Comparison of the Allergenicity and Immunogenicity of Camel and Cow’s Milk—A Study in Brown Norway Rats

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Products
2.2. In Silico Protein Analyses
2.3. Denaturation of Products
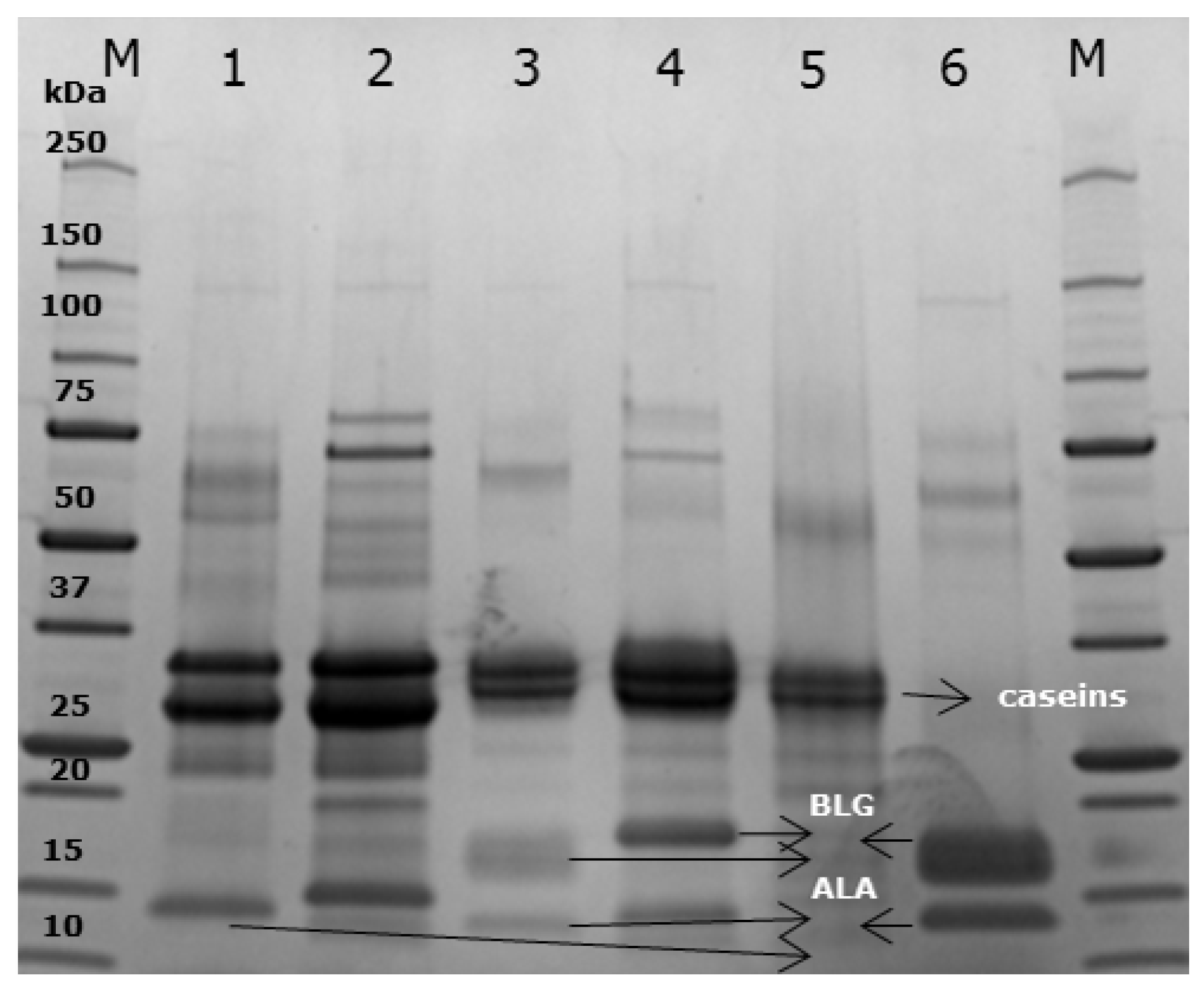
2.4. SDS-PAGE Electrophoresis
2.5. Animals
2.6. Animal Sensitisation Studies
2.7. Ear Swelling Test
2.8. Indirect ELISA for Specific IgG1 Detection
2.9. Antibody Capture ELISA to Detect Specific IgE
2.10. Avidity Measurements
2.11. Inhibitory ELISA
2.12. Immunoblotting
2.13. Statistical Analysis of Data
3. Results
3.1. Protein Characterisation
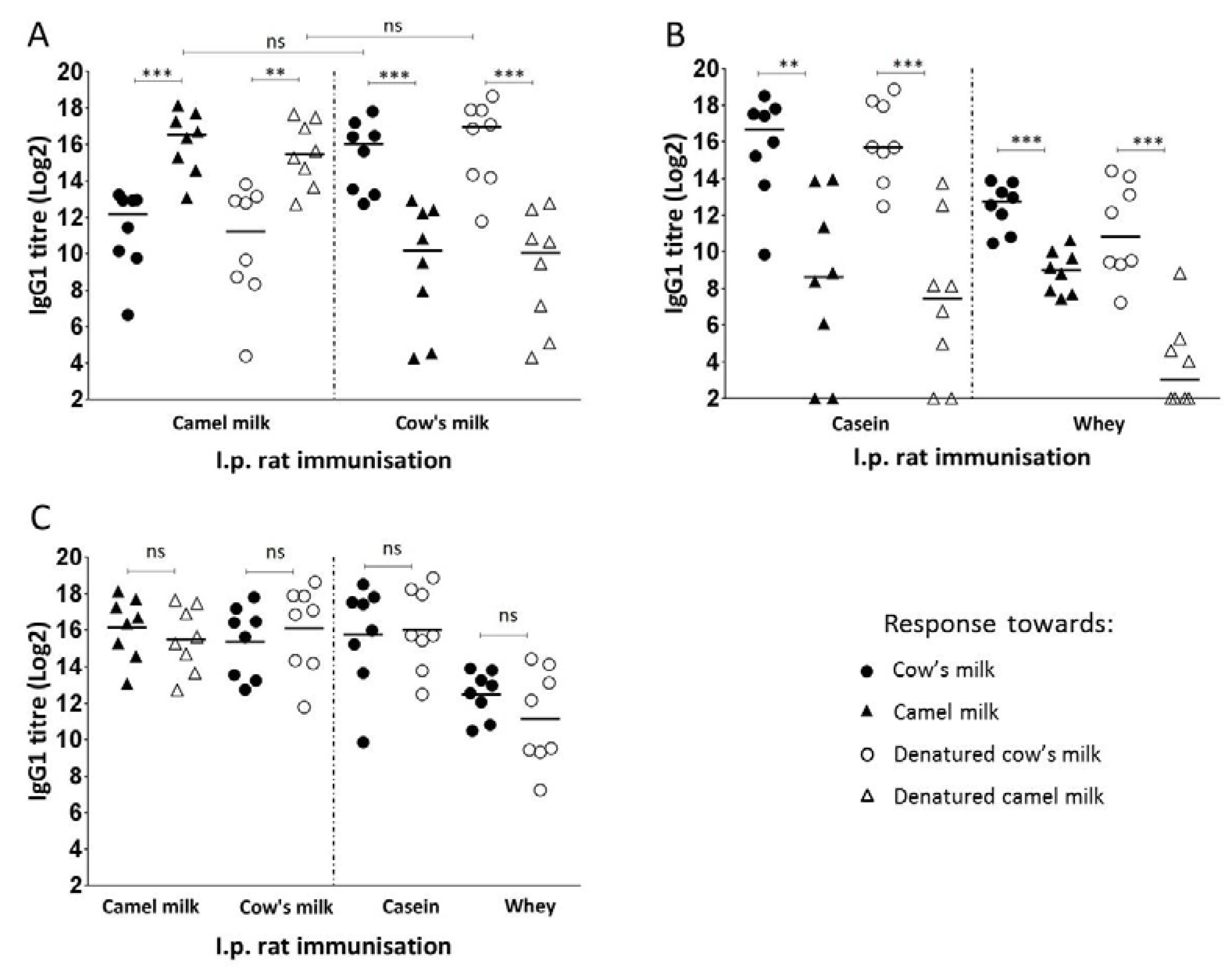
3.2. Camel and Cow’s Milk Immunogenicity and Cross-Reactivity
3.3. Linear and Conformational Epitope Recognition
3.4. Inhibitory ELISA
3.4.1. IgG1 Antibody Competition of Native Camel and Cow’s Milk
3.4.2. IgG1 Antibody Competition Towards Denatured Camel and Cow’s Milk
3.5. Specific IgG1 Antibody Avidity
3.6. Sensitising Capacity of Camel and Cow’s Milk
3.7. Eliciting Capacity of Camel and Cow’s Milk
3.8. Immunoblot
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALA | α-lactalbumin |
| BLG | β-lactoglobulin |
| BN | Brown Norway |
| CMA | Cow’s milk allergy |
| DIG | dioxigenin |
| ELISA | Enzyme-linked immunosorbent assay |
| EU | endotoxin units |
| HRP | horse-radish peroxidase |
| IC50 | half minimum inhibitory concentration |
| i.p. | intraperitoneally |
| kDa | kilodalton |
| KSCN | potassium thiocyanate |
| LF | actoferrin |
| LP | lactoperoxidase |
| MW | molecular weight |
| NA | not available |
| OD | optical density |
| OVA | ovalbumin |
| PBS | phosphate buffered saline |
| PBT-T | phosphate buffered saline-tween |
| PAGE | polyacrylamide gel electrophoresis |
| PVDF | polyvinylidene difluoride |
| RT | room temperature |
| SA | serum albumin |
| SD | standard deviation |
| SDS | sodium dodecyl sulphate |
| TMB | 3,3′,5,5′-tetramethylbenzidine |
| Tris | 2-amino-2-(hydroxymethyl)-1,3-propenediol |
| WHO | World Health Organisation |
References
- Hochwallner, H.; Schulmeister, U.; Swoboda, I.; Spitzauer, S.; Valenta, R. Cow’s milk allergy: From allergens to new forms of diagnosis, therapy and prevention. Methods 2014, 66, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Bellioni-Businco, B.; Paganelli, R.; Lucenti, P.; Giampietro, P.G.; Perborn, H.; Businco, L. Allergenicity of goat’s milk in children with cow’s milk allergy. J. Allergy Clin. Immunol. 1999, 103, 1191–1194. [Google Scholar] [CrossRef]
- Carrard, A.; Rizzuti, D.; Sokollik, C. Update on food allergy. Eur. J. Allergy Clin. Immunol. 2015, 70, 1511–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiocchi, A.; Brozek, J.; Schünemann, H. World Allergy Organization (WAO) Diagnosis and Rationale for Action against Cow’s Milk Allergy (DRACMA) Guidelines. Pediatric. Allergy Immunol. 2010, 21, 1–125. [Google Scholar] [CrossRef]
- El-Agamy, E.I. The challenge of cow milk protein allergy. Small Rumin. Res. 2007, 68, 64–72. [Google Scholar] [CrossRef]
- Many Kids May Not Outgrow Cow’s Milk Allergy. Reuters. Available online: https://www.reuters.com/article/us-milk-allergy/many-kids-may-not-outrow-cows-milk-allergy-idUSCOL87231020071218 (accessed on 25 November 2018).
- Milk and Dairy Allergy. ACAAI. Available online: https://acaai.org/allergies/types-allergies/food-allergy/types-food-allergy/milk-dairy-allergy (accessed on 25 November 2018).
- El-Agamy, E.I.; Nawar, M.; Shamsia, S.M.; Awad, S.; Haenlein, G.F.W. Are camel milk proteins convenient to the nutrition of cow milk allergic children? Small Rumin. Res. 2009, 82, 1–6. [Google Scholar] [CrossRef]
- Von Berg, A.; Koletzko, S.; Grübl, A.; Filipiak-Pittroff, B.; Wichmann, H.E.; Bauer, C.P.; Reinhardt, D.; Berdel, D.; German Infant Nutritional Intervention Study Group. The effect of hydrolyzed cow’s milk formula for allergy prevention in the first year of life: The German Infant Nutritional Intervention Study, a randomized double-blind trial. J. Allergy Clin. Immunol. 2003, 111, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Gateway to Dairy Production and Products. Food Agric. Organ. United Nations. Available online: http://www.fao.org/dairy-production-products/production/dairy-animals/en/ (accessed on 1 January 2018).
- Järvinen, K.-M.; Chatchatee, P.; Bardina, L.; Beyer, K.; Sampson, H.A. IgE and IgG binding epitopes on α-lactalbumin and β-lactoglobulin in cow’s milk allergy. Int. Arch. Allergy Immunol. 2001, 126, 111–118. [Google Scholar] [CrossRef]
- Dean, T.P.; Adler, B.R.; Ruge, F. In vitro allergenicity of cow’s milk substitutes. Clin. Exp. Allergy 1993, 23, 205–210. [Google Scholar] [CrossRef]
- Restani, P.; Gaiaschi, A.; Plebani, A.; Beretta, B.; Cavagni, G.; Fiocchi, A.; Poiesi, C.; Velona, A.; Ugazio, G.; Galli, C.L. Cross-reactivity between milk proteins from different animal species. Clin. Exp. Allergy 1999, 29, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Ehlayel, M.; Bener, A.; Abu Hazeima, K.; Al-Mesaifri, F. Camel Milk Is a Safer Choice than Goat Milk for Feeding Children with Cow Milk Allergy. ISRN Allergy 2011, 2011, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Spuergin, P.; Walter, M.; Schiltz, E.; Deichmann, K.; Forster, J.; Mueller, H. Allergenicity of α-caseins from cow, sheep, and goat. Allergy 1997, 52, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Fiocchi, A.; Dahda, L.; Dupont, C.; Campoy, C.; Fierro, V.; Nieto, A. Cow’s milk allergy: Towards an update of DRACMA guidelines. World Allergy Organ. J. 2016, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lonnerdal, B. Nutritional aspects of soy formula. Acta Pediatr Suppl. 1994, 83, 105–108. [Google Scholar] [CrossRef]
- Hinz, K.; O’Connor, P.M.; Huppertz, T.; Ross, R.P.; Kelly, A.L. Comparison of the principal proteins in bovine, caprine, buffalo, equine and camel milk. J. Dairy Res. 2012, 79, 185–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hailu, Y.; Hansen, E.B.; Seifu, E.; Eshetu, M.; Ipsen, R.; Kappeler, S. Functional and technological properties of camel milk proteins: A review. J. Dairy Res. 2016, 83, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Navarrete-Rodríguez, E.M.; Ríos-Villalobos, L.A.; Alcocer-Arreguín, C.R.; Del-Rio-Navarro, B.E.; Del Rio-Chivardi, J.M.; Saucedo-Ramírez, O.J.; Sienra-Monge, J.J.L.; Frias, R.V. Cross-over clinical trial for evaluating the safety of camel’s milk intake in patients who are allergic to cow’s milk protein. Allergol. Immunopathol. 2017, 4, 149–154. [Google Scholar] [CrossRef]
- Nagy, P.; Thomas, S.; Marko, O.; Juhasz, J. Milk production, raw milk quality and fertility of Dromedary camels (Camelus Dromedarius) under intensive management. Acta Weterinaria Hungarica. 2013, 61, 71–84. [Google Scholar] [CrossRef]
- Spencer, P.B.S.; Woolnough, A.P. Assessment and genetic characterisation of Australian camels using microsatellite polymorphisms. Live Stock Sci. 2010, 129, 241–245. [Google Scholar] [CrossRef]
- Nutrition. Available online: https://desertfarms.co.uk/pages/nutrition (accessed on 23 November 2018).
- Fresh and Frozen Camel Milk. Available online: https://www.oasismilk.com/en/camel-milk/ (accessed on 23 November 2018).
- Wolkaro, T.; Yusuf Kurtu, M.; Eshetu, M.; Ketema, M. Analysis of camel milk value chain in the pastoral areas of eastern Ethiopia. J. Camelid Sci. 2017, 10, 1–16. [Google Scholar]
- Sawaya, W.N.; Khalil, J.K.; Al-Shalhat, A.; Al-Mohammad, H. Chemical composition and nutritional quality of camel milk. J. Food Sci. 1984, 49, 744–747. [Google Scholar] [CrossRef]
- Madsen, J.; Kroghsbo, S.; Madsen, C.; Pozdnyakova, I.; Barkholt, V.; Bøgh, K.L. The impact of structural integrity and route of administration on the antibody specificity against three cow’s milk allergens—A study in Brown Norway rats. Clin. Transl. Allergy 2014, 4, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bøgh, K.L.; Barkholt, V.; Madsen, C.B. The sensitising capacity of intact β-lactoglobulin is reduced by Co-administration with digested β-lactoglobulin. Int. Arch. Allergy Immunol. 2013, 161, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Bertino, E.; Gastaldi, D.; Monti, G.; Baro, C.; Fortunato, D.; Garoffo, L.P. Detailed proteomic analysis on DM: Insight into its hypoallergenicity. Front. Biosci. 2010, 2, 526–536. [Google Scholar] [CrossRef]
- Rijnkels, M.; Elnitski, L.; Miller, W.; Rosen, J.M. Multispecies comparative analysis of a mammalian-specific genomic domain encoding secretory proteins. Genomics 2003, 82, 417–432. [Google Scholar] [CrossRef]
- Omar, A.; Harbourne, N.; Oruna-Concha, M.J. Quantification of major camel milk proteins by capillary electrophoresis. Int. Dairy J. 2016, 58, 31–35. [Google Scholar] [CrossRef]
- Broekman, H.C.H.; Eiwegger, T.; Upton, J.; Bøgh, K.L. IgE–the main player of food allergy. Drug Discov. Today Dis. Model. 2015, 17–18, 37–44. [Google Scholar] [CrossRef]
- Levieux, D.; Levieux, A.; El-Hatmi, H.; Rigaudière, J.P. Immunochemical quantification of heat denaturation of camel (Camelus dromedarius) whey proteins. J. Dairy Res. 2006, 73, 1–9. [Google Scholar] [CrossRef]
- Youcef, N.; Saidi, D.; Mezemaze, F.; Kaddouri, H.; Negaoui, H.; Chekroun, A. Cross Reactivity between Dromedary Whey Proteins and IgG Anti Bovine α-Lactalbumin and Anti Bovine β-Lactoglobulin. Am. J. Appl. Sci. 2009, 6, 1448–1452. [Google Scholar] [CrossRef]
- O’Mahony, J.A.; Fox, P.F. Milk Proteins: Introduction and Historical Aspects. Adv. Dairy Chem. 2013, 1, 43–85. [Google Scholar] [CrossRef]
- McMahon, D.J.; Oommen, B.S. Supramolecular Structure of the Casein Micelle. J. Dairy Sci. Elsevier 2008, 91, 1709–1721. [Google Scholar] [CrossRef] [PubMed]
- Beg, O.U.; Bahr-Lindstrom, H.; Zaidi, Z.H.; Jornvall, H. The primary structure of α-lactalbumin from camel milk. Eur. J. Biochem. 1985, 147, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Shabo, Y.; Barzel, R.; Margoulis, M.; Yagil, R. Camel milk for food allergies in children. Isr. Med. Assoc. J. 2005, 7, 796–798. [Google Scholar] [PubMed]
- Restani, P.; Beretta, B.; Fiocchi, A.; Ballabio, C.; Galli, C.L. Cross-reactivity between mammalian proteins. Ann. Allergy Asthma Immunol. 2002, 89, 11–15. [Google Scholar] [CrossRef]
- Wolpe, L.Z.; Silverstone, P.C. A series of substitutes for milk in the treatment of allergies. J. Pediatr. 1942, 21, 635–658. [Google Scholar] [CrossRef]
- Järvinen, K.M.; Chatchatee, P. Mammalian milk allergy: Clinical suspicion, cross-reactivities and diagnosis. Curr. Opin. Allergy Clin. Immunol. 2009, 9, 251–258. [Google Scholar] [CrossRef]
- Natale, M.; Bisson, C.; Monti, G.; Peltran, A.; Garoffo, L.P.; Valentini, S.; Fabris, C.; Bertino, E.; Coscia, A.; Conti, A. Cow’s milk allergens identification by two-dimensional immunoblotting and mass spectrometry. Mol. Nutr. Food Res. 2004, 48, 363–369. [Google Scholar] [CrossRef]
- Kroghsbo, S.; Bøgh, K.L.; Rigby, N.M.; Mills, E.N.C.; Rogers, A.; Madsen, C.B. Sensitization with 7S globulins from peanut, hazelnut, soy or pea induces ige with different biological activities which are modified by soy tolerance. Int. Arch. Allergy Immunol. 2011, 155, 212–224. [Google Scholar] [CrossRef]
- Aalberse, R.C. Structural biology of allergens. J. Allergy Clin. Immunol. 2000, 106, 228–238. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Goat | Sheep | Camel | Human (c) | ||
|---|---|---|---|---|---|
| Casein | β-casein | 91 | 91 | 67 | 55 (60) |
| αs1-casein | 88 | 88 | 47 | 33 (40) | |
| αs2-casein | 88 | 89 | 56 | NA (a) | |
| κ-casein | 85 | 85 | 58 | 52 (60) | |
| Whey | α-lactalbumin | 95 | 95 | 60 | 74 (62) |
| β-lactoglobulin | 93 | 93 | NA (b) | NA (b) | |
| serum albumin | 88 | 92 | 81 | 76 (76) | |
| lactoferrin | 92 | 92 | 75 | 70 (74) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maryniak, N.Z.; Hansen, E.B.; Ballegaard, A.-S.R.; Sancho, A.I.; Bøgh, K.L. Comparison of the Allergenicity and Immunogenicity of Camel and Cow’s Milk—A Study in Brown Norway Rats. Nutrients 2018, 10, 1903. https://doi.org/10.3390/nu10121903
Maryniak NZ, Hansen EB, Ballegaard A-SR, Sancho AI, Bøgh KL. Comparison of the Allergenicity and Immunogenicity of Camel and Cow’s Milk—A Study in Brown Norway Rats. Nutrients. 2018; 10(12):1903. https://doi.org/10.3390/nu10121903
Chicago/Turabian StyleMaryniak, Natalia Zofia, Egon Bech Hansen, Anne-Sofie Ravn Ballegaard, Ana Isabel Sancho, and Katrine Lindholm Bøgh. 2018. "Comparison of the Allergenicity and Immunogenicity of Camel and Cow’s Milk—A Study in Brown Norway Rats" Nutrients 10, no. 12: 1903. https://doi.org/10.3390/nu10121903
APA StyleMaryniak, N. Z., Hansen, E. B., Ballegaard, A. -S. R., Sancho, A. I., & Bøgh, K. L. (2018). Comparison of the Allergenicity and Immunogenicity of Camel and Cow’s Milk—A Study in Brown Norway Rats. Nutrients, 10(12), 1903. https://doi.org/10.3390/nu10121903