Myoprotective Potential of Creatine Is Greater than Whey Protein after Chemically-Induced Damage in Rat Skeletal Muscle



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Procedures
2.3. Assessment of Contractile Properties
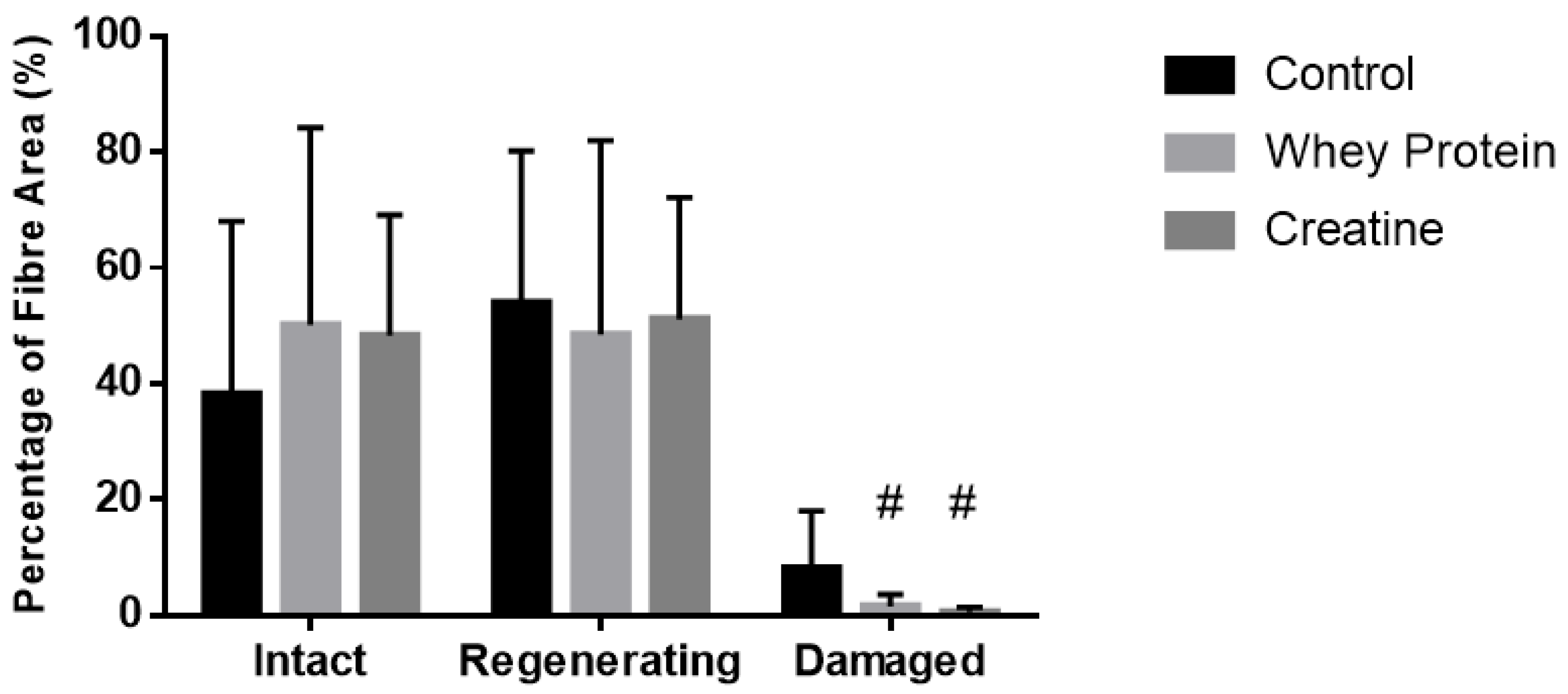
2.4. Histological Analysis
2.5. Protein Analysis
2.6. Statistical Analysis
3. Results
3.1. Supplement Ingestion
3.2. Body Weight and Muscle Mass
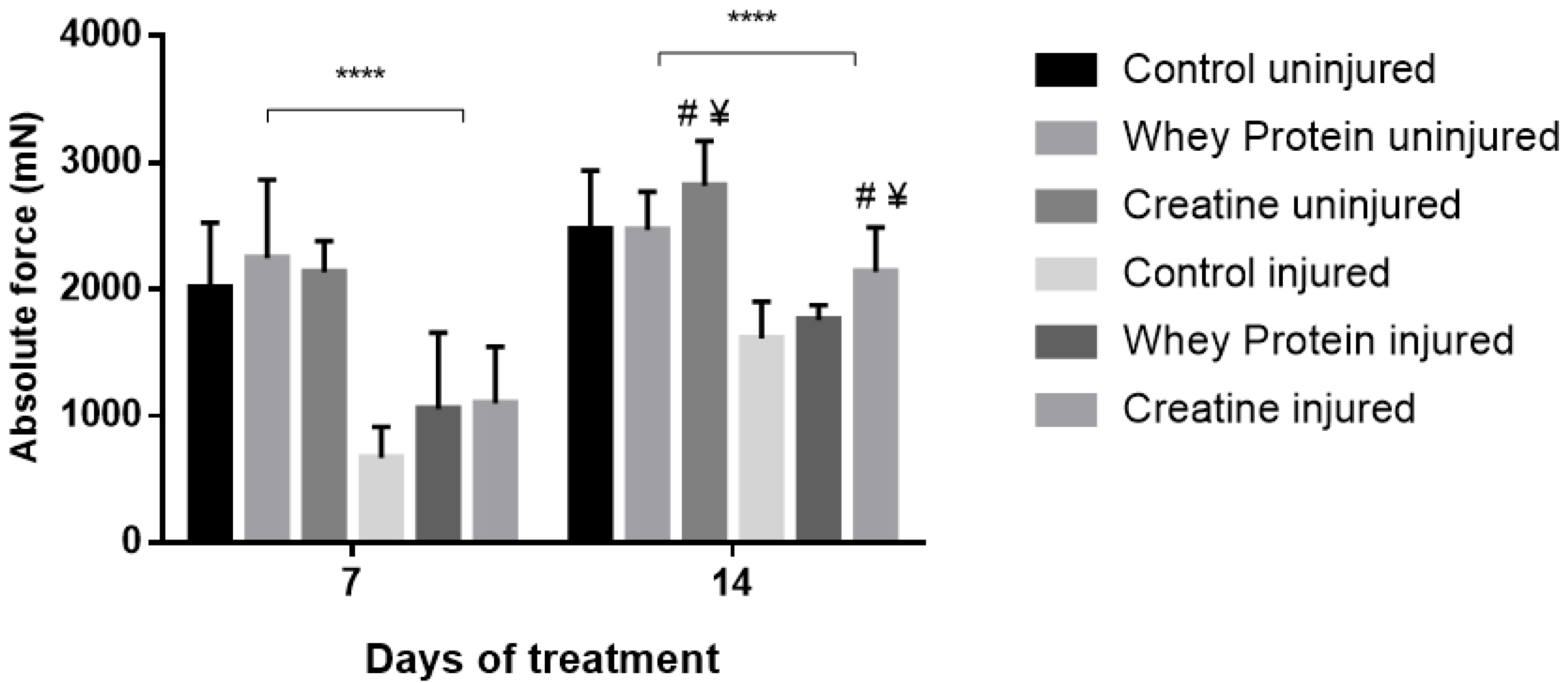
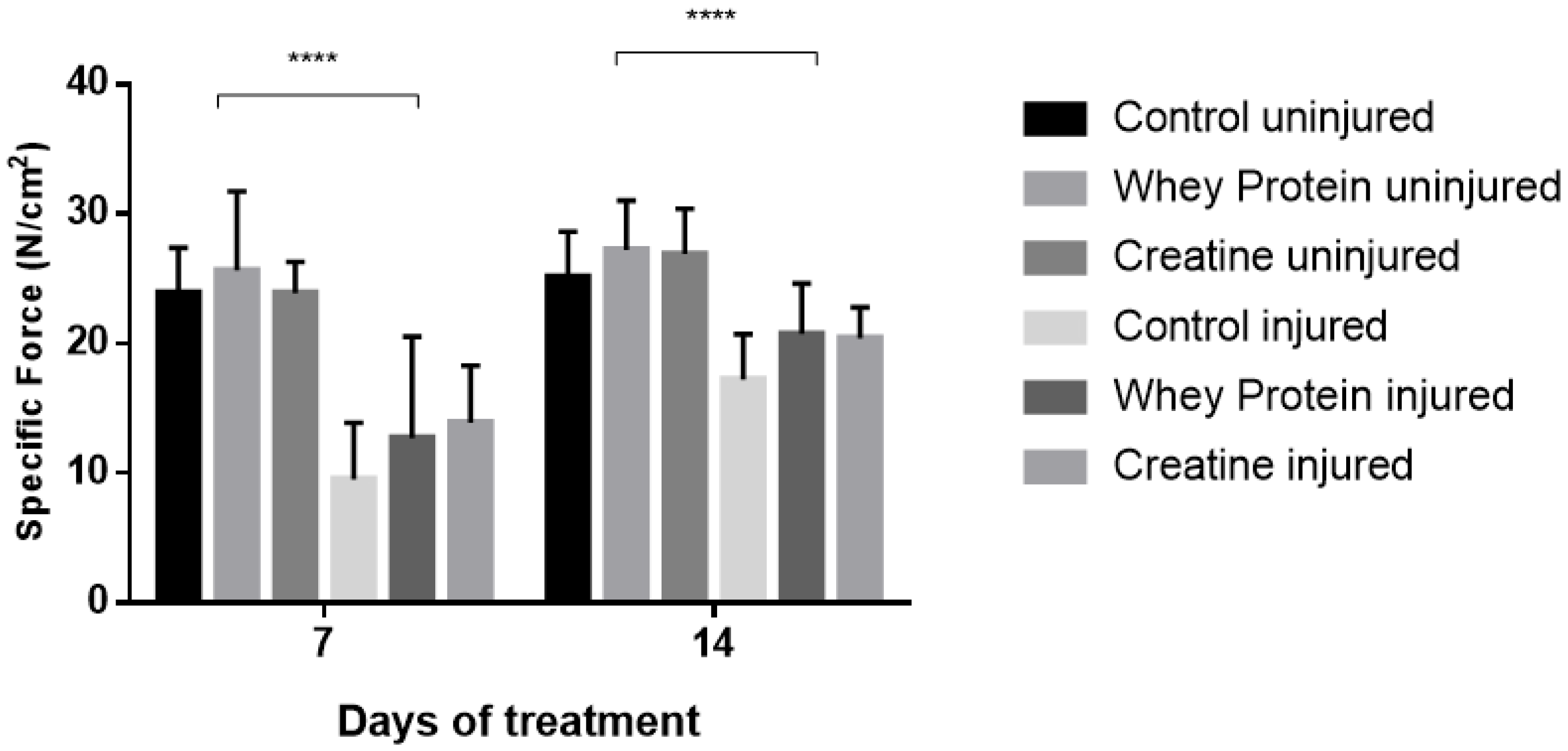
3.3. Contractile Properties
3.4. Total and Contractile Protein
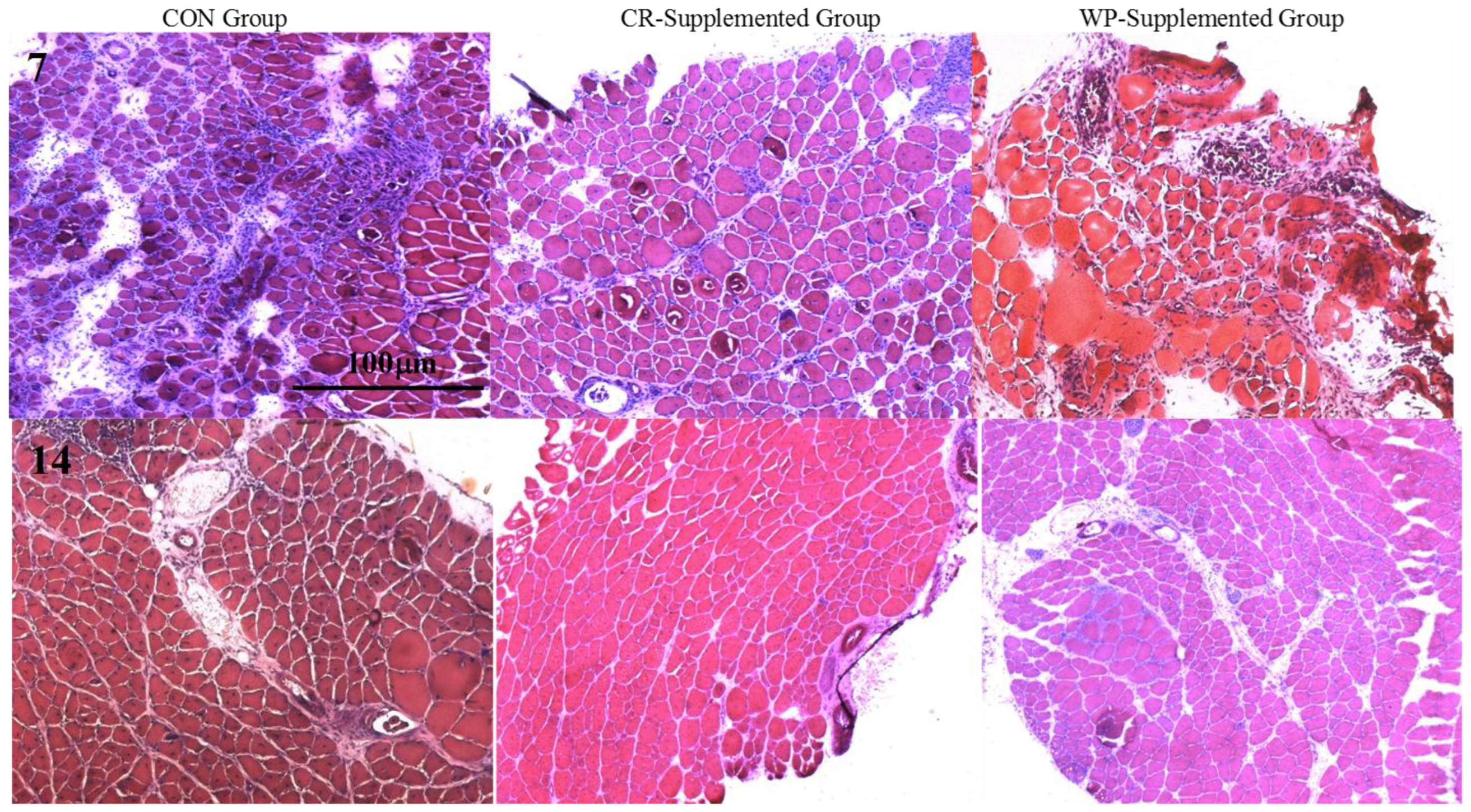
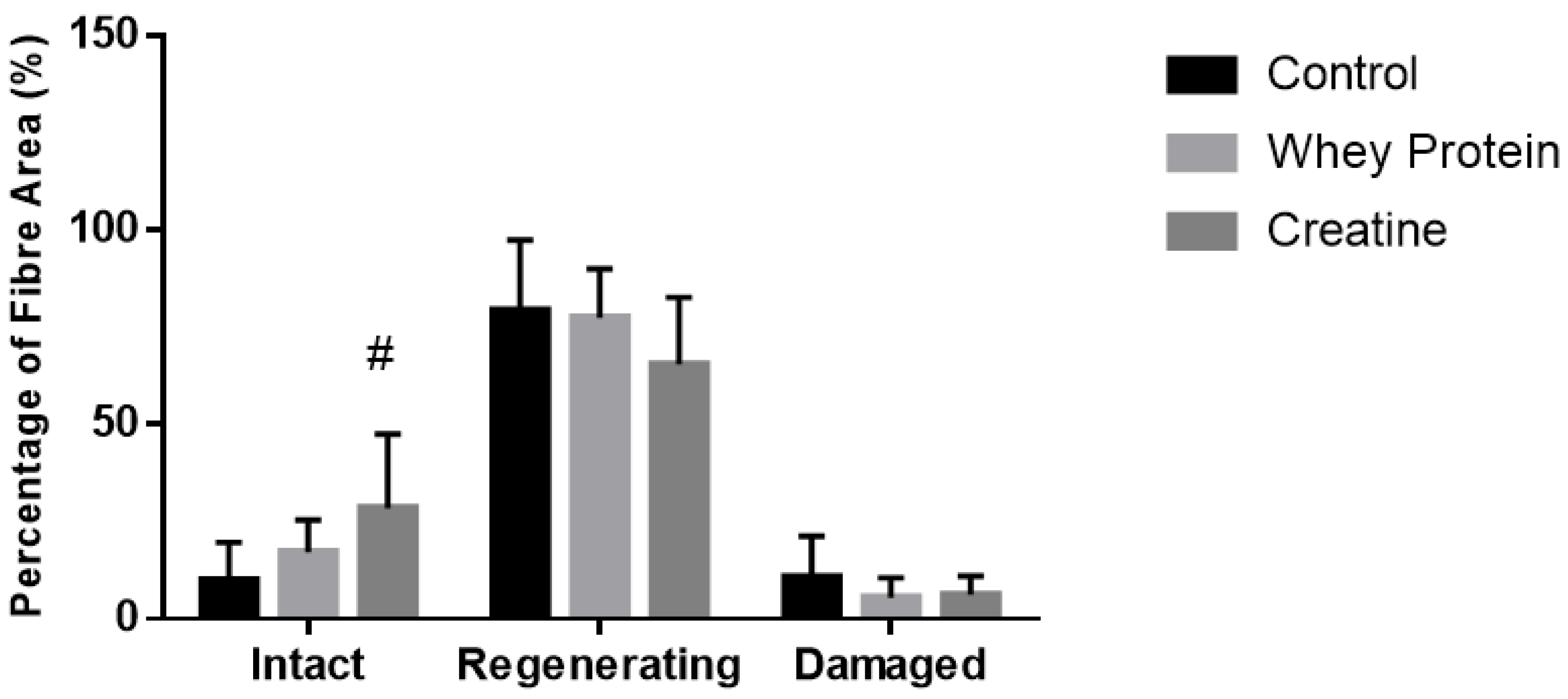
3.5. Histological Analysis
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Huard, J.; Li, Y.; Fu, F.H. Muscle injuries and repair: Current trends in research. J. Bone Jt. Surg Am. 2002, 84, 822–832. [Google Scholar] [CrossRef]
- Tidball, J.G. Inflammatory processes in muscle injury and repair. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R345–R353. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.; Kruger, M.J.; Smith, R.M.; Myburgh, K.H. The inflammatory response to skeletal muscle injury: Illuminating complexities. Sports Med. 2008, 38, 947–969. [Google Scholar] [CrossRef] [PubMed]
- Tidball, J.G. Mechanisms of muscle injury, repair, and regeneration. Compr. Physiol. 2011, 1, 2029–2062. [Google Scholar] [PubMed]
- Sorichter, S.; Puschendorf, B.; Mair, J. Skeletal muscle injury induced by eccentric muscle action: Muscle proteins as markers of muscle fiber injury. Exerc. Immunol. Rev. 1999, 5, 5–21. [Google Scholar] [PubMed]
- Gissel, H.; Clausen, T. Excitation-induced ca(2+) influx in rat soleus and edl muscle: Mechanisms and effects on cellular integrity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R917–R924. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.G.; Whitehead, N.P.; Yeung, E.W. Mechanisms of stretch-induced muscle damage in normal and dystrophic muscle: Role of ionic changes. J. Physiol. 2005, 567, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.; Naclerio, F.; Allgrove, J.; Jimenez, A. Creatine supplementation with specific view to exercise/sports performance: An update. J. Int. Soc. Sports Nutr. 2012, 9, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, R.W.; Carson, B.P.; Jakeman, P.M. The effect of whey protein supplementation on the temporal recovery of muscle function following resistance training: A systematic review and meta-analysis. Nutrients 2018, 10, 221. [Google Scholar] [CrossRef] [PubMed]
- Cooke, M.B.; Rybalka, E.; Williams, A.D.; Cribb, P.J.; Hayes, A. Creatine supplementation enhances muscle force recovery after eccentrically-induced muscle damage in healthy individuals. J. Int. Soc. Sports Nutr. 2009, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Rawson, E.S.; Conti, M.P.; Miles, M.P. Creatine supplementation does not reduce muscle damage or enhance recovery from resistance exercise. J. Strength Cond. Res. 2007, 21, 1208–1213. [Google Scholar] [PubMed]
- Rawson, E.S.; Gunn, B.; Clarkson, P.M. The effects of creatine supplementation on exercise-induced muscle damage. J. Strength Cond. Res. 2001, 15, 178–184. [Google Scholar] [PubMed]
- McKinnon, N.B.; Graham, M.T.; Tiidus, P.M. Effect of creatine supplementation on muscle damage and repair following eccentrically-induced damage to the elbow flexor muscles. J. Sports Sci. Med. 2012, 11, 653–659. [Google Scholar] [PubMed]
- Veggi, K.F.; Machado, M.; Koch, A.J.; Santana, S.C.; Oliveira, S.S.; Stec, M.J. Oral creatine supplementation augments the repeated bout effect. Int. J. Sport Nutr. Exerc. Metab. 2013, 23, 378–387. [Google Scholar] [CrossRef]
- Rosene, J.; Matthews, T.; Ryan, C.; Belmore, K.; Bergsten, A.; Blaisdell, J.; Gaylord, J.; Love, R.; Marrone, M.; Ward, K.; et al. Short and longer-term effects of creatine supplementation on exercise induced muscle damage. J. Sports Sci. Med. 2009, 8, 89–96. [Google Scholar] [PubMed]
- White, J.P.; Wilson, J.M.; Austin, K.G.; Greer, B.K.; St John, N.; Panton, L.B. Effect of carbohydrate-protein supplement timing on acute exercise-induced muscle damage. J. Int. Soc. Sports Nutr. 2008, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Betts, J.A.; Toone, R.J.; Stokes, K.A.; Thompson, D. Systemic indices of skeletal muscle damage and recovery of muscle function after exercise: Effect of combined carbohydrate-protein ingestion. Appl. Physiol. Nutr. Metab. 2009, 34, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Nosaka, K.; Clarkson, P.M. Changes in indicators of inflammation after eccentric exercise of the elbow flexors. Med. Sci. Sports Exerc. 1996, 28, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Nosaka, K.; Sacco, P.; Mawatari, K. Effects of amino acid supplementation on muscle soreness and damage. Int. J. Sport Nutr. Exerc. Metab. 2006, 16, 620–635. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, Y.; Yamamoto, Y.; Bajotto, G.; Sato, J.; Murakami, T.; Shimomura, N.; Kobayashi, H.; Mawatari, K. Nutraceutical effects of branched-chain amino acids on skeletal muscle. J. Nutr. 2006, 136, 529S–532S. [Google Scholar] [CrossRef] [PubMed]
- Cockburn, E.; Hayes, P.R.; French, D.N.; Stevenson, E.; St Clair Gibson, A. Acute milk-based protein-cho supplementation attenuates exercise-induced muscle damage. Appl. Physiol. Nutr. Metab. 2008, 33, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Cockburn, E.; Stevenson, E.; Hayes, P.R.; Robson-Ansley, P.; Howatson, G. Effect of milk-based carbohydrate-protein supplement timing on the attenuation of exercise-induced muscle damage. Appl. Physiol. Nutr. Metab. 2010, 35, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Buckley, J.D.; Thomson, R.L.; Coates, A.M.; Howe, P.R.; DeNichilo, M.O.; Rowney, M.K. Supplementation with a whey protein hydrolysate enhances recovery of muscle force-generating capacity following eccentric exercise. J. Sci. Med. Sport 2010, 13, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Etheridge, T.; Philp, A.; Watt, P.W. A single protein meal increases recovery of muscle function following an acute eccentric exercise bout. Appl. Physiol. Nutr. Metab. 2008, 33, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Cooke, M.B.; Rybalka, E.; Stathis, C.G.; Cribb, P.J.; Hayes, A. Whey protein isolate attenuates strength decline after eccentrically-induced muscle damage in healthy individuals. J. Int. Soc. Sports Nutr. 2010, 7, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassit, R.A.; Pinheiro, C.H.; Vitzel, K.F.; Sproesser, A.J.; Silveira, L.R.; Curi, R. Effect of short-term creatine supplementation on markers of skeletal muscle damage after strenuous contractile activity. Eur. J. Appl. Physiol. 2010, 108, 945–955. [Google Scholar] [CrossRef] [PubMed]
- Crassous, B.; Richard-Bulteau, H.; Deldicque, L.; Serrurier, B.; Pasdeloup, M.; Francaux, M.; Bigard, X.; Koulmann, N. Lack of effects of creatine on the regeneration of soleus muscle after injury in rats. Med. Sci. Sports Exerc. 2009, 41, 1761–1769. [Google Scholar] [CrossRef] [PubMed]
- Warren, G.L.; Fennessy, J.M.; Millard-Stafford, M.L. Strength loss after eccentric contractions is unaffected by creatine supplementation. J. Appl. Physiol. 2000, 89, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Areta, J.L.; Burke, L.M.; Ross, M.L.; Camera, D.M.; West, D.W.; Broad, E.M.; Jeacocke, N.A.; Moore, D.R.; Stellingwerff, T.; Phillips, S.M.; et al. Timing and distribution of protein ingestion during prolonged recovery from resistance exercise alters myofibrillar protein synthesis. J. Physiol. 2013, 591, 2319–2331. [Google Scholar] [CrossRef] [PubMed]
- Rosenblatt, J.D. A time course study of the isometric contractile properties of rat extensor digitorum longus muscle injected with bupivacaine. Comp. Biochem. Physiol. Comp. Physiol. 1992, 101, 361–367. [Google Scholar] [CrossRef]
- Goodman, C.A.; Horvath, D.; Stathis, C.; Mori, T.; Croft, K.; Murphy, R.M.; Hayes, A. Taurine supplementation increases skeletal muscle force production and protects muscle function during and after high-frequency in vitro stimulation. J. Appl. Physiol. (1985) 2009, 107, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Brooks, S.V.; Faulkner, J.A. Contractile properties of skeletal muscles from young, adult and aged mice. J. Physiol. 1988, 404, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Mendez, J.; Keys, A. Density and composition of mammalian muscle. Metabolism 1960, 9, 184–188. [Google Scholar]
- Timpani, C.A.; Trewin, A.J.; Stojanovska, V.; Robinson, A.; Goodman, C.A.; Nurgali, K.; Betik, A.C.; Stepto, N.; Hayes, A.; McConell, G.K.; et al. Attempting to compensate for reduced neuronal nitric oxide synthase protein with nitrate supplementation cannot overcome metabolic dysfunction but rather has detrimental effects in dystrophin-deficient mdx muscle. Neurotherapeutics 2017, 14, 429–446. [Google Scholar] [CrossRef] [PubMed]
- Pompeani, N.; Rybalka, E.; Latchman, H.; Murphy, R.M.; Croft, K.; Hayes, A. Skeletal muscle atrophy in sedentary zucker obese rats is not caused by calpain-mediated muscle damage or lipid peroxidation induced by oxidative stress. J. Negat Results Biomed. 2014, 13, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grounds, M. Quantification of histopathology in haemotoxylin and eosin stained muscle sections. In Standard Operating Procedures; TREAT-NMD: Washington, DC, USA, 2010. [Google Scholar]
- Beitzel, F.; Gregorevic, P.; Ryall, J.G.; Plant, D.R.; Sillence, M.N.; Lynch, G.S. Beta2-adrenoceptor agonist fenoterol enhances functional repair of regenerating rat skeletal muscle after injury. J. Appl. Physiol. 2004, 96, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hall-Craggs, E.C. Rapid degeneration and regeneration of a whole skeletal muscle following treatment with bupivacaine (marcain). Exp. Neurol. 1974, 43, 349–358. [Google Scholar] [CrossRef]
- Plant, D.R.; Beitzel, F.; Lynch, G.S. Length-tension relationships are altered in regenerating muscles of the rat after bupivacaine injection. J. Appl. Physiol. 2005, 98, 1998–2003. [Google Scholar] [CrossRef] [PubMed]
- Becque, M.D.; Lochmann, J.D.; Melrose, D.R. Effects of oral creatine supplementation on muscular strength and body composition. Med. Sci. Sports Exerc. 2000, 32, 654–658. [Google Scholar] [CrossRef] [PubMed]
- Chrusch, M.J.; Chilibeck, P.D.; Chad, K.E.; Davison, K.S.; Burke, D.G. Creatine supplementation combined with resistance training in older men. Med. Sci. Sports Exerc. 2001, 33, 2111–2117. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, A.F.; Aguiar, D.H.; Felisberto, A.D.; Carani, F.R.; Milanezi, R.C.; Padovani, C.R.; Dal-Pai-Silva, M. Effects of creatine supplementation during resistance training on myosin heavy chain (mhc) expression in rat skeletal muscle fibers. J. Strength Cond. Res. 2010, 24, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Antonio, J.; Ciccone, V. The effects of pre versus post workout supplementation of creatine monohydrate on body composition and strength. J. Int. Soc. Sports Nutr. 2013, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Olsen, S.; Aagaard, P.; Kadi, F.; Tufekovic, G.; Verney, J.; Olesen, J.L.; Suetta, C.; Kjaer, M. Creatine supplementation augments the increase in satellite cell and myonuclei number in human skeletal muscle induced by strength training. J. Physiol. 2006, 573, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Parise, G.; Mihic, S.; MacLennan, D.; Yarasheski, K.E.; Tarnopolsky, M.A. Effects of acute creatine monohydrate supplementation on leucine kinetics and mixed-muscle protein synthesis. J. Appl. Physiol. 2001, 91, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Louis, M.; Poortmans, J.R.; Francaux, M.; Berre, J.; Boisseau, N.; Brassine, E.; Cuthbertson, D.J.; Smith, K.; Babraj, J.A.; Waddell, T.; et al. No effect of creatine supplementation on human myofibrillar and sarcoplasmic protein synthesis after resistance exercise. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E1089–E1094. [Google Scholar] [CrossRef] [PubMed]
- Louis, M.; Van Beneden, R.; Dehoux, M.; Thissen, J.P.; Francaux, M. Creatine increases igf-i and myogenic regulatory factor mrna in c(2)c(12) cells. FEBS Lett. 2004, 557, 243–247. [Google Scholar] [CrossRef]
- Sestili, P.; Barbieri, E.; Martinelli, C.; Battistelli, M.; Guescini, M.; Vallorani, L.; Casadei, L.; D'Emilio, A.; Falcieri, E.; Piccoli, G.; et al. Creatine supplementation prevents the inhibition of myogenic differentiation in oxidatively injured c2c12 murine myoblasts. Mol. Nutr. Food Res. 2009, 53, 1187–1204. [Google Scholar] [CrossRef] [PubMed]
- Deldicque, L.; Theisen, D.; Bertrand, L.; Hespel, P.; Hue, L.; Francaux, M. Creatine enhances differentiation of myogenic c2c12 cells by activating both p38 and akt/pkb pathways. Am. J. Physiol. Cell. Physiol. 2007, 293, C1263–C1271. [Google Scholar] [CrossRef] [PubMed]
- Volek, J.S.; Duncan, N.D.; Mazzetti, S.A.; Staron, R.S.; Putukian, M.; Gomez, A.L.; Pearson, D.R.; Fink, W.J.; Kraemer, W.J. Performance and muscle fiber adaptations to creatine supplementation and heavy resistance training. Med. Sci. Sports Exerc. 1999, 31, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Sestili, P.; Ambrogini, P.; Barbieri, E.; Sartini, S.; Fimognari, C.; Calcabrini, C.; Diaz, A.R.; Guescini, M.; Polidori, E.; Luchetti, F.; et al. New insights into the trophic and cytoprotective effects of creatine in in vitro and in vivo models of cell maturation. Amino Acids 2016, 48, 1897–1911. [Google Scholar] [CrossRef] [PubMed]
- Wallimann, T.; Tokarska-Schlattner, M.; Schlattner, U. The creatine kinase system and pleiotropic effects of creatine. Amino Acids 2011, 40, 1271–1296. [Google Scholar] [CrossRef] [PubMed]
- Smith, I.J.; Dodd, S.L. Calpain activation causes a proteasome-dependent increase in protein degradation and inhibits the akt signalling pathway in rat diaphragm muscle. Exp. Physiol. 2007, 92, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Passaquin, A.C.; Renard, M.; Kay, L.; Challet, C.; Mokhtarian, A.; Wallimann, T.; Ruegg, U.T. Creatine supplementation reduces skeletal muscle degeneration and enhances mitochondrial function in mdx mice. Neuromuscul. Disord. 2002, 12, 174–182. [Google Scholar] [CrossRef]
- Korge, P.; Byrd, S.K.; Campbell, K.B. Functional coupling between sarcoplasmic-reticulum-bound creatine kinase and Ca(2+)-atpase. Eur. J. Biochem. 1993, 213, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Zima, A.V.; Blatter, L.A. Redox regulation of cardiac calcium channels and transporters. Cardiovasc. Res. 2006, 71, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Dolder, M.; Walzel, B.; Speer, O.; Schlattner, U.; Wallimann, T. Inhibition of the mitochondrial permeability transition by creatine kinase substrates. Requirement for microcompartmentation. J. Biol Chem 2003, 278, 17760–17766. [Google Scholar] [CrossRef] [PubMed]
- Fimognari, C.; Sestili, P.; Lenzi, M.; Cantelli-Forti, G.; Hrelia, P. Protective effect of creatine against rna damage. Mutat Res. 2009, 670, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Rybalka, E. Impaired Metabolism in x-Linked Muscular Dystrophy: Experimental Evaluation of Potential Therapies to Improve Calcium Regulation, Bioenergetics and Muscle Architecture; Victoria University: Melbourne, Australia, 2008. [Google Scholar]
- Greenhaff, P.L. Creatine and its application as an ergogenic aid. Int. J. Sport Nutr. 1995, 5, S100–S110. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, F.; Paulsen, G.; Raastad, T.; Bergersen, L.H.; Owe, S.G. Gross ultrastructural changes and necrotic fiber segments in elbow flexor muscles after maximal voluntary eccentric action in humans. J. Appl. Physiol. 2009, 107, 1923–1934. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Yi, S.; Zhang, H.; Wang, J. Ingestion of soy-whey blended protein augments sports performance and ameliorates exercise-induced fatigue in a rat exercise model. Food Funct 2017, 8, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.C.; Huang, W.C.; Chiu, C.C.; Chang, Y.K.; Huang, C.C. Whey protein improves exercise performance and biochemical profiles in trained mice. Med. Sci. Sports Exerc. 2014, 46, 1517–1524. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1 | Initial BW (g) | BW (g) after 14 Days | BW (g) after 21 Days |
| CON (n = 9) | 184.8 ± 23.6 | 209.4 ± 27.9 | 222.5 ± 24.2 |
| CR (n = 7) | 201.4 ± 16.4 | 232.3 ± 20.0 # | 248 ± 18.5 |
| Group 2 | Initial BW (g) | BW (g) after 14 days | BW (g) after 28 days |
| CON (n = 9) | 217.3 ± 15.7 | 243.7 ± 19.9 | 259.2 ± 21.4 |
| CR (n = 8) | 210.6 ± 14.8 | 242.0 ± 19.1 | 260.1 ± 19.5 |
| Group 3 | Initial BW (g) | BW (g) after 7 days | |
| WP (n = 9) | 224.8 ± 24.8 | 234.4 ± 18.1 | |
| Group 4 | Initial BW (g) | BW (g) after 14 days | |
| WP (n = 9) | 232.9 ± 22.3 | 249.1 ± 28.6 |
| Treatment | CON-INJ | WP-INJ | CR-INJ | CON-NORM | WP-NORM | CR-NORM |
|---|---|---|---|---|---|---|
| Duration | ||||||
| 7 days | ||||||
| n | 8 | 8 | 7 | 8 | 8 | 7 |
| MM (mg) | 100.2 + 21.7 * | 115.5 + 28.9 * | 113.2 + 6.7 * | 117.9 + 27.3 | 124.0 + 25.7 | 129.1 + 14.5 |
| CSA (µm2) | 740.9 ± 99 *** | 948.2 ± 163 *** | 1082.4 ± 80 ***# | 2046.6 ±149 | 2087.9 ± 561 | 2353.8 ± 463 # |
| MM/BM (mg∙g−1) | 0.44 + 0.07 * | 0.50 + 0.10 * | 0.49 + 0.05 * | 0.54 + 0.09 | 0.52 + 0.08 | 0.54 + 0.06 |
| Lo (cm) | 2.9 ± 0.2 | 3.0 ± 0.2 | 2.9 ± 0.1 | 2.9 ± 0.1 | 3.0 ± 0.1 | 2.9 ± 0.1 |
| Pt (mN) | 120.1 ± 46 *** | 278.1 ± 190 *** | 220.7 ± 100 *** | 610.6 ± 263 | 582.6 ± 238 | 493.9 ± 158 |
| TTP (ms) | 51.4 ± 11.9 | 54.6 ± 10.7 | 45.5 ± 13.3 | 46.0 ± 6.3 | 46.3 ± 13.7 | 44.8 ± 10.0 |
| ½ RT (ms) | 82.1 ± 19.9 ** | 78.4 ± 25.9 ** | 53.3 ± 11.8 **¥# | 57.4 ± 8.8 | 56.6 ± 20.1 | 51.4 ± 13.1 ¥# |
| 14 days | ||||||
| n | 7 | 8 | 8 | 7 | 8 | 8 |
| MM (mg) | 139.2 + 21.7 | 124.6 + 25.7 | 150.6 + 12.0¥ | 145.4 + 11.8 | 139.4 + 20.3 | 153.9 + 8.8¥ |
| CSA (µm2) | 1476.5 ± 311 *** | 1648.8 ± 205 *** | 1627.6 ± 261 *** | 2287.1 ± 222 | 2433.9 ± 385 | 2497.7 ± 319 |
| MM/BM (mg∙g−1) | 0.52 + 0.08 | 0.50 + 0.07 | 0.58 + 0.07¥# | 0.56 + 0.04 | 0.56 + 0.07 | 0.60 + 0.04¥# |
| Lo (cm) | 3.1 ± 0.1 | 3.1 ± 0.1 | 3.1 ± 0.1 | 3.2 ± 0.3 | 3.1 ± 0.1 | 3.1 ± 0.2 |
| Pt (mN) | 452.2 ± 57 *** | 383.9 ± 133 *** | 562.9 ± 69 ***¥ | 713.2 ± 230 | 668.5 ± 168 | 795.6 ± 69 ¥ |
| TTP (ms) | 51.7 ± 15.5 * | 47.1 ± 10.9 * | 45.1 ± 7.9 * | 46.0 ± 6.9 | 38.4 ± 9.1 | 38.6 ± 4.0 |
| ½ RT (ms) | 102.9 ± 66.8 ** | 68.5 ± 14.6 **# | 59.2 ± 12.7 **# | 56.7 ± 8.1 | 45.8 ± 9.6 # | 46.5 ± 4.5 # |
| Treatment | CON-INJ | WP-INJ | CR-INJ | CON-NORM | WP-NORM | CR-NORM |
|---|---|---|---|---|---|---|
| Duration | ||||||
| 7 days | ||||||
| n | 8 | 8 | 7 | 8 | 8 | 7 |
| protein (mg/g) | 116.3 ± 33.8 * | 137.5 ± 38.5 * | 142.9 ± 47.1 * | 170.0 ± 33.8 | 163.8 ± 70.3 | 161.4 ± 43.7 |
| contractile protein (mg/g) | 30.6 ± 10.3 ** | 32.9 ± 10.7 ** | 41.9 ± 13.6 ** | 46.9 ± 17.2 | 43.8 ± 12.4 | 52.4 ± 8.6 |
| % Contractile protein | 28.3 ± 11.7 | 24.0 ± 4.5 | 30.7 ± 9.4 | 29.4 ± 12.9 | 32.5 ± 19.3 | 34.4 ± 10.2 |
| 14 days | ||||||
| n | 7 | 8 | 8 | 7 | 8 | 8 |
| protein (mg/g) | 127.1 ± 40.3 * | 127.5 ± 41.3 * | 156.3 ± 29.7 * | 167.1 ± 35.9 | 145.0 ± 36.7 | 170.0 ± 20.0 |
| contractile protein (mg/g) | 32.4 ± 13.0 | 38.1 ± 12.8 | 46.6 ± 15.3 #¥ | 42.1 ± 11.4 | 36.3 ± 9.9 | 54.3 ± 14.1 #¥ |
| % Contractile protein | 25.3 ± 4.8 | 31.7 ± 10.7 | 29.3 ± 5.8 | 25.1 ± 5.9 | 25.9 ± 8.8 | 32.5 ± 9.7 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cooke, M.B.; Rybalka, E.; Stathis, C.G.; Hayes, A. Myoprotective Potential of Creatine Is Greater than Whey Protein after Chemically-Induced Damage in Rat Skeletal Muscle. Nutrients 2018, 10, 553. https://doi.org/10.3390/nu10050553
Cooke MB, Rybalka E, Stathis CG, Hayes A. Myoprotective Potential of Creatine Is Greater than Whey Protein after Chemically-Induced Damage in Rat Skeletal Muscle. Nutrients. 2018; 10(5):553. https://doi.org/10.3390/nu10050553
Chicago/Turabian StyleCooke, Matthew B., Emma Rybalka, Christos G. Stathis, and Alan Hayes. 2018. "Myoprotective Potential of Creatine Is Greater than Whey Protein after Chemically-Induced Damage in Rat Skeletal Muscle" Nutrients 10, no. 5: 553. https://doi.org/10.3390/nu10050553
APA StyleCooke, M. B., Rybalka, E., Stathis, C. G., & Hayes, A. (2018). Myoprotective Potential of Creatine Is Greater than Whey Protein after Chemically-Induced Damage in Rat Skeletal Muscle. Nutrients, 10(5), 553. https://doi.org/10.3390/nu10050553