Lactobacillus plantarum Strain Ln4 Attenuates Diet-Induced Obesity, Insulin Resistance, and Changes in Hepatic mRNA Levels Associated with Glucose and Lipid Metabolism

Abstract
:1. Introduction
2. Materials & Method
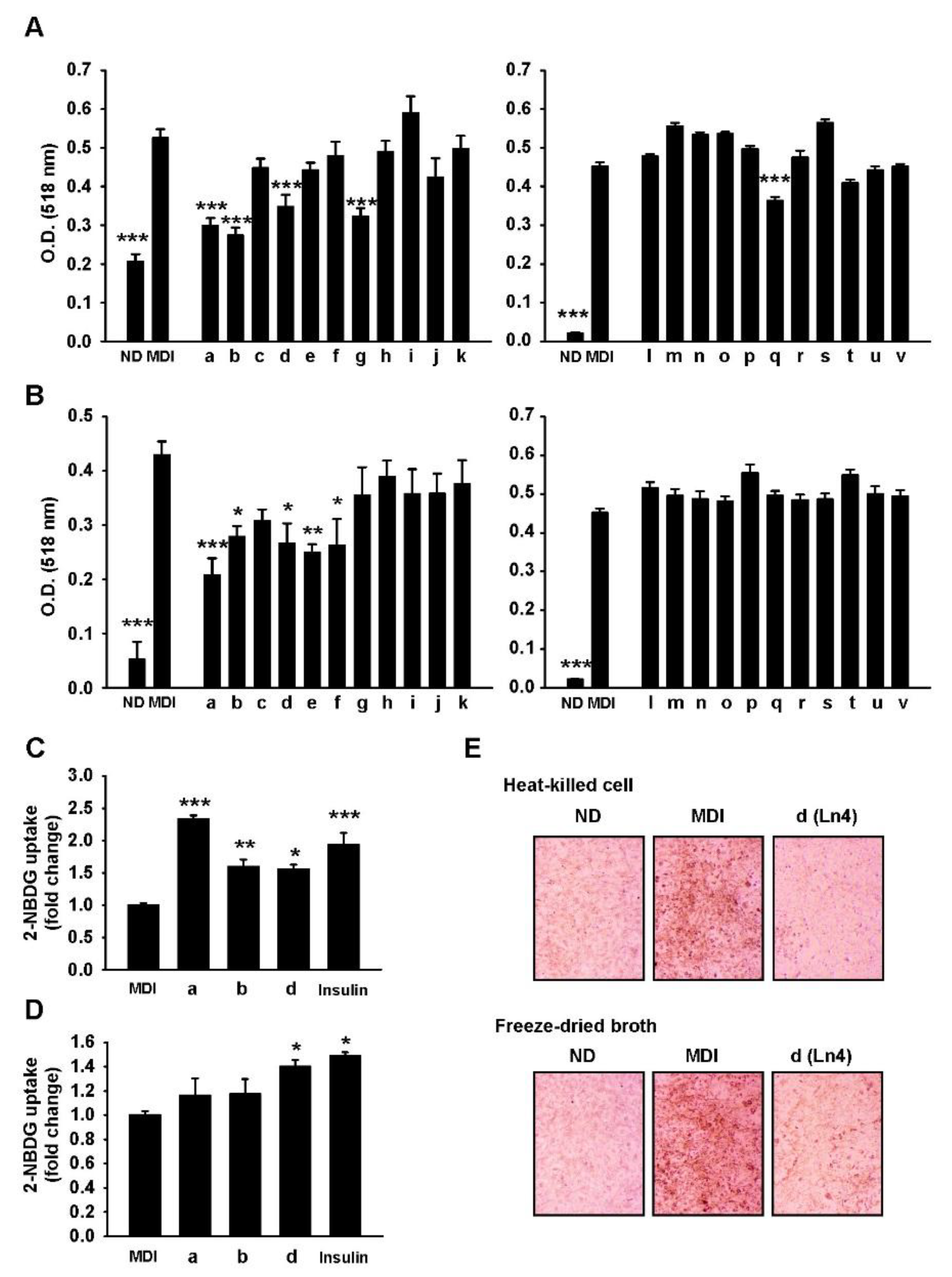
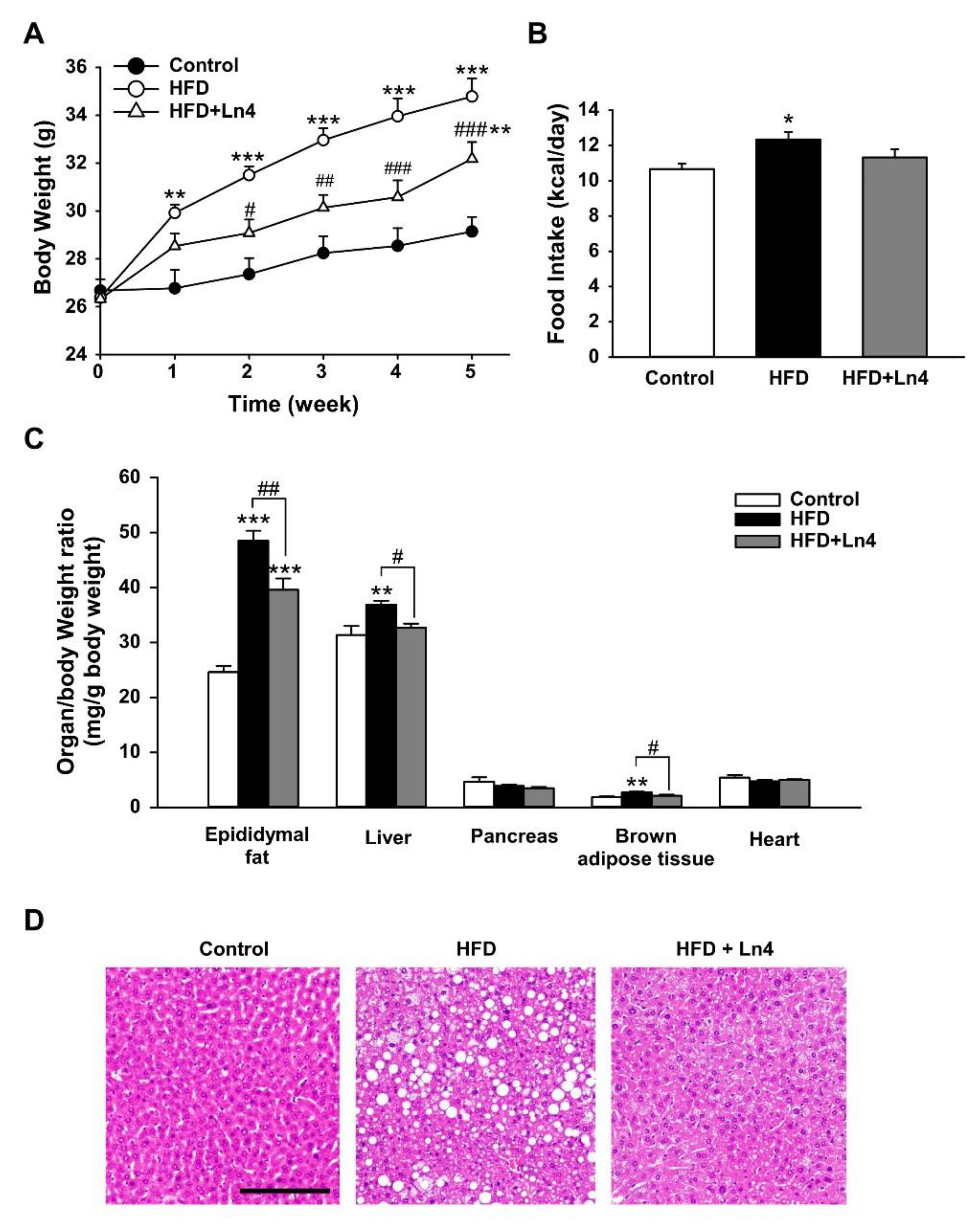
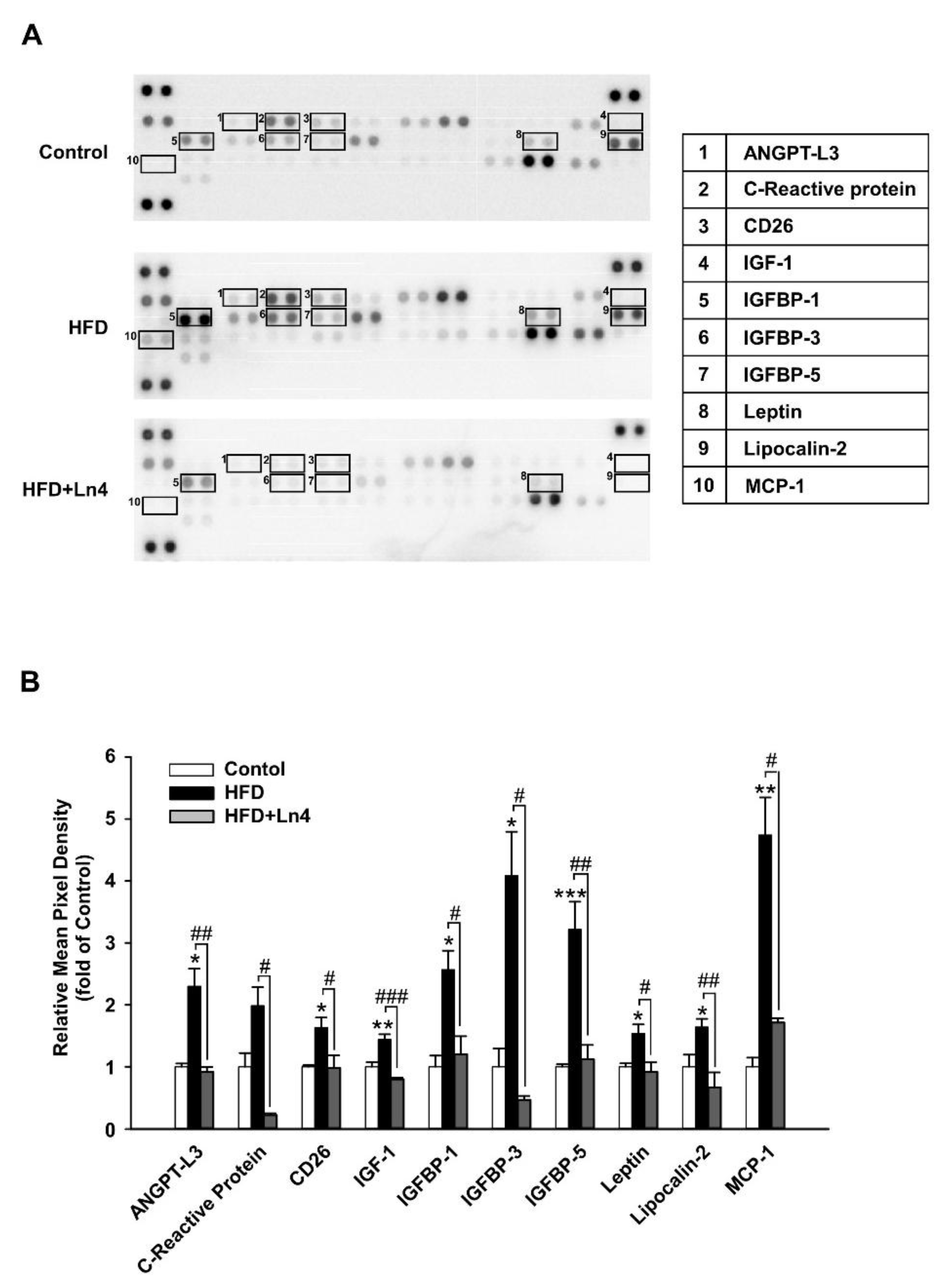
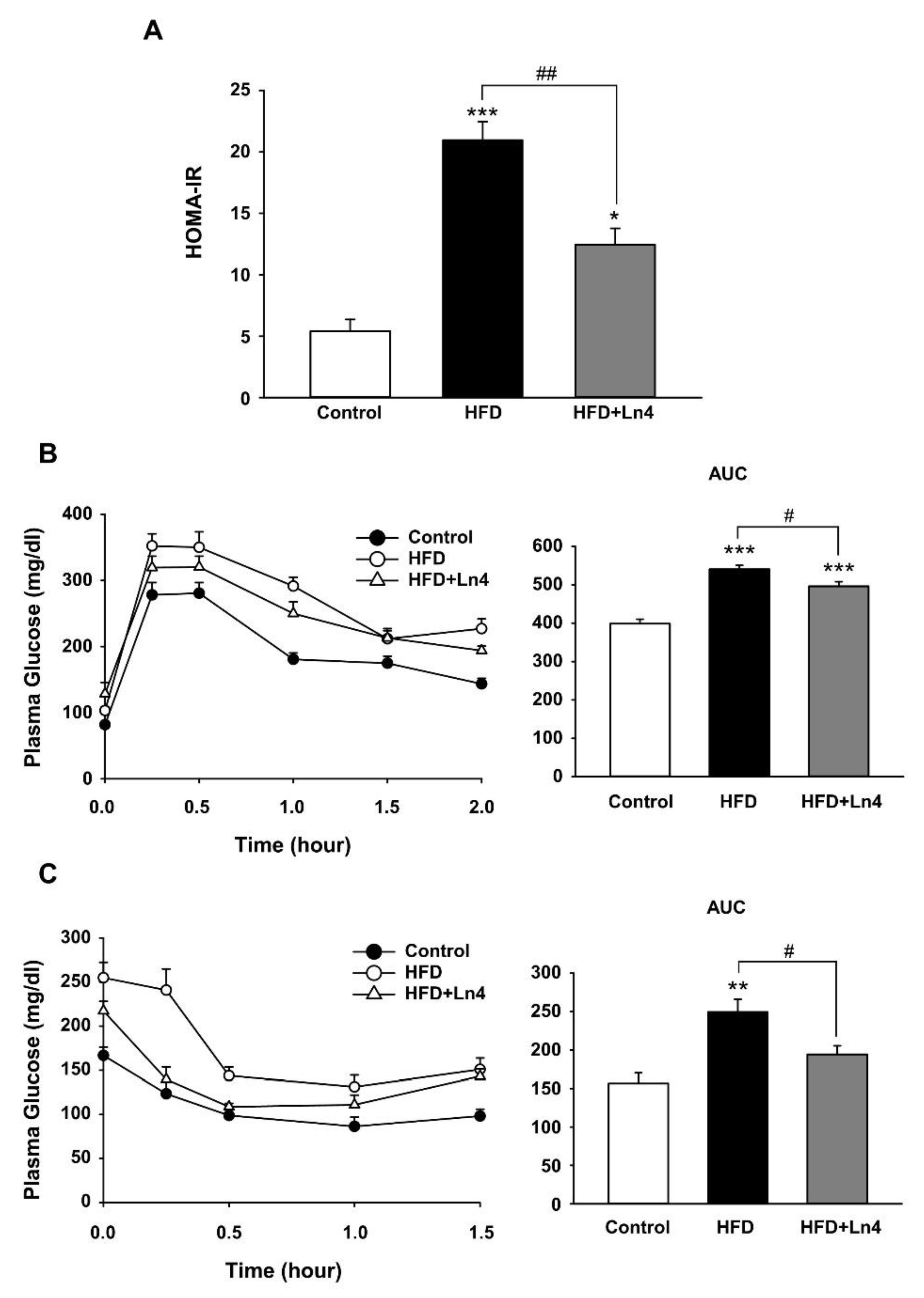
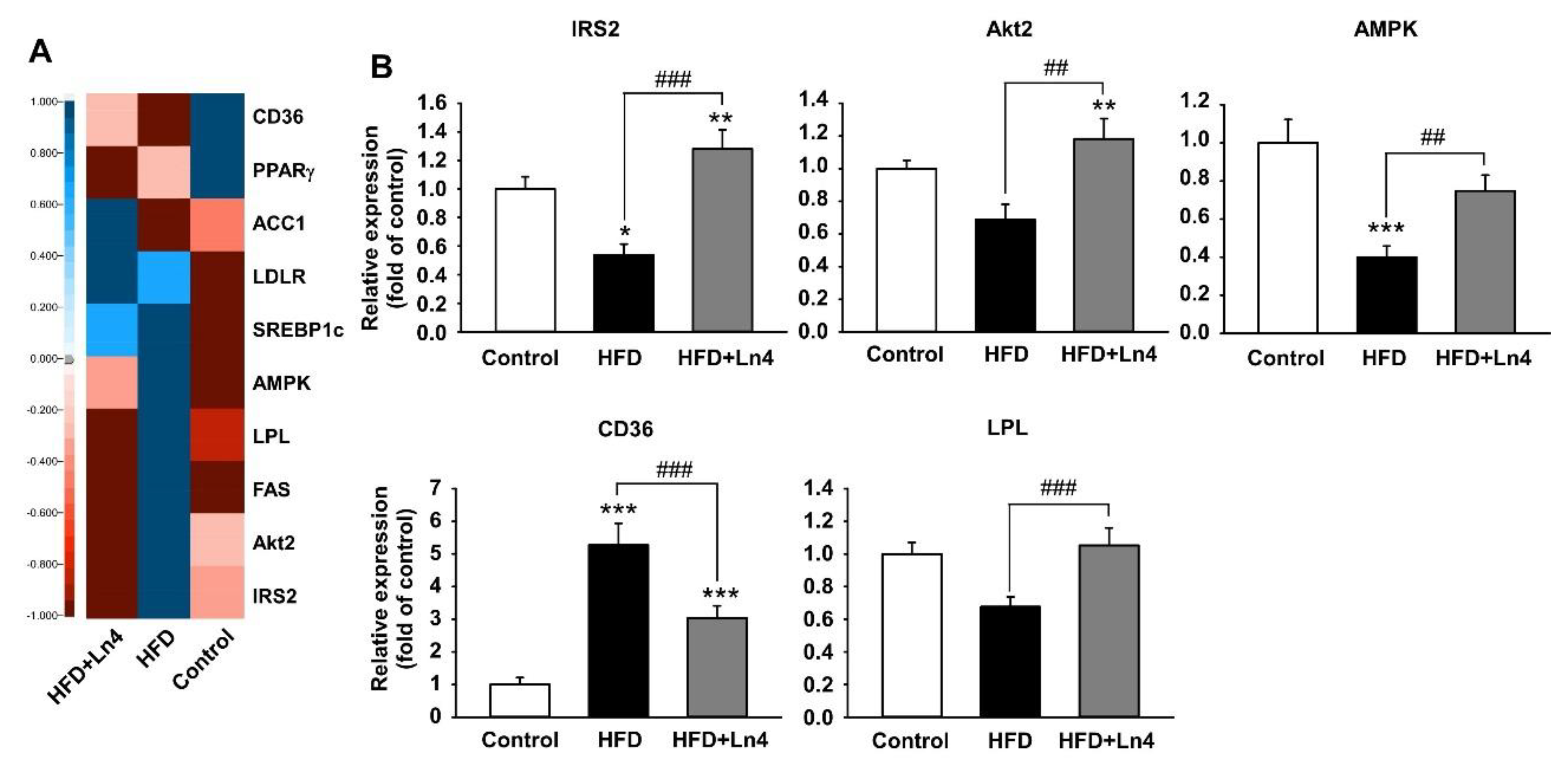
3. Results
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Arroyo-Johnson, C.; Mincey, K.D. Obesity epidemiology worldwide. Gastroenterol. Clin. N. Am. 2016, 45, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, E.J.; LeRoith, D. Obesity and diabetes: The increased risk of cancer and cancer-related mortality. Physiol. Rev. 2015, 95, 727–748. [Google Scholar] [CrossRef] [PubMed]
- Bastien, M.; Poirier, P.; Lemieux, I.; Despres, J.P. Overview of epidemiology and contribution of obesity to cardiovascular disease. Prog. Cardiovasc. Dis. 2014, 56, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Samuel, V.T.; Shulman, G.I. Mechanisms for insulin resistance: Common threads and missing links. Cell 2012, 148, 852–871. [Google Scholar] [CrossRef] [PubMed]
- Muoio, D.M.; Newgard, C.B. Mechanisms of disease:Molecular and metabolic mechanisms of insulin resistance and beta-cell failure in type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Alberti, K.G.; Zimmet, P.; Shaw, J. International diabetes federation: A consensus on type 2 diabetes prevention. Diabet. Med. 2007, 24, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claesson, M.J.; Jeffery, I.B.; Conde, S.; Power, S.E.; O’Connor, E.M.; Cusack, S.; Harris, H.M.; Coakley, M.; Lakshminarayanan, B.; O’Sullivan, O.; et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012, 488, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The effect of diet on the human gut microbiome: A metagenomic analysis in humanized gnotobiotic mice. Sci. Transl. Med. 2009, 1, 6ra14. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delzenne, N.M.; Neyrinck, A.M.; Backhed, F.; Cani, P.D. Targeting gut microbiota in obesity: Effects of prebiotics and probiotics. Nat. Rev. Endocrinol. 2011, 7, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Salas-Salvado, J.; Guasch-Ferre, M.; Diaz-Lopez, A.; Babio, N. Yogurt and diabetes: Overview of recent observational studies. J. Nutr. 2017, 147, 1452S–1461S. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; An, S.Y.; Lee, M.S.; Kim, T.H.; Lee, H.K.; Hwang, W.S.; Choe, S.J.; Kim, T.Y.; Han, S.J.; Kim, H.J.; et al. Fermented kimchi reduces body weight and improves metabolic parameters in overweight and obese patients. Nutr. Res. 2011, 31, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Parvez, S.; Malik, K.A.; Ah Kang, S.; Kim, H.Y. Probiotics and their fermented food products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.Y.; Daily, J.W., 3rd; Kim, H.J.; Park, S. Antidiabetic effects of fermented soybean products on type 2 diabetes. Nutr. Res. 2010, 30, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.H.; Ahn, I.S.; Kim, S.O.; Kong, C.S.; Chung, H.Y.; Do, M.S.; Park, K.Y. Anti-obesity and hypolipidemic effects of black soybean anthocyanins. J. Med. Food 2007, 10, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Kadooka, Y.; Sato, M.; Ogawa, A.; Miyoshi, M.; Uenishi, H.; Ogawa, H.; Ikuyama, K.; Kagoshima, M.; Tsuchida, T. Effect of Lactobacillus gasseri SBT2055 in fermented milk on abdominal adiposity in adults in a randomised controlled trial. Br. J. Nutr. 2013, 110, 1696–1703. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Lee, S.R.; Hwang, K.T.; Ji, G.E. The anti-obesity effects of the dietary combination of fermented red ginseng with levan in high fat diet mouse model. Phytother. Res. 2014, 28, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Jeon, B.S.; Park, J.W.; Kim, B.K.; Kim, H.K.; Jung, T.S.; Hahm, J.R.; Kim, D.R.; Cho, Y.S.; Cha, J.Y. Fermented mushroom milk-supplemented dietary fibre prevents the onset of obesity and hypertriglyceridaemia in otsuka long-evans tokushima fatty rats. Diabetes Obes. Metab. 2005, 7, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.H.; Noh, J.S.; Han, J.S.; Kim, H.J.; Han, E.S.; Song, Y.O. Kimchi, a fermented vegetable, improves serum lipid profiles in healthy young adults: Randomized clinical trial. J. Med. Food 2013, 16, 223–229. [Google Scholar] [CrossRef] [PubMed]
- An, S.Y.; Lee, M.S.; Jeon, J.Y.; Ha, E.S.; Kim, T.H.; Yoon, J.Y.; Ok, C.O.; Lee, H.K.; Hwang, W.S.; Choe, S.J.; et al. Beneficial effects of fresh and fermented kimchi in prediabetic individuals. Ann. Nutr. Metab. 2013, 63, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Lee, S.H.; Kim, J.M.; Park, M.S.; Bae, J.W.; Hahn, Y.; Madsen, E.L.; Jeon, C.O. Metagenomic analysis of kimchi, a traditional korean fermented food. Appl. Environ. Microbiol. 2011, 77, 2264–2274. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.T.; Cheng, P.C.; Pan, T.M. Anti-obesity effects of gut microbiota are associated with lactic acid bacteria. Appl. Microbiol. Biotechnol. 2014, 98, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Moon, P.D.; Kim, S.J.; Choi, I.Y.; An, H.J.; Myung, N.Y.; Jeong, H.J.; Um, J.Y.; Hong, S.H.; Kim, H.M. Lipid profile lowering effect of soypro fermented with lactic acid bacteria isolated from kimchi in high-fat diet-induced obese rats. Biofactors 2008, 33, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Park, J.A.; Tirupathi Pichiah, P.; Yu, J.J.; Oh, S.H.; Daily, J.; Cha, Y.S. Anti-obesity effect of kimchi fermented with weissella koreensis OK1-6 as starter in high-fat diet-induced obese C57BL/6J mice. J. Appl. Microbiol. 2012, 113, 1507–1516. [Google Scholar] [CrossRef] [PubMed]
- Park, K.-Y.; Jeong, J.-K.; Lee, Y.-E.; Daily, J.W., III. Health benefits of kimchi (korean fermented vegetables) as a probiotic food. J. Med. Food 2014, 17, 6–20. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-H.; Song, J.-L.; Park, E.-S.; Ju, J.; Kim, H.-Y.; Park, K.-Y. Anti-obesity effects of starter fermented kimchi on 3t3-l1 adipocytes. Preventive Nutr. Food Sci. 2015, 20, 298. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Oh, S.H.; Cha, Y.S. Lactobacillus brevis OPK-3 isolated from kimchi inhibits adipogenesis and exerts anti-inflammation in 3T3-L1 adipocyte. J. Sci. Food Agric. 2014, 94, 2514–2520. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ojeda, F.J.; Rupérez, A.I.; Gomez-Llorente, C.; Gil, A.; Aguilera, C.M. Cell models and their application for studying adipogenic differentiation in relation to obesity: A review. Int. J. Mol. Sci. 2016, 17, 1040. [Google Scholar] [CrossRef] [PubMed]
- Dunne, C.; O’Mahony, L.; Murphy, L.; Thornton, G.; Morrissey, D.; O’Halloran, S.; Feeney, M.; Flynn, S.; Fitzgerald, G.; Daly, C.; et al. In vitro selection criteria for probiotic bacteria of human origin: Correlation with in vivo findings. Am. J. Clin. Nutr. 2001, 73, 386S–392S. [Google Scholar] [CrossRef] [PubMed]
- Son, S.-H.; Jeon, H.-L.; Jeon, E.B.; Lee, N.-K.; Park, Y.-S.; Kang, D.-K.; Paik, H.-D. Potential probiotic lactobacillus plantarum Ln4 from kimchi: Evaluation of β-galactosidase and antioxidant activities. LWT Food Sci. Technol. 2017, 85, 181–186. [Google Scholar] [CrossRef]
- Seo, S.G.; Yang, H.; Shin, S.H.; Min, S.; Kim, Y.A.; Yu, J.G.; Lee, D.E.; Chung, M.Y.; Heo, Y.S.; Kwon, J.Y.; et al. A metabolite of daidzein, 6,7,4′-trihydroxyisoflavone, suppresses adipogenesis in 3T3-L1 preadipocytes via ATP-competitive inhibition of PI3K. Mol. Nutr. Food Res. 2013, 57, 1446–1455. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Choi, M.-S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed]
- Kadooka, Y.; Sato, M.; Imaizumi, K.; Ogawa, A.; Ikuyama, K.; Akai, Y.; Okano, M.; Kagoshima, M.; Tsuchida, T. Regulation of abdominal adiposity by probiotics (Lactobacillus gasseri SBT2055) in adults with obese tendencies in a randomized controlled trial. Eur. J. Clin. Nutr. 2010, 64, 636–643. [Google Scholar] [CrossRef] [PubMed]
- Bordalo Tonucci, L.; Dos Santos, K.M.; De Luces Fortes Ferreira, C.L.; Ribeiro, S.M.; De Oliveira, L.L.; Martino, H.S. Gut microbiota and probiotics: Focus on diabetes mellitus. Crit. Rev. Food Sci. Nutr. 2017, 57, 2296–2309. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.A. The probiotic paradox: Live and dead cells are biological response modifiers. Nutr. Res. Rev. 2010, 23, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, F.C.; Lan, C.C.; Huang, T.Y.; Chen, K.W.; Chai, C.Y.; Chen, W.T.; Fang, A.H.; Chen, Y.H.; Wu, C.S. Heat-killed and live lactobacillus reuteri GMNL-263 exhibit similar effects on improving metabolic functions in high-fat diet-induced obese rats. Food Funct. 2016, 7, 2374–2388. [Google Scholar] [CrossRef] [PubMed]
- Chuang, L.; Wu, K.G.; Pai, C.; Hsieh, P.S.; Tsai, J.J.; Yen, J.H.; Lin, M.Y. Heat-killed cells of lactobacilli skew the immune response toward T helper 1 polarization in mouse splenocytes and dendritic cell-treated T cells. J. Agric. Food Chem. 2007, 55, 11080–11086. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Weng, W.L.; Lai, W.L.; Tsai, H.P.; Liu, W.H.; Lee, M.H.; Tsai, Y.C. Effect of lactobacillus plantarum strain K21 on high-fat diet-fed obese mice. Evid. Based Complement. Alternat. Med. 2015, 2015, 391767. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Jeon, G. The inhibitory effect of lactobacillus sakei kbl isolated from kimchi on the adipogenesis of 3T3-L1 cells. Afr. J. Biotechnol. 2015, 14, 2611–2615. [Google Scholar]
- Kim, H.S.; Ali, O.; Shim, M.; Lee, K.W.; Vuguin, P.; Muzumdar, R.; Barzilai, N.; Cohen, P. Insulin-like growth factor binding protein-3 induces insulin resistance in adipocytes in vitro and in rats in vivo. Pediatr. Res. 2007, 61, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Motaghedi, R.; Gujral, S.; Sinha, S.; Ranchandani, N.; Ten, S.; Sison, M.C.I.; Maclaren, N.K. Igfbp-3 is increased by hyperinsulinemia in insulin resistant, obese children. Diabetes 2004, 53, A414–A415. [Google Scholar]
- Yan, Q.-W.; Yang, Q.; Mody, N.; Graham, T.E.; Hsu, C.-H.; Xu, Z.; Houstis, N.E.; Kahn, B.B.; Rosen, E.D. The adipokine lipocalin 2 is regulated by obesity and promotes insulin resistance. Diabetes 2007, 56, 2533–2540. [Google Scholar] [CrossRef] [PubMed]
- Silha, J.V.; Krsek, M.; Skrha, J.V.; Sucharda, P.; Nyomba, B.; Murphy, L.J. Plasma resistin, adiponectin and leptin levels in lean and obese subjects: Correlations with insulin resistance. Eur. J. Endocrinol. 2003, 149, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Kanda, H.; Tateya, S.; Tamori, Y.; Kotani, K.; Hiasa, K.-I.; Kitazawa, R.; Kitazawa, S.; Miyachi, H.; Maeda, S.; Egashira, K. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J. Clin. Investig. 2006, 116, 1494–1505. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (mouse) | Primer Sequence | Product Size (bp) |
|---|---|---|
| SREBP1c | F: 5’- GATGTGCGAACTGGACA-3’ R: 5’-CATAGGGGGCGTCAAACAG-3’ | 104 |
| ACC1 | F: 5’-CCTCCGTCAGCTCAGATACA-3’ R: 5’-TTTACTAGGTGCAAGCCAGACA-3’ | 103 |
| FAS | F: 5’-AGGGGTCGACCTGGTCCTCA-3’ R: 5’-GCCATGCCCAGAGGGTGGTT-3’ | 132 |
| PPARγ | F: 5’-TTGATTTCTCCAGCATTTCT-3’ R: 5’-TGTTGTAGAGCTGGGTCTTT-3’ | 172 |
| LDLR | F: 5’-TGACTCAGACGAACAAGGCTG-3’ R: 5’-ATCTAGGCAATCTCGGTCTCC-3’ | 118 |
| CD36 | F: 5’-CAGATGACGTGGCAAAGAAC-3’ R: 5’-TGGCTCCATTGGGCTGTA-3’ | 144 |
| AMPK | F: 5’-AAGATCGGACACTACGTCCTG-3’ R: 5’-TGCCACTTTATGGCCTGTCAA-3’ | 96 |
| Akt2 | F: 5’-ACGTGGTGAATACATCAAGACC-3’ R: 5’-ACCCAATGAAAGATCCATCACTC-3’ | 71 |
| IRS2 | F: 5’-TCTACACCCGAGACGAACACT-3’ R: 5’-TGGGCCTTTGCCCGATTATG-3’ | 103 |
| LPL | F: 5’-TTGCCCTAAGGACCCCTGAA-3’ R: 5’-ACAGAGTCTGCTAATCCAGGAAT-3’ | 70 |
| β-actin | F: 5’-TGTCCACCTTCCAGCAGATGT-3’ R: 5’-AGCTCAGTAACAGTCCGCCTAGA-3’ | 101 |
| Control | HFD | HFD+Ln4 | |
|---|---|---|---|
| 5-h Fasted Glucose (mg/dL) | 155.2 ± 13.8 | 230.5 ± 9.3 *** | 199.2 ± 3.8 * |
| 5-h Fasted Insulin (μU/mL) | 18.8 ± 2.8 | 38.1 ± 1.9 *** | 28.4 ± 3.4 # |
| Total TG (mg/dL) | 158.2 ± 6.1 | 203.9 ± 11.1 ** | 150.8 ± 11.1 ## |
| HDL-Cholesterol (mg/dL) | 49.2 ± 5.4 | 79.5 ± 2.0 *** | 74.2 ± 2.7 *** |
| LDL-Cholesterol (mg/dL) | 21.0 ± 1.9 | 29.6 ± 1.4 ** | 30.5 ± 2.1 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, E.; Jung, S.-R.; Lee, S.-Y.; Lee, N.-K.; Paik, H.-D.; Lim, S.-I. Lactobacillus plantarum Strain Ln4 Attenuates Diet-Induced Obesity, Insulin Resistance, and Changes in Hepatic mRNA Levels Associated with Glucose and Lipid Metabolism. Nutrients 2018, 10, 643. https://doi.org/10.3390/nu10050643
Lee E, Jung S-R, Lee S-Y, Lee N-K, Paik H-D, Lim S-I. Lactobacillus plantarum Strain Ln4 Attenuates Diet-Induced Obesity, Insulin Resistance, and Changes in Hepatic mRNA Levels Associated with Glucose and Lipid Metabolism. Nutrients. 2018; 10(5):643. https://doi.org/10.3390/nu10050643
Chicago/Turabian StyleLee, Eunjung, So-Ra Jung, So-Young Lee, Na-Kyoung Lee, Hyun-Dong Paik, and Seong-Il Lim. 2018. "Lactobacillus plantarum Strain Ln4 Attenuates Diet-Induced Obesity, Insulin Resistance, and Changes in Hepatic mRNA Levels Associated with Glucose and Lipid Metabolism" Nutrients 10, no. 5: 643. https://doi.org/10.3390/nu10050643
APA StyleLee, E., Jung, S. -R., Lee, S. -Y., Lee, N. -K., Paik, H. -D., & Lim, S. -I. (2018). Lactobacillus plantarum Strain Ln4 Attenuates Diet-Induced Obesity, Insulin Resistance, and Changes in Hepatic mRNA Levels Associated with Glucose and Lipid Metabolism. Nutrients, 10(5), 643. https://doi.org/10.3390/nu10050643