Effect of Wheat Bran on Fecal Butyrate-Producing Bacteria and Wheat Bran Combined with Barley on Bacteroides Abundance in Japanese Healthy Adults

Abstract
:1. Introduction
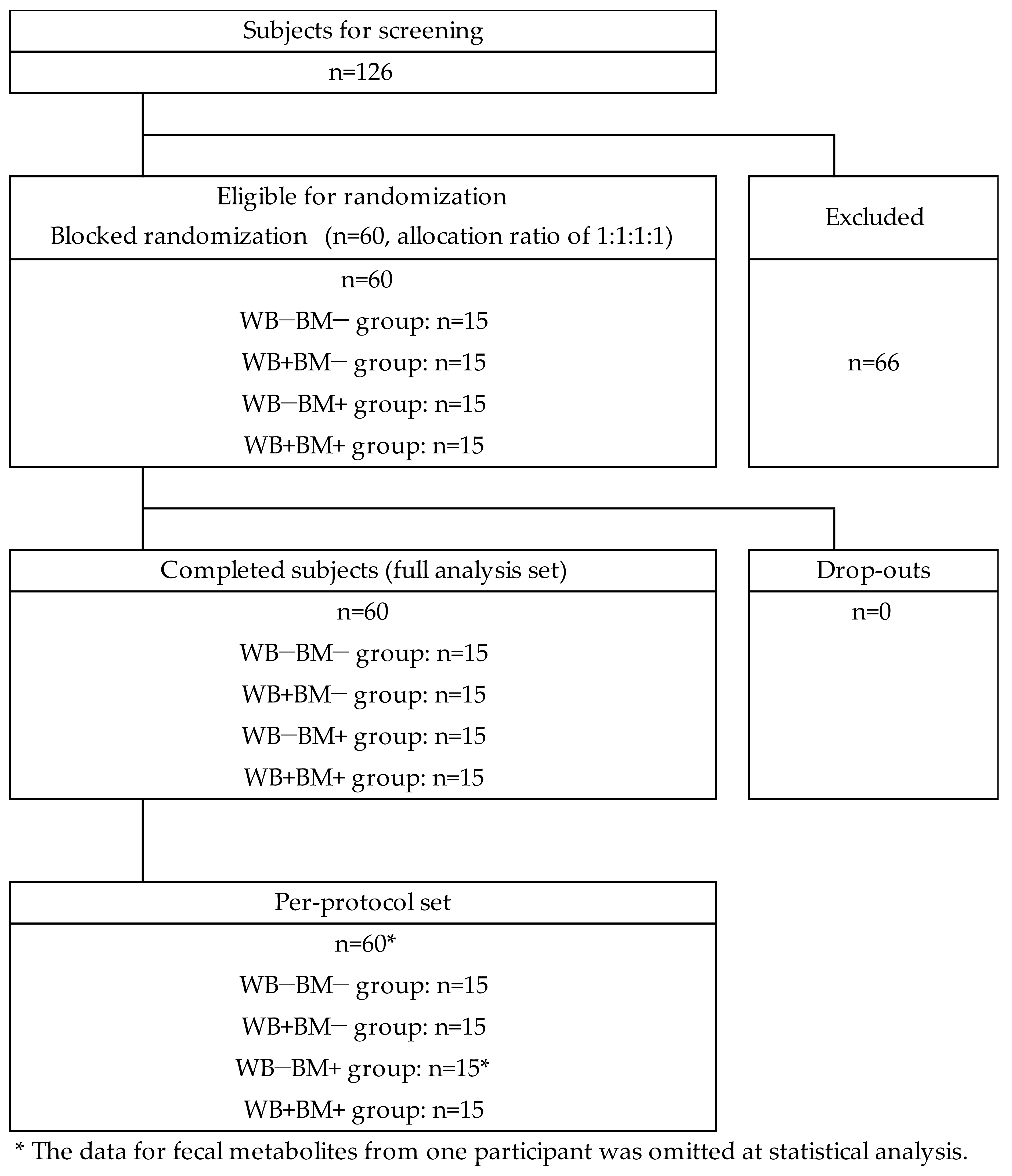
2. Materials and Methods
2.1. Study Design
2.2. Subjects
- Regular use of a medicine for intestinal disorders or an aperient (including a laxative).
- Regular consumption of health foods thought to improve constipation at the time of the screening examination.
- Use of any drugs that could affect digestion and nutrient absorption including antibiotics at the time of the screening examination.
- Inability or unwillingness to stop consuming probiotic or prebiotic supplements such as lactic acid bacteria, Bifidobacterium, Bacillus subtilis var. natto, food fortified with oligosaccharides, dietary fiber, large quantities of sugar or alcohol, barley-rich food, or other dietary health supplements for the entire duration of the study.
- Having a food allergy.
- Having a disease urgently needing treatment or having serious complications.
- Having a history of a digestive organ disease or an operation that could affect digestion, nutrient absorption, or bowel movements.
- Judged from the answers to the subject background questionnaire to be inappropriate as a subject.
- Judged from the results of a blood test during the screening examination to be inappropriate as a subject.
- Currently pregnant or intended to become pregnant during the study period.
- Currently nursing an infant.
- Having any drug dependency.
- Having an anamnesis or medical history of alcohol dependence.
- Participating in another study involving the intake of food, drugs, or cosmetics, or judged by one of the examining doctors involved in the trial to be inappropriate as a subject, e.g., subjects who plan to change their lifestyle during the test period or who would not obey the rules of the trial.
2.3. Dietary Supplementation
2.4. Food Frequency Questionnaire
2.5. Fecal Collection
2.6. Anthropometrical Measures and Biochemical Analysis
2.7. DNA Extraction and Analysis of Intestinal Microbiota Composition by High-Throughput Sequencing
2.8. Fecal Organic Acids, Indoles, Phenols, and Ammonia
2.9. Statistical Analysis
3. Results
3.1. Dietary Intake and Adherence
3.2. Anthropometrical Measures and Biochemical Analysis
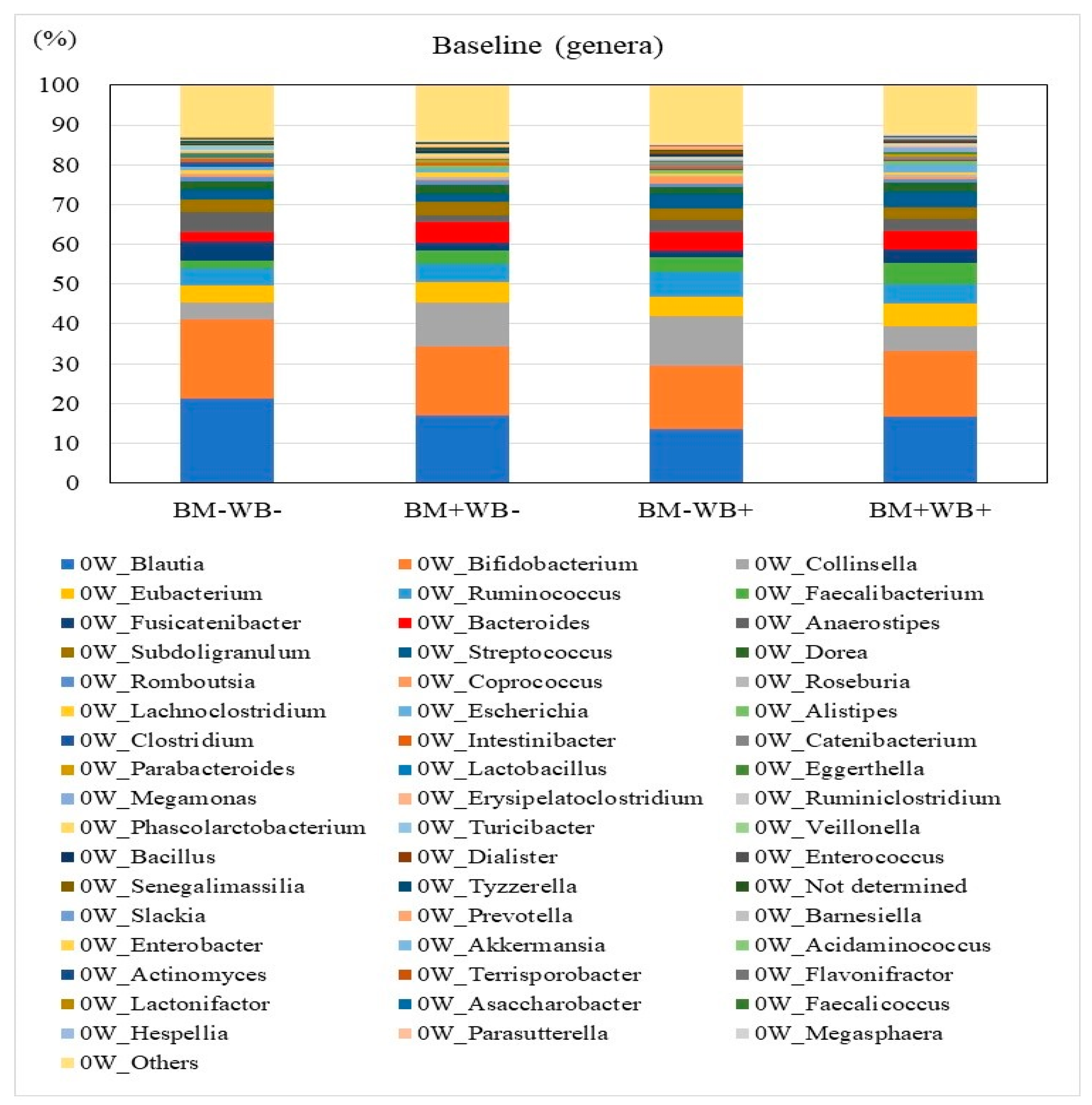
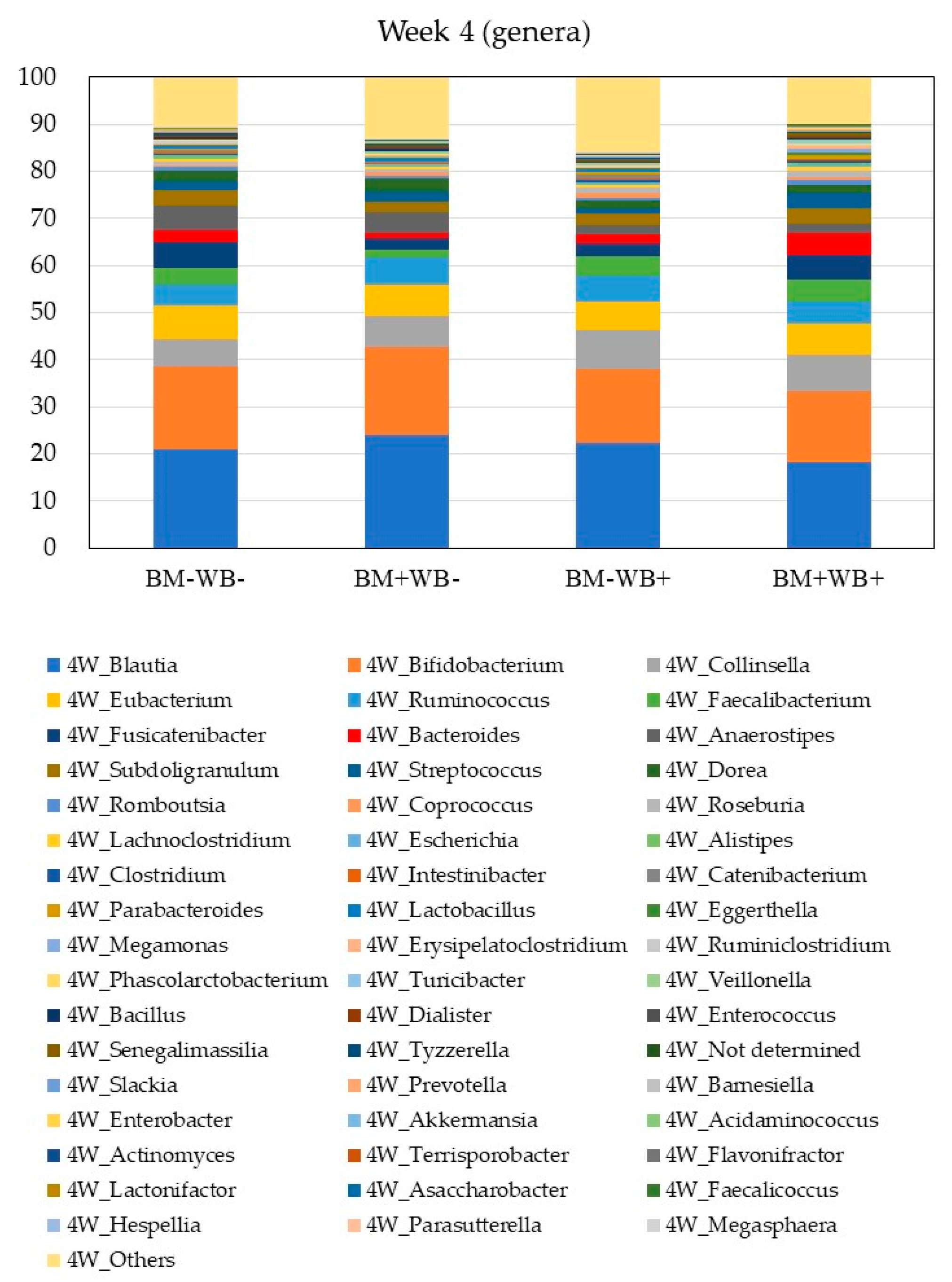
3.3. Fecal Microbiota Composition
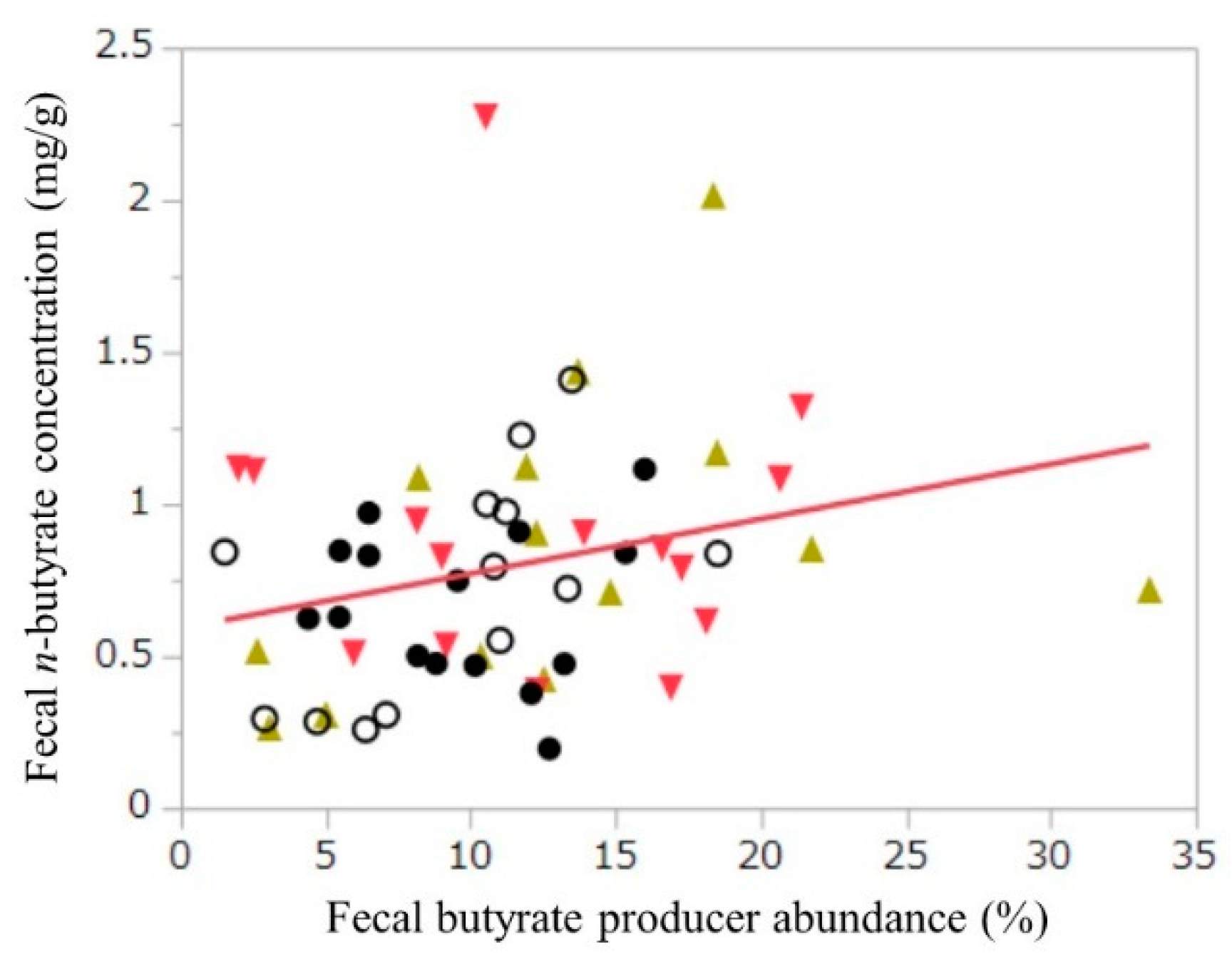
3.4. Fecal Organic Acids and Putrefaction Products
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ma, Y.; Hu, M.; Zhou, L.; Ling, S.; Li, Y.; Kong, B.; Huang, P. Dietary fiber intake and risks of proximal and distal colon cancers: A meta-analysis. Medicine 2018, 97, e11678. [Google Scholar] [CrossRef] [PubMed]
- Kritchevsky, D. Epidemiology of fibre, resistant starch and colorectal cancer. Eur. J. Cancer Prev. 1995, 4, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Giovannucci, E.; Willett, W.C. Dietary factors and risk of colon cancer. Ann. Med. 1994, 26, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.G. The benefits of breakfast cereal consumption: A systematic review of the evidence base. Adv. Nutr. 2014, 5, 636S–673S. [Google Scholar] [CrossRef]
- Sonnenburg, E.D.; Smits, S.A.; Tikhonov, M.; Higginbottom, S.K.; Wingreen, N.S.; Sonnenburg, J.L. Diet-induced extinctions in the gut microbiota compound over generations. Nature 2016, 529, 212–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, C.; Round, J.L. Defining dysbiosis and its influence on host immunity and disease. Cell. Microbiol. 2014, 16, 1024–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buttó, L.F.; Schaubeck, M.; Haller, D. Mechanisms of microbe–host interaction in Crohn’s disease: Dysbiosis vs. pathobiont selection. Front. Immunol. 2015, 6, 555. [Google Scholar] [CrossRef]
- Stevenson, L.; Phillips, F.; O’Sullivan, K.; Walton, J. Wheat bran: Its composition and benefits to health, a European perspective. Int. J. Food Sci. Nutr. 2012, 63, 1001–1013. [Google Scholar] [CrossRef]
- Rosa-Sibakova, N.; Poutanen, K.; Micard, V. How does wheat grain, bran and aleurone structure impact their nutritional and technological properties? Trends Food Sci. Technol. 2015, 41, 118–134. [Google Scholar] [CrossRef]
- Bijkerk, C.J.; Muris, J.W.M.; Knottnerus, J.A.; Hoes, A.W.; de Wit, N.J. Systematic review: The role of different types of fibre in the treatment of irritable bowel syndrome. Aliment. Pharmacol. Ther. 2004, 19, 245–251. [Google Scholar] [CrossRef]
- Vuksan, V.; Jenkins, A.L.; Jenkins, D.J.; Rogovik, A.L.; Sievenpiper, J.L.; Jovanovski, E. Using cereal to increase dietary fiber intake to the recommended level and the effect of fiber on bowel function in healthy persons consuming North American diets. Am. J. Clin. Nutr. 2008, 88, 1256–1262. [Google Scholar] [PubMed]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.J. The role of butyrate on colonic function. Aliment. Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, W.; Degroote, J.; Possemiers, S.; Chen, D.; De Smet, S.; Michiels, J. Arabinoxylan in wheat is more responsible than cellulose for promoting intestinal barrier function in weaned male piglets. J. Nutr. 2015, 145, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Russell, W.R.; Quartieri, A.; Rossi, M.; Parkhill, J.; Walker, A.W.; Flint, H.J. Wheat bran promotes enrichment within the human colonic microbiota of butyrate-producing bacteria that release ferulic acid. Environ. Microbiol. 2016, 18, 2214–2225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, S.A.; Shewry, P.R.; Gibson, G.R.; McCleary, B.V.; Rastall, R.A. In vitro fermentation of oat and barley derived β-glucans by human faecal microbiota. FEMS Microbiol. Ecol. 2008, 64, 482–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, A.; Kitazono, E.; Imose, K.; Urita, S.; Matsui, T. Effect of functional barley BARLEYmax (Tantangara) on intestinal regulation: A double-blind, randomized, placebo-controlled parallel group comparison clinical study. Jpn. Pharmacol. Ther. 2017, 45, 1047–1055. [Google Scholar]
- Henningsson, A.M.; Björck, I.M.; Nyman, E.M. Combinations of indigestible carbohydrates affect short-chain fatty acid formation in the hindgut of rats. J. Nutr. 2002, 132, 3098–3104. [Google Scholar] [CrossRef] [PubMed]
- Muir, J.G.; Yeow, E.G.W.; Keogh, J.; Pizzey, C.; Bird, A.R.; Sharpe, K.; O’Dea, K.; Macrae, F.A. Combining wheat bran with resistant starch has more beneficial effects on fecal indexes than does wheat bran alone. Am. J. Clin. Nutr. 2004, 79, 1020–1028. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Kondo, S.; Odamaki, T.; Miyaji, K.; Yaeshima, T.; Iwatsuki, K.; Togashi, H.; Benno, Y. Effect of yogurt containing Bifidobacterium longum BB536 on the defecation frequency and fecal characteristics of healthy adults: A double-blind cross over study. Jpn. J. Lact. Acid Bact. 2007, 18, 31–36. [Google Scholar] [CrossRef]
- Lee, S.C.; Rodriguez, F.; Storey, M.; Farmakalidis, E.; Prosky, L. Determination of soluble and insoluble dietary fiber in psyllium containing cereal products. J. AOAC Int. 1995, 78, 724–729. [Google Scholar]
- McCleary, B.V.; Codd, R. Measurement of 1-3, 1-4-β-d-glucan in barley and oats: A streamlined enzymic procedure. J. Sci. Food Agric. 1991, 55, 303–312. [Google Scholar] [CrossRef]
- McCleary, B.V.; Monaghan, D.A. Measurement of resistant starch. J. AOAC Int. 2002, 85, 665–675. [Google Scholar]
- Cyran, M.R.; Dynkowska, W.M. Mode of endosperm and wholemeal arabinoxylans solubilisation during rye breadmaking: Genotypic diversity in the level, substitution degree and macromolecular characteristics. Food Chem. 2014, 145, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Standard Tables of Food Composition in Japan, Seventh Revised Version (2015). Available online: http://www.mext.go.jp/en/policy/science_technology/policy/title01/detail01/sdetail01/sdetail01/1385122.htm (accessed on 28 October 2017).
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of Bacteria and Archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef] [PubMed]
- Erik, A. EA-Utils ”Command-Line Tools for Processing Biological Sequencing Data”. Available online: http://code.google.com/p/ea-utils (accessed on 30 September 2018).
- Available online: http://hannonlab.cshl.edu/fastx_toolkit/ (accessed on 31 October 2018).
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Available online: http://rdp.cme.msu.edu/classifier/ (accessed on 31 October 2018).
- Available online: http://greengenes.secondgenome.com/ (accessed on 31 October 2018).
- Higashimura, Y.; Naito, T.; Takagi, T.; Uchida, K.; Mizushima, K.; Ushiroda, C.; Ohnogi, H.; Kudo, Y.; Tasui, M.; Inui, S.; et al. Protective effect of agaro-oligosaccharides on gut dysbiosis and colon tumorigenesis in high-fat diet-fed mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G367–G375. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhou, F.; Ji, B.; Nout, R.M.J.; Fang, Q.; Yang, Z. Determination of organic acids evolution during apple cider fermentation using an improved HPLC analysis method. Eur. Food Res. Technol. 2008, 227, 1183–1190. [Google Scholar] [CrossRef] [Green Version]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damen, B.; Cloetens, L.; Broekaert, W.F.; François, I.; Lescroart, O.; Trogh, I.; Arnaut, F.; Welling, G.W.; Wijffels, J.; Delcour, J.A.; et al. Consumption of breads containing in situ-produced arabinoxylan oligosaccharides alters gastrointestinal effects in healthy volunteers. J. Nutr. 2012, 142, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.; de Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and SCFA. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Schwiertz, A.; Hold, G.I.; Duncan, S.H.; Gruhl, B.; Collins, M.D.; Lawson, P.A.; Flint, H.J.; Blaut, M. Anaerostipes caccae gen. nov., sp. nov., a new saccharolytic acetate-utilising butyrate-producing bacterium from human faeces. Syst. Appl. Microbiol. 2002, 25, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Matsumoto, K.; Okumura, T.; Yokoi, W.; Naito, E.; Yoshida, Y.; Nomoto, K.; Ito, M.; Sawada, H. Isolation of lactate-utilising butyrate-producing bacteria from human feces and in vivo administration of Anaerostipes caccae strain L2 and galacto-oligosaccharides in a rat model. FEMS Microbiol. Ecol. 2008, 66, 528–536. [Google Scholar] [CrossRef] [PubMed]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary fiber intervention on gut microbiota composition in healthy adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2018, 107, 965–983. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.A.; Mikkelsen, D.; le Paih, L.; Gidley, M.J. In vitro fermentation kinetics and end-products of cereal arabinoxylans and (1,3;1,4)-β-glucans by porcine faeces. J. Cereal Sci. 2011, 53, 53–58. [Google Scholar] [CrossRef]
- De Vries, J.; Miller, P.E.; Verbeke, K. Effects of cereal fiber on bowel function: A systematic review of intervention trials. World J. Gastroenterol. 2015, 21, 8952–8963. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.J.; Heaton, K.W. Increasing butyrate concentration in the distal colon by accelerating intestinal transit. Gut 1997, 41, 245–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roager, H.M.; Hansen, L.B.; Bahl, M.I.; Frandsen, H.L.; Carvalho, V.; Gobel, R.J.; Dalgaard, M.D.; Plichta, D.R.; Sparholt, M.H.; Vestergaard, H.; et al. Colonic transit time is related to bacterial metabolism and mucosal turnover in the gut. Nat. Microbiol. 2016, 1, 16093. [Google Scholar] [CrossRef] [PubMed]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimarăes, V.D.; Sokol, H.; Doré, J.; Corthier, G.; Furet, J.P. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Vemuri, R.; Gundamaraju, R.; Shastri, M.D.; Shukla, S.D.; Kalpurath, K.; Ball, M.; Tristram, S.; Shankar, E.M.; Ahuja, K.; Eri, R. Gut microbial changes, interactions, and their implications on human lifecycle: An ageing perspective. BioMed Res. Int. 2018, 2018, 4178607. [Google Scholar] [CrossRef] [PubMed]
- Mantis, N.J.; Rol, N.; Corthésy, B. Secretory IgA’s complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WB−BM− Group | WB+BM− Group | WB−BM+ Group | WB+BM+ Group | ||
|---|---|---|---|---|---|
| Subjects | N | 15 | 15 | 15 | 15 |
| Sex (male/female) | N | 5/10 | 5/10 | 5/10 | 4/11 |
| Age b | Y | 46.4 ± 8.80 | 46.4 ± 7.9 | 46.5 ± 8.0 | 46.4 ± 10.5 |
| Height | Cm | 160.6 ± 6.6 | 162.4 ± 8.6 | 161.5 ± 6.6 | 160.3 ± 6.3 |
| Body weight | Kg | 52.1 ± 8.8 | 58.3 ± 8.5 | 59.7 ± 15.1 | 53.0 ± 7.1 |
| Body mass index | kg/m2 | 20.1 ± 2.4 | 22.1 ± 2.4 | 22.9 ± 5.6 | 20.6 ± 2.8 |
| Body fat | % | 23.1 ± 6.4 | 27.5 ± 5.9 | 26.0 ± 9.8 | 24.9 ± 9.2 |
| Systolic blood pressure | mmHg | 114.7 ± 10.7 | 114.1 ± 11.7 | 114.5 ± 12.5 | 112.7 ± 11.9 |
| Diastolic blood pressure | mmHg | 74.1 ± 8.0 | 70.0 ± 8.2 | 71.6 ± 10.4 | 72.7 ± 10.1 |
| Pulse rate | bpm | 69.2 ± 10.3 | 65.2 ± 8.1 | 66.5 ± 8.0 | 69.7 ± 8.1 |
| WB−BM− | WB+BM− | WB−BM+ | WB+BM+ | |
|---|---|---|---|---|
| Cellulose | 4.5 g | 1.7 g | 2.8 g | ― |
| Wheat bran | ― | 6.0 g | ― | 6.0 g |
| BARLEYmax | ― | ― | 6.0 g | 6.0 g |
| Wheat flour | 17.9 g | 14.9 g | 13.8 g | 10.6 g |
| Egg | 4.4 g | 4.4 g | 4.4 g | 4.4 g |
| Margarine | 9.2 g | 9.2 g | 9.2 g | 9.2 g |
| Emulsifier | 0.5 g | 0.5 g | 0.5 g | 0.5 g |
| Brown sugar | 7.3 g | 7.3 g | 7.3 g | 7.3 g |
| Cocoa powder | 0.9 g | 0.9 g | 0.9 g | 0.9 g |
| Salt | 0.05 g | 0.05 g | 0.05 g | 0.05 g |
| Vanilla flavor | 1.2 g | 1.2 g | 1.2 g | 1.2 g |
| Caramel color | 0.2 g | ― | ― | ― |
| Total | 46.15g | 46.15 g | 46.15 g | 46.15 g |
| WB−BM− | WB+BM− | WB−BM+ | WB+BM+ | |
|---|---|---|---|---|
| Energy (kcal/100 g) | 454 | 447 | 455 | 448 |
| Protein (g/100 g) | 6.0 | 7.8 | 7.1 | 8.6 |
| Fat (g/100 g) | 22.1 | 22.6 | 22.2 | 23.4 |
| Available carbohydrate (g/100 g) | 50.9 | 46 | 49.6 | 43.5 |
| Total dietary fiber (g/100 g) | 12.7 | 13.0 | 12.0 | 12.1 |
| Soluble dietary fiber (g/100 g) | <0.5 | 1.1 | 1.2 | 1.8 |
| Insoluble dietary fiber (g/100 g) | 12.0 | 11.5 | 10.5 | 10.7 |
| β-glucan (g/100 g) | nd. | 0.4 | 0.8 | 1.2 |
| Arabinoxylan (g/100 g) b | nd. | 2.6 | 1.0 | 3.6 |
| Resistant starch (g/100 g) | nd. | nd. | 1.4 | 1.1 |
| Group | Average Daily Record | p-Value b | |
|---|---|---|---|
| Energy (kcal) | WB−BM− | 1403.0 ± 374.2 | 0.207 |
| WB+BM− | 1491.3 ± 446.8 | ||
| WB−BM+ | 1361.8 ± 320.2 | ||
| WB+BM+ | 1364.2 ± 361.9 | ||
| Protein (g) | WB−BM− | 51.7 ± 16.8 | 0.343 |
| WB+BM− | 54.3 ± 18.2 | ||
| WB−BM+ | 51.5 ± 13.5 | ||
| WB+BM+ | 48.9 ± 15.2 | ||
| Fat (g) | WB−BM− | 51.6 ± 20.7 | 0.261 |
| WB+BM− | 55.0 ± 26.5 | ||
| WB−BM+ | 47.6 ± 15.4 | ||
| WB+BM+ | 49.8 ± 20.0 | ||
| Carbohydrate (g) | WB−BM− | 174.5 ± 54.9 | 0.604 |
| WB+BM− | 185.7 ± 57.6 | ||
| WB−BM+ | 174.4 ± 50.9 | ||
| WB+BM+ | 175.4 ± 53.6 | ||
| Total dietary fiber (g) | WB−BM− | 8.8 ± 3.4 | 0.904 |
| WB+BM− | 8.3 ± 3.3 | ||
| WB−BM+ | 8.6 ± 3.4 | ||
| WB+BM+ | 8.6 ± 3.6 |
| p-Value (Two-Way ANOVA) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Phylum | Group | Baseline | Week 4 | WB | BM | WB × BM | |||
| Baseline | Week 4 | Baseline | Week 4 | Baseline | Week 4 | ||||
| Firmicutes % | WB−BM− | 64.0 ± 8.5 | 69.9 ± 13.6 | 0.53 | 0.79 | 0.60 | 0.86 | 0.73 | 0.98 |
| WB+BM− | 67.5 ± 14.0 | 69.0 ± 13.1 | |||||||
| WB−BM+ | 63.4 ±16.4 | 70.7 ± 16.0 | |||||||
| WB+BM+ | 64.4 ± 15.2 | 69.6 ± 14.8 | |||||||
| Actinobacteria % | WB−BM− | 28.8 ± 12.6 | 24.5 ± 13.4 | 0.48 | 1.00 | 0.64 | 0.96 | 0.75 | 0.56 |
| WB+BM− | 24.8 ± 13.0 | 26.7 ± 12.3 | |||||||
| WB−BM+ | 29.3 ± 16.0 | 26.5 ± 15.7 | |||||||
| WB+BM+ | 27.8 ± 16.4 | 24.3 ± 15.4 | |||||||
| Bacteroidetes % | WB−BM− | 6.0 ± 5.9 | 4.7 ± 2.7 | 0.34 | 0.26 | 0.50 | 0.71 | 0.60 | 0.02 |
| WB+BM− | 6.7 ± 6.5 | 3.6 ± 3.5 | |||||||
| WB−BM+ | 4.0 ± 4.7 | 2.0 ± 1.7 | |||||||
| WB+BM+ | 6.4 ± 7.7 | 5.5 ± 6.1 | |||||||
| Proteobacteria % | WB−BM− | 0.9 ± 1.0 | 0.6 ± 0.0 | 0.23 | 0.59 | 0.19 | 0.96 | 0.38 | 0.85 |
| WB+BM− | 0.5 ± 0.9 | 0.5 ± 1.2 | |||||||
| WB−BM+ | 3.0 ± 0.0 | 0.6 ± 1.3 | |||||||
| WB+BM+ | 1.0 ± 1.9 | 0.4 ± 0.4 | |||||||
| p-Value (Two-Way ANOVA) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Genus | Group | Baseline | Week 4 | WB | BM | WB × BM | |||
| Baseline | Week 4 | Baseline | Week 4 | Baseline | Week 4 | ||||
| Bacteroides % | WB−BM− | 4.2 ± 4.8 | 3.1 ± 1.8 | 0.34 | 0.31 | 0.46 | 0.87 | 0.70 | 0.03 |
| WB+BM− | 4.9 ± 5.7 | 2.2 ± 2.2 | |||||||
| WB−BM+ | 2.7 ± 3.9 | 1.4 ± 1.3 | |||||||
| WB+BM+ | 4.5 ± 5.8 | 3.7 ± 4.5 | |||||||
| Bifidobacterium % | WB−BM− | 19.5 ± 10.3 | 17.2 ± 12.7 | 0.23 | 0.77 | 0.52 | 0.97 | 0.50 | 0.82 |
| WB+BM− | 13.6 ± 10.8 | 17.0 ± 13.4 | |||||||
| WB−BM+ | 19.4 ± 13.1 | 18.0 ± 10.7 | |||||||
| WB+BM+ | 17.7 ± 13.5 | 16.4 ± 12.5 | |||||||
| Lactobacillus % | WB−BM− | 0.2 ± 0.4 | 0.4 ± 1.0 | 0.55 | 0.96 | 0.74 | 0.33 | 0.41 | 0.44 |
| WB+BM− | 0.2 ± 0.6 | 0.7 ± 2.4 | |||||||
| WB−BM+ | 0.4 ± 0.9 | 0.4 ± 0.7 | |||||||
| WB+BM+ | 0.1 ± 0.4 | 0.1 ± 0.2 | |||||||
| Prevotella % | WB−BM− | 0.1 ± 0.2 | 0.2 ± 0.4 | 0.45 | 0.38 | 0.47 | 0.42 | 0.58 | 0.20 |
| WB+BM− | 0.1 ± 0.2 | 0.1 ± 0.2 | |||||||
| WB−BM+ | 0.4 ± 1.5 | 0.1 ± 0.3 | |||||||
| WB+BM+ | 0.1 ± 0.3 | 0.4 ± 0.9 | |||||||
| Clostridium % | WB−BM− | 6.8 ± 4.5 | 4.9 ± 3.4 | 0.66 | 0.67 | 0.55 | 0.35 | 0.06 | 0.36 |
| WB+BM− | 4.4 ± 3.3 | 3.6 ± 2.9 | |||||||
| WB−BM+ | 4.2 ± 3.8 | 5.0 ± 4.1 | |||||||
| WB+BM+ | 5.7 ± 4.6 | 5.5 ± 5.0 | |||||||
| Anaerostipes % | WB−BM− | 3.5 ± 2.5 | 5.2 ± 5.3 | 0.97 | <0.01 | 0.29 | 0.71 | 0.14 | 0.69 |
| WB+BM− | 2.0 ± 2.3 | 2.2 ± 2.2 | |||||||
| WB−BM+ | 3.1 ±3.6 | 4.5 ± 4.3 | |||||||
| WB+BM+ | 4.7 ± 6.2 | 2.2 ± 1.8 | |||||||
| Butyrate producers b % | WB−BM− | 12.7 ± 7.4 | 8.7 ± 4.4 | 0.77 | 0.05 | 0.03 | 0.76 | 0.22 | 0.80 |
| WB+BM− | 11.1 ± 6.4 | 12.0 ± 7.1 | |||||||
| WB−BM+ | 6.9 ± 6.5 | 8.6 ± 4.5 | |||||||
| WB+BM+ | 9.5 ± 5.7 | 11.2 ± 5.4 | |||||||
| p-Value (Two-Way ANOVA) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Diversity Index | Group | Baseline | Week 4 | WB | BM | WB × BM | |||
| Baseline | Week 4 | Baseline | Week 4 | Baseline | Week 4 | ||||
| Phylogenetic diversity (PD_whole_tree) | WB−BM− | 27.9 ± 4.3 | 27.1 ± 6.3 | 0.38 | 0.93 | 0.54 | 0.91 | 0.70 | 0.81 |
| WB+BM− | 28.9 ± 10.0 | 27.4 ± 7.5 | |||||||
| WB−BM+ | 25.9 ± 8.5 | 27.4 ± 7.8 | |||||||
| WB+BM+ | 28.5 ± 6.5 | 26.8 ± 6.0 | |||||||
| Chao1 | WB−BM− | 1123.3 ± 184.5 | 1144.4 ± 294.7 | 0.24 | 0.90 | 0.34 | 0.62 | 0.96 | 0.89 |
| WB+BM− | 1230.4 ± 451.4 | 1166.4 ± 322.4 | |||||||
| WB−BM+ | 1044.8 ± 348.8 | 1115.6 ± 332.7 | |||||||
| WB+BM+ | 1142.9 ± 286.3 | 1114.1 ± 300.9 | |||||||
| Observed number of OTUs (observed_species) | WB−BM− | 630.4 ± 101.6 | 626.5 ± 149.6 | 0.20 | 0.74 | 0.35 | 0.89 | 0.98 | 0.73 |
| WB+BM− | 688.7 ± 251.4 | 655.0 ± 165.4 | |||||||
| WB−BM+ | 585.8 ± 180.7 | 635.4 ± 175.8 | |||||||
| WB+BM+ | 646.4 ± 150.8 | 634.8 ± 167.5 | |||||||
| Shannon index | WB−BM− | 5.5 ± 0.5 | 5.6 ± 0.5 | 0.15 | 0.70 | 0.39 | 0.79 | 0.45 | 0.28 |
| WB+BM− | 5.6 ± 0.7 | 5.5 ± 0.5 | |||||||
| WB−BM+ | 5.2 ± 0.7 | 5.4 ± 0.5 | |||||||
| WB+BM+ | 5.6 ± 0.6 | 5.7 ± 0.7 | |||||||
| p-Value (Two-Way ANOVA) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Phylum | Group | Baseline | Week 4 | WB | BM | WB × BM | |||
| Baseline | Week 4 | Baseline | Week 4 | Baseline | Week 4 | ||||
| Acetate (mg/g feces) | WB−BM− | 2.18 ± 1.42 | 1.96 ± 1.10 | 0.60 | 0.04 | 0.42 | 0.39 | 0.35 | 0.90 |
| WB+BM− | 2.30 ± 0.88 | 2.48 ± 0.98 | |||||||
| WB−BM+ | 2.73 ± 1.17 | 2.16 ± 0.97 | |||||||
| WB+BM+ | 2.26 ± 1.19 | 2.74 ± 0.94 | |||||||
| Propionate (mg/g feces) | WB−BM− | 1.09 ± 0.59 | 0.90 ± 0.37 | 0.55 | 0.16 | 0.80 | 0.70 | 0.90 | 0.43 |
| WB+BM− | 1.02 ± 0.47 | 1.16 ± 0.66 | |||||||
| WB−BM+ | 1.14 ± 0.40 | 0.95 ± 0.31 | |||||||
| WB+BM+ | 1.04 ± 0.57 | 1.02 ± 0.39 | |||||||
| iso-Butyrate (mg/g feces) | WB−BM− | 0.13 ± 0.08 | 0.12 ± 0.06 | 0.60 | 0.66 | 0.24 | 0.89 | 0.77 | 0.88 |
| WB+BM− | 0.11 ± 0.08 | 0.12 ± 0.07 | |||||||
| WB−BM+ | 0.15 ± 0.11 | 0.13 ± 0.07 | |||||||
| WB+BM+ | 0.15 ± 0.09 | 0.12 ± 0.06 | |||||||
| n-Butyrate (mg/g feces) | WB−BM− | 0.89 ± 0.86 | 0.67 ± 0.25 | 0.53 | 0.05 | 0.98 | 0.93 | 0.97 | 0.59 |
| WB+BM− | 0.78 ± 0.53 | 0.96 ± 0.62 | |||||||
| WB−BM+ | 0.88 ± 0.47 | 0.74 ± 0.36 | |||||||
| WB+BM+ | 0.78 ± 0.54 | 0.91 ± 0.47 | |||||||
| Total SCFA (mg/g feces) | WB−BM− | 4.64 ± 2.85 | 3.94 ± 1.45 | 0.53 | 0.04 | 0.50 | 0.58 | 0.67 | 0.81 |
| WB+BM− | 4.52 ± 1.85 | 4.98 ± 2.03 | |||||||
| WB−BM+ | 5.29 ± 1.72 | 4.29 ± 1.63 | |||||||
| WB+BM+ | 4.66 ± 2.32 | 5.12 ± 1.61 | |||||||
| Putrefaction Product | p-Value (Two-Way ANOVA) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Group | Baseline | Week 4 | WB | BM | WB × BM | ||||
| Baseline | Week 4 | Baseline | Week 4 | Baseline | Week 4 | ||||
| Ammonium (mg/g) | WB−BM− | 0.5 ± 0.2 | 0.5 ± 0.2 | 0.777 | 0.872 | 0.568 | 0.701 | 0.367 | 0.892 |
| WB+BM− | 0.4 ± 0.2 | 0.5 ± 0.2 | |||||||
| WB−BM+ | 0.5 ± 0.3 | 0.5 ± 0.2 | |||||||
| WB+BM+ | 0.5 ± 0.4 | 0.4 ± 0.2 | |||||||
| p-Cresol (μg/g) | WB−BM− | 75.8 ± 67.4 | 59.6 ± 57.3 | 0.129 | 0.043 | 0.191 | 0.284 | 0.616 | 0.416 |
| WB+BM− | 53.7 ± 58.9 | 41.8 ± 51.2 | |||||||
| WB−BM+ | 113.5 ± 110.1 | 86.1 ± 60.9 | |||||||
| WB+BM+ | 70.7 ± 72.3 | 45.6 ± 42.0 | |||||||
| Indole (μg/g) | WB−BM− | 46.7 ± 30.8 | 33.7 ± 23.4 | 0.040 | 0.070 | 0.459 | 0.546 | 0.503 | 0.720 |
| WB+BM− | 25.6 ± 19.8 | 21.6 ± 12.4 | |||||||
| WB−BM+ | 47.1 ± 39.5 | 35.0 ± 24.0 | |||||||
| WB+BM+ | 36.3 ± 22.9 | 26.8 ± 22.6 | |||||||
| Total putrefaction products (except ammonium) (μg/g) | WB−BM− | 134.4 ± 92.1 | 102.2 ± 67.9 | 0.604 | 0.037 | 0.421 | 0.385 | 0.346 | 0.896 |
| WB+BM− | 89.6 ± 74.1 | 72.9 ± 63.8 | |||||||
| WB−BM+ | 174.6 ± 161.4 | 127.5 ± 83.7 | |||||||
| WB+BM+ | 121.4 ± 91.0 | 81.5 ± 50.0 | |||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aoe, S.; Nakamura, F.; Fujiwara, S. Effect of Wheat Bran on Fecal Butyrate-Producing Bacteria and Wheat Bran Combined with Barley on Bacteroides Abundance in Japanese Healthy Adults. Nutrients 2018, 10, 1980. https://doi.org/10.3390/nu10121980
Aoe S, Nakamura F, Fujiwara S. Effect of Wheat Bran on Fecal Butyrate-Producing Bacteria and Wheat Bran Combined with Barley on Bacteroides Abundance in Japanese Healthy Adults. Nutrients. 2018; 10(12):1980. https://doi.org/10.3390/nu10121980
Chicago/Turabian StyleAoe, Seiichiro, Fumiko Nakamura, and Suguru Fujiwara. 2018. "Effect of Wheat Bran on Fecal Butyrate-Producing Bacteria and Wheat Bran Combined with Barley on Bacteroides Abundance in Japanese Healthy Adults" Nutrients 10, no. 12: 1980. https://doi.org/10.3390/nu10121980
APA StyleAoe, S., Nakamura, F., & Fujiwara, S. (2018). Effect of Wheat Bran on Fecal Butyrate-Producing Bacteria and Wheat Bran Combined with Barley on Bacteroides Abundance in Japanese Healthy Adults. Nutrients, 10(12), 1980. https://doi.org/10.3390/nu10121980