Role of Protein and Amino Acids in Sustaining and Enhancing Performance; National Academies Press: Washington, DC, USA, 1999. [Google Scholar]
- Choi, S.; Disilvio, B.; Fernstrom, M.H.; Fernstrom, J.D. Meal ingestion, amino acids and brain neurotransmitters: Effects of dietary protein source on serotonin and catecholamine synthesis rates. Physiol. Behav. 2009, 98, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Growdon, J.H.; Cohen, E.L.; Wurtman, R.J. Effects of oral choline administration on serum and CSF choline levels in patients with Huntington’s disease. J. Neurochem. 1977, 28, 229–231. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, T.; Mallick, H.N.; Torii, K. Activation of the gut-brain axis by dietary glutamate and physiologic significance in energy homeostasis. Am. J. Clin. Nutr. 2009, 90, 832S–837S. [Google Scholar] [CrossRef] [PubMed]
- Uematsu, A.; Tsurugizawa, T.; Uneyama, H.; Torii, K. Brain-gut communication via vagus nerve modulates conditioned flavor preference. Eur. J. Neurosci. 2010, 31, 1136–1143. [Google Scholar] [CrossRef] [PubMed]
- Avena, N.M.; Rada, P.; Hoebel, B.G. Sugar and fat bingeing have notable differences in addictive-like behaviour. J. Nutr. 2009, 139, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.; Hare, T.; Rangel, A. Temporally dissociable mechanisms of self-control: Early attentional filtering versus late value modulation. J. Neurosci. 2013, 33, 18917–18931. [Google Scholar] [CrossRef] [PubMed]
- Neseliler, S.; Hu, W.; Larcher, K.; Zacchia, M.; Dadar, M.; Scala, S.G.; Lamarche, M.; Zeighami, Y.; Stotland, S.C.; Larocque, M.; et al. Neurocognitive and hormonal correlates of voluntary weight loss in humans. Cell Metab. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hare, T.A.; Malmaud, J.; Rangel, A. Focusing attention on the health aspects of foods changes value signals in vmPFC and improves dietary choice. J. Neurosci. 2011, 31, 11077–11087. [Google Scholar] [CrossRef] [PubMed]
- Deckersbach, T.; Das, S.K.; Urban, L.E.; Salinardi, T.; Batra, P.; Rodman, A.M.; Arulpragasam, A.R.; Dougherty, D.D.; Roberts, S.B. Pilot randomized trial demonstrating reversal of obesity-related abnormalities in reward system responsivity to food cues with a behavioural intervention. Nutr. Diabetes 2014, 4, e129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochner, C.N.; Laferrere, B.; Afifi, L.; Atalayer, D.; Geliebter, A.; Teixeira, J. Neural responsivity to food cues in fasted and fed states pre and post gastric bypass surgery. Neurosci. Res. 2012, 74, 138–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neill, B.V.; Bullmore, E.T.; Miller, S.; McHugh, S.; Simons, D.; Dodds, C.M.; Koch, A.; Napolitano, A.; Nathan, P.J. The relationship between fat mass, eating behaviour and obesity-related psychological traits in overweight and obese individuals. Appetite 2012, 59, 656–661. [Google Scholar]
- Murdaugh, D.L.; Cox, J.E.; Cook, E.W.; Weller, R.E. fMRI reactivity to high-calorie food pictures predicts short- and long-term outcome in a weight-loss program. NeuroImage 2012, 59, 2709–2721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Romero, A.; Adam, T.C.; Hu, H.H.; Monterosso, J.; Page, K.A. Abdominal fat is associated with a greater brain reward response to high-calorie food cues in Hispanic women. Obesity 2013, 21, 2029–2036. [Google Scholar] [CrossRef] [PubMed]
- Jastreboff, A.M.; Lacadie, C.; Seo, D.; Kubat, J.; Van Name, M.A.; Giannini, C.; Savoye, M.; Todd Constable, R.; Sherwin, R.S.; Caprio, S.; et al. Leptin is associated with exaggerated brain reward and emotion responses to food images in adolescent obesity. Diabetes Care 2014, 37, 3061–3068. [Google Scholar] [CrossRef] [PubMed]
- Grosshans, M.; Vollmert, C.; Vollstadt-Klein, S.; Tost, H.; Leber, S.; Bach, P.; Bu¨hler, M.; von der Goltz, C.; Mutschler, J.; Loeber, S.; et al. Association of leptin with food cue-induced activation in human reward pathways. Arch. Gen. Psychiatry 2012, 69, 529–537. [Google Scholar] [PubMed]
- Rosenbaum, M.; Sy, M.; Pavlovich, K.; Leibel, R.L.; Hirsch, J. Leptin reverses weight loss-induced changes in regional neural activity responses to visual food stimuli. J. Clin. Investig. 2008, 118, 2583–2591. [Google Scholar] [CrossRef] [PubMed]
- Cazettes, F.; Cohen, J.I.; Yau, P.L.; Talbot, H.; Convit, A. Obesity-mediated inflammation may damage the brain circuit that regulates food intake. Brain Res. 2011, 1373, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felger, J.C.; Treadway, M.T. Inflammation Effects on Motivation and Motor Activity: Role of Dopamine. Neuropsychopharmacology 2017, 42, 216–241. [Google Scholar] [CrossRef] [PubMed]
- Heni, M.; Kullmann, S.; Preissl, H.; Fritsche, A.; Haring, H.U. Impaired insulin action in the human brain: Causes and metabolic consequences. Nat. Rev. Endocrinol. 2015, 11, 701–711. [Google Scholar] [CrossRef] [PubMed]
- Tiedemann, L.J.; Schmid, S.M.; Hettel, J.; Giesen, K.; Francke, P.; Buchel, C.; Brassen, S. Central insulin modulates food valuation via mesolimbic pathways. Nat. Commun. 2017, 8, 16052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerterp-Plantenga, M.S.; Lejeune, M.P.; Nijs, I.; van Ooijen, M.; Kovacs, E.M. High protein intake sustains weight maintenance after body weight loss in humans. Int. J. Obes. Relat. Metab. Disord. 2004, 28, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Westerterp-Plantenga, M.S.; Kempen, K.P.; Saris, W.H. Determinants of weight maintenance in women after diet-induced weight reduction. Int. J. Obes. Relat. Metab. Disord. 1998, 22, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Rideout, C.A.; McLean, J.A.; Barr, S.I. Women with high scores for cognitive dietary restraint choose foods lower in fat and energy. J. Am. Dietetic Assoc. 2004, 104, 1154–1157. [Google Scholar] [CrossRef] [PubMed]
- Moreira, P.; de Almeida, M.D.; Sampaio, D. Cognitive restraint is associated with higher intake of vegetables in a sample of university students. Eat. Behav. 2005, 6, 229–237. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| MP (n = 12) | 2 Years | HP (n = 15) | 2 Years | Total (n = 27) | 2 Years | |
|---|---|---|---|---|---|---|
| Baseline | Baseline | Baseline | ||||
| Age | 54.7 (10.6) | 52.7 (10.3) | 53.6 (10.3) | |||
| Height | 1.74 (0.1) | 1.69 (0.12) | 1.71 (0.12) | |||
| Weight (kg) | 99.4 (17.4) | 90.6 (16.0) | 89.7 (15.4) | 81.9 (15.6) | 94.0 (16.7) | 85.8 (16.1) *** |
| BMI (kg/m2) | 32.7 (3.1) | 29.9 (3.5) | 31.2 (3.3) | 28.5 (4.1) | 31.8 (3.2) | 29.1 (3.8) *** |
| FM | 40.9 (8.8) | 33.9 (12.0) | 35.8 (8.7) | 29.7 (10.9) | 38.1 (9.0) | 31.6 (11.4) *** |
| Body-fat % | 41.4 (6.7) | 37.2 (10.1) | 40.2 (7.3) | 36.2 (9.9) | 40.7 (6.9) | 36.6 (9.8) *** |
| Insulin | 13.0 (6.4) | 10.4 (5.5) | 14.6 (6.5) | 10.4 (3.7) | 13.9 (6.4) | 10.4 (4.5) ** |
| Glucose | 6.4 (0.8) | 6.0 (0.7) | 6.3 (0.5) | 5.9 (0.4) | 6.4 (0.6) | 5.9 (0.5) ** |
| HOMA-IR | 3.7 (1.9) | 2.8 (1.8) | 4.2 (2.1) | 2.7 (1.1) | 4.0 (2.0) | 2.8 (1.4) * |
| CPM | 302.4 (116.1) | 262.2 (97.4) | 380.1 (109.3) | 338.5 (116.8) | 345.2 (116.3) | 304.2 (112.6) |
| TFEQ f1 | 7.7 (4.4) | 13.6 (2.8) | 8.9 (4.8) | 13.1 (3.5) | 8.3 (4.5) | 13.3 (3.1) *** |
| TFEQ f2 | 7.5 (3.3) | 6.4 (2.3) | 8.6 (3.7) | 7.5 (4.0) | 8.1 (3.5) | 7.0 (3.4) |
| TFEQ f3 | 5.7 (3.0) | 4.1 (3.1) | 5.9 (3.6) | 5.0 (3.6) | 5.8 (3.3) | 4.6 (3.3) * |
| Contrast | Variable | AAL | k | r | r * | p * | x | y | z | Cluster Extent |
|---|---|---|---|---|---|---|---|---|---|---|
| ΔF > nF | ΔBody-fat% | gyrus rectus L | 540 | −0.66 | −0.54 * | 0.005 * | −1 | 55 | −16 | 906 |
| thalamus L | 0.64 | 0.62 * | 0.001 * | −20 | −22 | 13 | 1010 | |||
| no label (white matter L) | −0.63 | −0.72 * | <0.001 * | −20 | −13 | 38 | 579 | |||
| middle frontal gyrus L | 0.63 | 0.65 * | <0.001 * | −24 | 23 | 41 | 1314 | |||
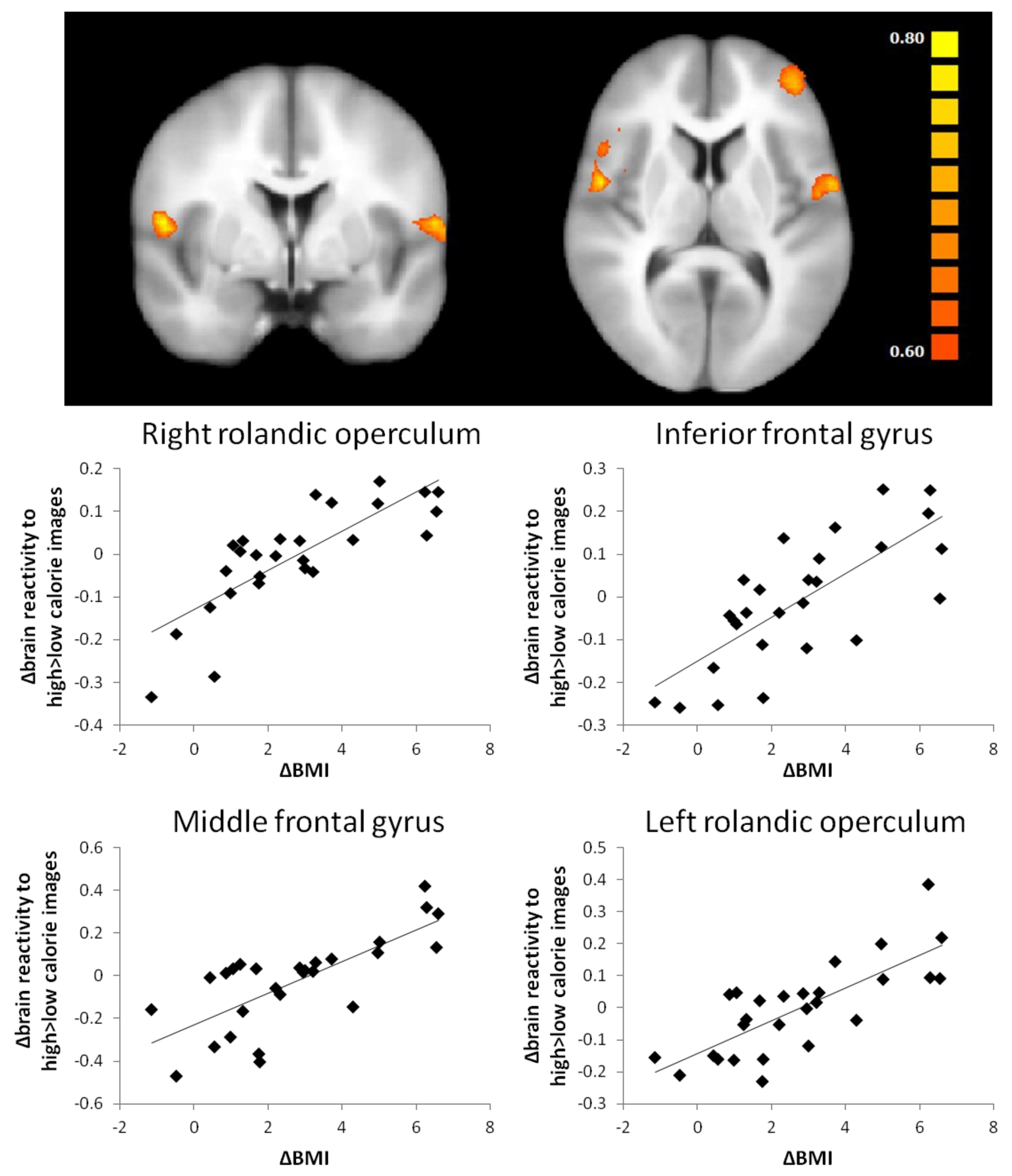
| ΔH > L | ΔBMI | rolandic operculum R | 621 | 0.66 | - | - | 55 | 0 | 9 | 688 |
| inferior frontal gyrus R | 0.63 | - | - | 47 | 17 | 3 | 1807 | |||
| middle frontal gyrus L | 0.66 | - | - | −41 | 50 | 8 | 1610 | |||
| rolandic operculum L | 0.65 | - | - | −58 | −2 | 8 | 796 | |||
| ΔBody-fat % | angular gyrus R | 675 | 0.65 | 0.71 * | <0.001 * | 61 | −51 | 34 | 2881 | |
| middle temporal gyrus R | 0.62 | 0.72 * | <0.001 * | 48 | 1 | −25 | 893 | |||
| putamen R | 0.65 | 0.68 * | <0.001 * | 31 | 6 | −1 | 11,852 | |||
| superior frontal gyrus L | 0.65 | 0.65 * | <0.001 * | −15 | 59 | 16 | 8828 | |||
| superior occipital gyrus R | 0.64 | 0.57 * | 0.003 * | 17 | −86 | 21 | 1269 | |||
| insula L | 0.65 | 0.71 * | <0.001 * | −40 | 14 | 2 | 8394 | |||
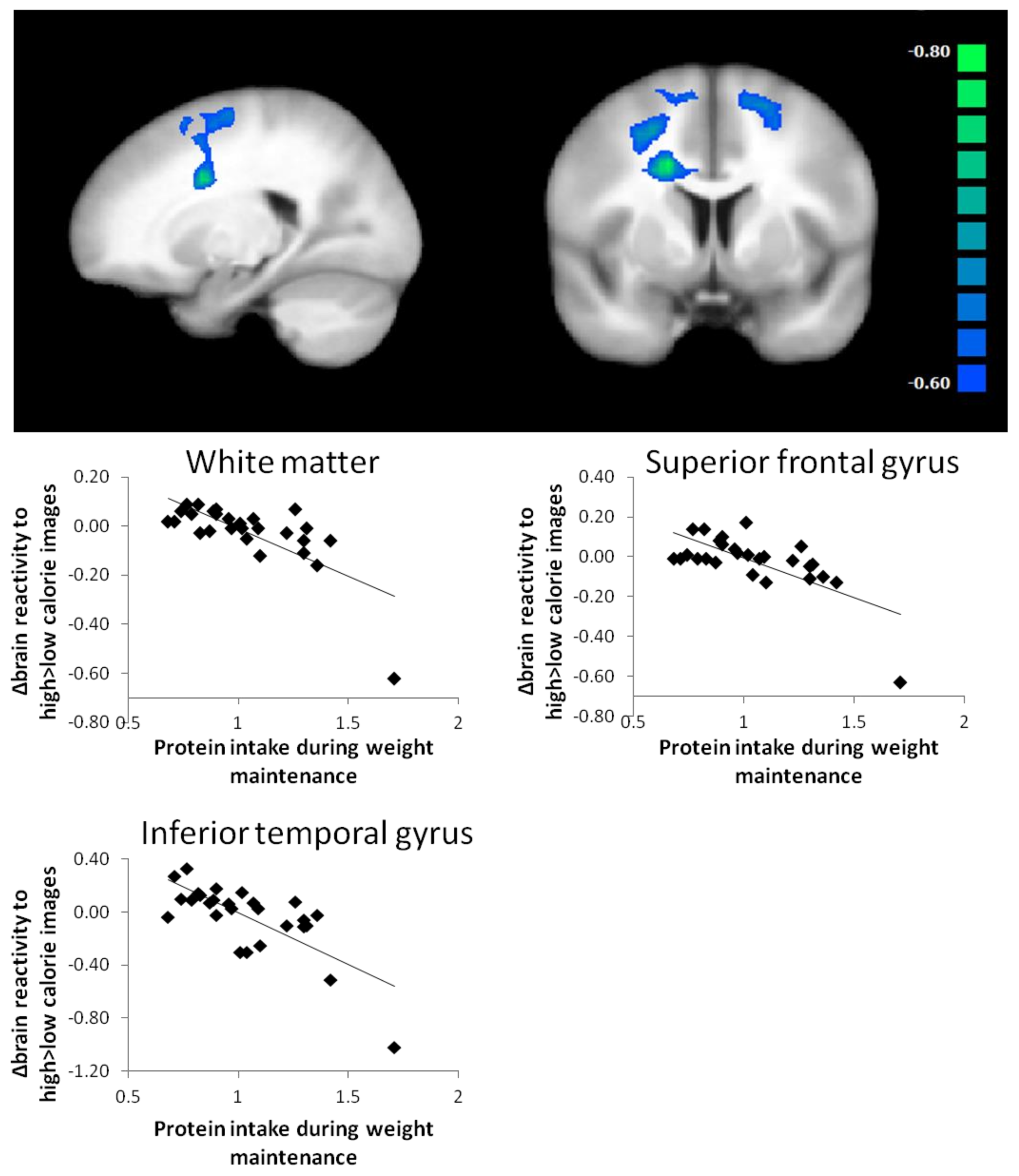
| WM protein intake (g/kg) | no label (white matter R) | 405 | −0.63 | −0.71 * | <0.001 * | 20 | 4 | 49 | 4040 | |
| superior frontal gyrus L | −0.64 | −0.67 * | <0.001 * | −16 | 9 | 59 | 2398 | |||
| inferior temporal gyrus L | −0.64 | −0.72 * | <0.001 * | −65 | −34 | −18 | 1062 | |||
| ΔTFEQ f1 | superior temporal gyrus R | 569 | −0.65 | −0.69 | <0.001 * | 59 | −44 | 15 | 1835 | |
| precentral gyrus R | −0.67 | −0.72 | <0.001 * | 51 | 6 | 38 | 1127 | |||
| superior occipital gyrus R | −0.64 | −0.64 | 0.001 * | 28 | −79 | 23 | 601 | |||
| superior occipital gyrus L | −0.66 | −0.80 | <0.001 * | −21 | −79 | 33 | 1014 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drummen, M.; Dorenbos, E.; Vreugdenhil, A.C.E.; Stratton, G.; Raben, A.; Westerterp-Plantenga, M.S.; Adam, T.C. Associations of Brain Reactivity to Food Cues with Weight Loss, Protein Intake and Dietary Restraint during the PREVIEW Intervention. Nutrients 2018, 10, 1771. https://doi.org/10.3390/nu10111771
Drummen M, Dorenbos E, Vreugdenhil ACE, Stratton G, Raben A, Westerterp-Plantenga MS, Adam TC. Associations of Brain Reactivity to Food Cues with Weight Loss, Protein Intake and Dietary Restraint during the PREVIEW Intervention. Nutrients. 2018; 10(11):1771. https://doi.org/10.3390/nu10111771
Chicago/Turabian StyleDrummen, Mathijs, Elke Dorenbos, Anita C. E. Vreugdenhil, Gareth Stratton, Anne Raben, Margriet S. Westerterp-Plantenga, and Tanja C. Adam. 2018. "Associations of Brain Reactivity to Food Cues with Weight Loss, Protein Intake and Dietary Restraint during the PREVIEW Intervention" Nutrients 10, no. 11: 1771. https://doi.org/10.3390/nu10111771
APA StyleDrummen, M., Dorenbos, E., Vreugdenhil, A. C. E., Stratton, G., Raben, A., Westerterp-Plantenga, M. S., & Adam, T. C. (2018). Associations of Brain Reactivity to Food Cues with Weight Loss, Protein Intake and Dietary Restraint during the PREVIEW Intervention. Nutrients, 10(11), 1771. https://doi.org/10.3390/nu10111771