Association of Tea Consumption with Risk of Alzheimer’s Disease and Anti-Beta-Amyloid Effects of Tea

 ,
,  and
and
Abstract
:1. Introduction
2. Epidemiological Evidence
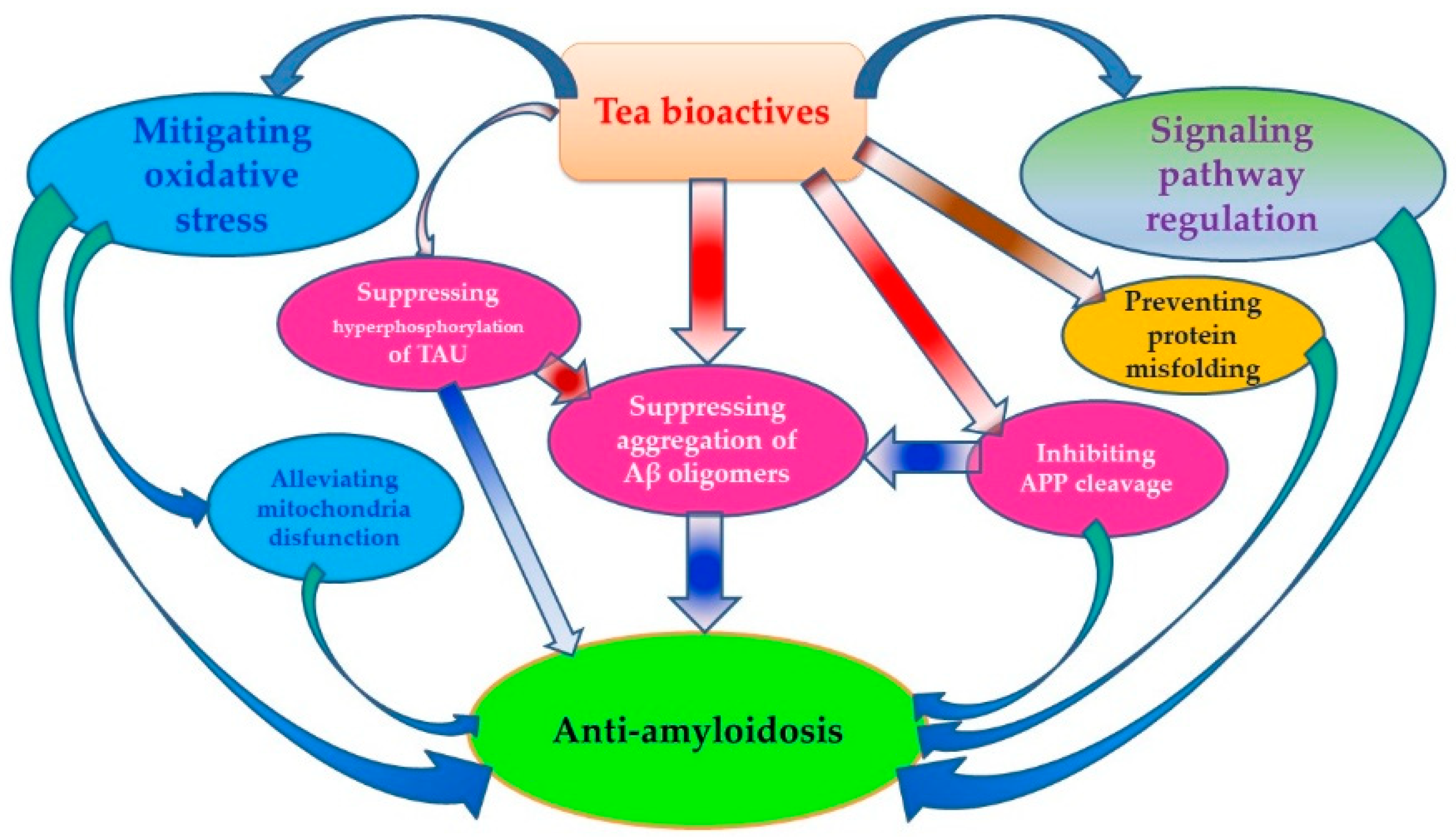
3. Anti-Aβ Effects of Tea
3.1. Inhibiting APP Cleavage by Regulating Activity of Related Enzymes
3.2. Preventing Protein Misfolding and Aβ-Induced Membrane Damages
3.3. Mitigating Aβ-Induced Oxidative Stress
3.4. Suppressing Aggregation of Aβ Oligomers and Formation of Aβ Fibrils
3.5. Regulating Signaling Pathways Involving Aβ Generation
3.6. Alleviating Aβ-Induced Mitochondria Disfunction
3.7. Inhibiting Hyperphosphorylation of TAU Protein
4. Conclusion and Future Challenges
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kou, X.; Chen, N. Resveratrol as a natural autophagy regulator for prevention and treatment of Alzheimer’s disease. Nutrients 2017, 9, 927. [Google Scholar] [CrossRef]
- Swaminathan, N. How to save your brain. Psychol. Today 2012, 45, 74–79. [Google Scholar]
- Geiser, R.J.; Chastain, S.E.; Moss, M.A. Regulation of Bace1 mRNA expression in Alzheimer’s disease by green tea catechins and black tea theaflavins. Biophys. J. 2017, 112 (Suppl. 3), 362a. [Google Scholar] [CrossRef]
- Hwang, S.; Lim, J.W.; Kim, H. Inhibitory effect of lycopene on amyloid-β-induced apoptosis in neuronal cells. Nutrients 2017, 9, 883. [Google Scholar] [CrossRef] [PubMed]
- Ribarič, S. Peptides as potential therapeutics for Alzheimer’s disease. Molecules 2018, 23, 283. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.; Zhao, Q.; Guo, Q.; Meng, H.; Wang, B.; Yu, P.; Luo, J.; Zhou, Y.; Yu, L.; Zheng, L.; Chu, S.; et al. The Shanghai aging study: Study design, baseline characteristics, and prevalence of dementia. Neuroepidemiology 2014, 43, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Javier, O.P.; Carmelo, P.V. Dietary supplements for cognitive impairment. Actas Esp. Psiquiatr. 2017, 45, 37–47. [Google Scholar]
- Rubio-Perez, J.M.; Albaladeo, M.D.; Zafrilla, P.; Vidal-Guevara, M.L.; Morillas-Ruiz, J.M. Effects of an antioxidant beverage on biomarkers of oxidative stress in Alzheimer’s patients. Eur. J. Nutr. 2016, 55, 2105–2116. [Google Scholar] [CrossRef] [PubMed]
- Kesse-Guyot, E.; Fezeu, L.; Andreeva, V.A.; Touvier, M.; Scalbert, A.; Hercberg, S.; Galan, P. Total and specific polyphenol intakes in midlife are associated with cognitive function measured 13 years later. J. Nutr. 2012, 142, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Chong, M.S.; Lim, W.S.; Lee, T.S.; Kua, E.H.; Ng, T.P. Tea for Alzheimer prevention. J. Prev. Alzheimer Dis. 2015, 2, 136–141. [Google Scholar]
- Liu, K.; Liang, X.; Kuang, W. Tea consumption maybe an effective active treatment for adult attention deficit hyperactivity disorder (ADHD). Med. Hypotheses 2011, 76, 461–463. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, O.; Amit, T.; Mandel, S.; Youdim, M.B.H. Neuroprotective molecular mechanisms of (−)-epigallocatechin-3-gallate: A reflective outcome of its antioxidant, iron chelating and neuritogenic properties. Genes Nutr. 2009, 4, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.A.; Mandal, A.K.A.; Khan, Z.A. Potential neuroprotective properties of epigallocatechin-3-gallate (EGCG). Nutr. J. 2016, 15, 60. [Google Scholar] [CrossRef] [PubMed]
- Fernando, W.; Somaratne, G.; Goozee, K.G.; Williams, S.; Singh, H.; Martins, R.N. Diabetes and Alzheimer’s Disease: Can Tea Phytochemicals Play a Role in Prevention? J. Alzheimer Dis. 2017, 59, 481–501. [Google Scholar] [CrossRef] [PubMed]
- Rusted, J.; Sheppard, L. Action-based memory in Alzheimer’s disease: A longitudinal look at tea making. Neurocase 2002, 8, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Kuriyama, S.; Hozawa, A.; Ohmori, K.; Shimazu, T.; Matsui, T.; Ebihara, S.; Awata, S.; Nagatomi, R.; Arai, H.; Tsuji, I. Green tea consumption and cognitive function: A cross-sectional study from the Tsurugaya Project. Am. J. Clin. Nutr. 2006, 83, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Tomata, Y.; Kakizaki, M.; Nakaya, N.; Tsuboya, T.; Sone, T.; Kuriyama, S.; Hozawa, A.; Tsuji, I. Green tea consumption and the risk of incident functional disability in elderly Japanese: The Ohsakicohort 2006 study. Am. J.Clin. Nutr. 2012, 95, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Noguchi-Shinohara, M.; Yuki, S.; Dohmoto, C.; Ikeda, Y.; Samuraki, M.; Iwasa, K.; Yokogawa, M.; Asai, K.; Komai, K.; Nakamura, H.; et al. Consumption of green tea, but not black tea or coffee, is associated with reduced risk of cognitive decline. PLoS ONE 2014, 9, E96013. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, K.; Watanabe, Y.; Nakamura, K.; Sanpei, K.; Wakasugi, M.; Yokoseki, A.; Onodera, O.; Ikeuchi, T.; Kuwano, R.; Momotsu, T.; et al. Modifiable factors associated with cognitive impairment in 1143 Japanese outpatients: The project in Sado for total health (PROST). Dement.Geriatr. Cogn. Disord. Extra 2016, 6, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Ide, K.; Yamada, H.; Takuma, N.; Kawasaki, Y.; Harada, S.; Nakase, J.; Ukawa, Y.; Sagesaka, Y.M. Effects of green tea consumption on cognitive dysfunction in an elderly population: A randomized placebo controlled study. Nutr. J. 2016, 15, 49. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.P.; Feng, L.; Niti, M.; Kua, E.H.; Yap, K.B. Tea consumption and cognitive impairment and decline in older Chinese adults. Am. J. Clin. Nutr. 2008, 88, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Gwee, X.; Kua, E.H.; Ng, T.P. Cognitive function and tea consumption in community dwelling older Chinese in Singapore. J. Nutr. Health Aging 2010, 14, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Li, J.L.; Kua, E.K.; Lee, T.S.; Yap, K.B.; Rush, A.J.; Ng, T.P. Association between tea consumption and depressive symptoms in older Chinese adults. J. Am. Geriatr. Soc. 2012, 60, 2358–2360. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.P.; Yong, P.Z.; Sun, Y.; Mahendran, R.; Wong, J.C.M.; Qiu, C.; Ng, T.P.; Kua, E.H.; Feng, L. Associations of long-term tea consumption with depressive and anxiety symptoms in community-living elderly: Findings from the diet and healthy aging study. J. Prev. Alzheimer Dis. 2018, 5, 21–25. [Google Scholar]
- Huang, C.Q.; Dong, B.R.; Zhang, Y.L.; Wu, H.M.; Liu, Q.X. Association of cognitive impairment with smoking, alcohol consumption, tea consumption, and exercise among Chinese nonagenarians/centenarians. Cogn. Behav. Neurol. 2009, 22, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Huang, Y.; Cheng, H.G. Lower intake of vegetables and legumes associated with cognitive decline among illiterate elderly Chinese: A 3-year cohort study. J. Nutr. Health Aging 2012, 16, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.J.; He, C.H.; Li, S.; Zhang, S.Y.; Duan, S.Y.; Sun, H.P.; Shen, Y.P.; Xu, Y.; Yin, J.Y.; Pan, C.W. Tea consumptionis associated with cognitive impairment in older Chinese adults. Aging Ment. Health 2017. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Yan, Z.R.; Sun, B.L.; Cai, C.Z.; Jiang, H.; Kua, E.H.; Ng, T.P.; Qiu, C.X. Tea consumption and depressive symptoms in older people in rural China. J. Am. Geriatr. Soc. 2013, 61, 1943–1947. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Xiao, Y.; Ying, X.; Li, S.; Zhai, Y.; Shang, X.; Li, F.; Wang, X.; He, F.; Lin, J. Tea consumption and cognitive impairment: A cross-sectional study among Chinese elderly. PLoS ONE 2015, 10, e0137781. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Jin, X.Q.; Yan, J.; Jin, Y.; Yu, W.; Wu, H.B.; Xu, S.H. Prevalence of dementia, cognitive status and associated risk factors among elderly of Zhejiang province, China in 2014. Age Ageing 2016, 45, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Sautter, J.; Gu, D. Associations between frequency of tea consumption and health and mortality: Evidence from old Chinese. Br. J. Nutr. 2012, 108, 1686–1697. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Li, J.; Ng, T.P.; Lee, T.S.; Kua, E.H.; Zeng, Y. Tea drinking and cognitive function in oldest-old Chinese. J. Nutr. Health Aging 2012, 16, 754–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.H.; Cheng, G.W.; Rong, S.; Zhu, H.L.; Chen, D.; Yang, B.; Li, W.F. A cross-sectional study on mild cognitive impairment among the elderly in communities of Huangshi City. Chin. J. Dis. Control Prev. 2017, 21, 767–771. [Google Scholar]
- Wang, Z.; Dong, B.; Zeng, G.; Li, J.; Wang, W.; Wang, B.; Yuan, Q. Is there an association between mild cognitiveimpairment and dietary pattern in Chineseelderly? Results from a cross-sectional population study. BMC Public Health 2010, 10, 595. [Google Scholar] [CrossRef] [PubMed]
- Nurk, E.; Refsum, H.; Drevon, C.A.; Tell, G.S.; Nygaard, H.A.; Engedal, K.; Smith, A.D. Intake of flavonoid-rich wine, tea, and chocolate by elderly men and women is associated with better cognitive test performance. J. Nutr. 2009, 139, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Beking, K.; Vieira, A. Flavonoid intake and disability-adjusted life years due to Alzheimer’s and related dementias: A population-based study involving twenty-three developed countries. Public Health Nutr. 2010, 13, 1403–1409. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.P.; Huang, C.; Cui, Q.Y.; Yang, D.J.; Sun, K.; Chen, X.; Li, X.H. Meta-analysis of the association between tea intake and the risk of cognitive disorders. PLoS ONE 2016, 11, e0165861. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Yoshiike, Y.; Takashima, A.; Hasegawa, K.; Naiki, H.; Yamada, M. Potent anti-amyloidogenic and fibril-destabilizing effects of polyphenols in vitro: Implications for the prevention and therapeutics of Alzheimer’s disease. J. Neurochem. 2003, 87, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Wu, X.K.; Wu, Q.; Gong, D.Z.; Shi, M.J.; Guan, L.L.; Zhang, J.; Liu, J.; Yuan, B.; Han, G.Z.; Zou, Y. Green tea polyphenols protect against okadaic acid-induced acute learning and memory impairments in rats. Nutrition 2014, 30, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Rezai-Zadeh, K.; Arendash, G.W.; Hou, H.Y.; Fernandez, F.; Jensen, M.; Runfeldt, M.; Shytle, R.D.; Tan, J. Green tea epigallocatechin-3-gallate (EGCG) reduces beta-amyloid mediated cognitive impairment and modulates tau pathology in Alzheimer transgenic mice. Brain Res. 2008, 1214, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Rezai-Zadeh, K.; Shytle, D.; Sun, N.; Mori, T.; Hou, H.Y.; Jeanniton, D.; Ehrhart, J.; Townsend, K.; Zeng, J.; Morgan, D.; Hardy, J.; et al. Green Tea epigallocatechin-3-gallate (EGCG) modulates amyloid precursor protein cleavage and reduces cerebral amyloidosis in Alzheimer transgenic mice. J. Neurosci. 2005, 25, 8807–8814. [Google Scholar] [CrossRef] [PubMed]
- Omar, S.H. Biophenols pharmacology against the amyloidogenic activity in Alzheimer’s disease. Biomed. Pharmacother. 2017, 89, 396–413. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.X.; Li, Y.B.; Zhao, R.P. Epigallocatechingallate attenuates beta-amyloid generation and oxidative stress involvement of PPAR gamma in N2a/APP695 cells. Neurochem. Res. 2017, 42, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Obregon, D.F.; Rezai-Zadeh, K.; Bai, Y.; Sun, N.; Hou, H.Y.; Ehrhart, J.; Zeng, J.; Mori, T.; Arendash, G.W.; Shytle, D.; et al. ADAM10 activation is required for green tea (−)-epigallocatechin-3-gallate-induced α-secretase cleavage of amyloid precursor protein. J. Biol. Chem. 2006, 281, 16419–16427. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.W.; Rezai-Zadeh, K.; Obregon, D.; Tan, J. EGCG functions through estrogen receptor-mediated activation of ADAM10 in the promotion of non-amyloidogenic processing of APP. FEBS Lett. 2010, 584, 4259–4267. [Google Scholar] [CrossRef] [PubMed]
- Cox, C.J.; Choudhry, F.; Peacey, E.; Perkinton, M.S.; Richardson, J.C.; Howlett, D.R.; Lichtenthaler, S.F.; Francis, P.T.; Williams, R.J. Dietary (−)-epicatechin as a potent inhibitor of β γ-secretase amyloid precursor protein processing. Neurobiol. Aging 2015, 36, 178–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rennex, D.; Hemmings, B.A.; Hofsteenge, J.; Stone, S.R. cDNA cloning of porcine brain prolylendopeptidase and identification of the active-site seryl residue. Biochemistry 1991, 30, 2195–2203. [Google Scholar] [CrossRef] [PubMed]
- Aoyagi, T.; Nagai, M.; Ogawa, K.; Gojima, F.; Okada, M.; Ikeda, T.; Hamada, M.; Takeuchi, T. Poststatin, a new inhibitor of prolylendopeptidase, produced by streptomycesviridochromogens MH534-30F3. I. Taxonomy, production, isolation, physico-chemical properties and biological activities. J. Antibiot. 1991, 44, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Ishiura, S.; Tsukahara, T.; Tabira, T.; Shimizu, T.; Arahata, K.; Sugita, H. Identification of a putative amyloid A4-generating enzyme as a prolylendopeptidase. FEBS Lett. 1990, 260, 131–134. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, S.I.; Song, K.S. Prolylendopeptidaseinhibitors from green tea. Arch. Pharm. Res. 2001, 24, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Yuksel, M.; Biberoglu, K.; Onder, S.; Akbulut, K.G.; Tacal, O. Effects of phenothiazine-structured compounds on APP processing in Alzheimer’s disease cellular model. Biochimie 2017, 138, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Jamal, Q.M.S.; Shams, S.; Al-Wabel, N.A.; Siddiqui, M.U.; Alzohairy, M.A.; Al Karaawi, M.A.; Kesari, K.K.; Mushtaq, G.; Kamal, M.A. In Silico Analysis of Green Tea Polyphenols as Inhibitors of AChE and BChE Enzymes in Alzheimer’s Disease Treatment. CNS Neurol. Disord.-Drug 2016, 15, 624–628. [Google Scholar] [CrossRef]
- Ayoub, S.; Melzig, M.F. Induction of neutral endopeptidase (NEP) activity of SK-N-SH cells by natural compounds from green tea. J. Pharm. Pharmacol. 2006, 58, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Yuk, D.Y.; Lee, J.W.; Lee, S.Y.; Ha, T.Y.; Oh, K.W.; Yun, Y.P.; Hong, J.T. Epigallocatechin-3-gallate prevents lipopolysaccharide-induced elevation of beta-amyloid generation and memory deficiency. Brain Res. 2009, 1250, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.J.; Warren, F.J.; Netzel, G.; Gidley, M.J. 3 or 3′-Galloyl substitution plays an important role in association of catechins and theaflavins with porcine pancreatic α-amylase: The kinetics of inhibition of α-amylase by tea polyphenols. J. Funct. Foods 2016, 26, 144–156. [Google Scholar] [CrossRef]
- Gauci, A.J.; Caruana, M.; Giese, A.; Scerri, C.; Vassallo, N. Identification of polyphenolic compounds and black tea extract as potent inhibitors of lipid membrane destabilization by Aβ(42) aggregates. J. Alzheimers Dis. 2011, 27, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Hudson, S.A.; Ecroyd, H.; Dehle, F.C.; Musgrave, I.F.; Carver, J. (−)-Epigallocatechin-3-gallate (EGCG) maintains k-casein in its pre-fibrillar state without redirecting its aggregation pathway. J. Mol. Biol. 2009, 392, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.A.; Cheng, X.R.; da Silva, I.S.; Hung, V.W.S.; Veloso, A.J.; Angnes, L.; Kerman, K. Electroanalysis of the interaction between (−)-epigallocatechin-3-gallate (EGCG) and amyloid-beta in the presence of copper. Metallomics 2013, 5, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Palhano, F.L.; Lee, J.; Grimster, N.P.; Kelly, J.W. Toward the molecular mechanism(s) by which EGCG treatment remodels mature amyloid fibrils. J. Am. Chem. Soc. 2013, 135, 7503–7510. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.T.; Jung, C.H.; Lee, S.R.; Bae, J.H.; Baek, W.K.; Suh, M.H.; Park, J.; Park, C.W.; Suh, S.I. The green tea polyphenol (−)-epigallocatechingallate attenuates β-amyloid-induced neurotoxicity in cultured hippocampal neurons. Life Sci. 2001, 70, 603–614. [Google Scholar] [CrossRef]
- Harvey, B.S.; Musgrave, I.F.; Ohlsson, K.S.; Fransson, A.; Smid, S.D. The green tea polyphenol (−)-epigallocatechin-3-gallate inhibits amyloid-β evoked fibril formation and neuronal cell death in vitro. Food Chem. 2011, 129, 1729–1736. [Google Scholar] [CrossRef]
- Avramovich-Tirosh, Y.; Rezrlichenko, D.; Amit, T.; Zheng, H.; Fridkin, M.; Weinreb, O.; Mandel, S.; Youdim, M.B.H. Neurorescue activity, APP regulation and amyloid-β peptide reduction by novel multi-functional brain permeable iron- chelating- antioxidants, m-30 and green tea polyphenol, EGCG. Curr. Alzheimer Res. 2007, 4, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Lee, C.; Park, G.H.; Jang, J.H. Neuroprotective effect of epigallocatechin-3-gallate against β-amyloid-induced oxidative and nitrosative cell death via augmentation of antioxidant defense capacity. Arch. Pharm. Res. 2009, 32, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Shimmyo, Y.; Kihara, T.; Akaike, A.; Niidome, T.; Sugimoto, H. Epigallocatechin-3-gallate and curcumin suppress amyloid beta-induced beta-site APP cleaving enzyme-1 upregulation. Neuroreport 2008, 19, 1329–1333. [Google Scholar] [CrossRef] [PubMed]
- Okello, E.J.; McDougall, G.J.; Kumar, S.; Seal, C.J. In vitro protective effects of colon-available extract of Camellia sinensis (tea) against hydrogen peroxide and beta-amyloid (A beta((1-42))) induced cytotoxicity in differentiated PC12 cells. Phytomedicine 2011, 18, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, E.; Limon, D.; Perez-Severiano, F.; Diaz, A.; Ortega, L.; Zenteno, E.; Guevara, J. Antioxidant effects of epicatechin on the hippocampal toxicity caused by amyloid-beta 25–35 in rats. Eur. J. Pharmacol. 2009, 616, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Arai, T.; Ohno, A.; Mori, K.; Kuwata, H.; Mizuno, M.; Imai, K.; Hara, S.; Shibanuma, M.; Kurihara, M.; Miyata, N.; et al. Inhibition of amyloid fibril formation and cytotoxicity by caffeic acid-conjugated amyloid-beta C-terminal peptides. Bioorg. Med. Chem. Lett. 2016, 26, 5468–5471. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.L.; Chen, T.F.; Chiu, M.J.; Way, T.D.; Lin, J.K. Epigallocatechingallate (EGCG) suppresses beta-amyloid-induced neurotoxicity through inhibiting c-Abl/FE65 nuclear translocation and GSK3 beta activation. Neurobiol. Aging 2009, 30, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Abedini, A.; Plesner, A.; Verchere, C.B.; Raleigh, D.P. The flavanol (−)-epigallocatechin 3-gallate inhibits amyloid formation by islet amyloid polypeptide, disaggregates amyloid fibrils and protects cultured cells against IAPP induced toxicity. Biochemistry 2010, 49, 8127–8133. [Google Scholar] [CrossRef] [PubMed]
- Lopez del Amo, J.M.; Fink, U.; Dasari, M.; Grelle, G.; Wanker, E.E.; Bieschke, J.; Reif, B. Structural properties of EGCG-induced, nontoxic Alzheimer’s disease A β oligomers. J. Mol. Biol. 2012, 421, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.; VanSchouwen, B.; Jafari, N.; Ni, X.D.; Ortega, J.; Melacini, G. Molecular mechanism for the (−)-epigallocatechingallate-induced toxic to nontoxic remodeling of A beta oligomers. J. Am. Chem. Soc. 2017, 139, 13720–13734. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.H.; Liu, F.F.; Dong, X.Y.; Sun, Y. Thermodynamic analysis of the molecular interactions between amyloid β-peptide 42 and (−)-epigallocatechin-3-gallate. J. Phys. Chem. B 2010, 114, 11576–11583. [Google Scholar] [CrossRef] [PubMed]
- Jo, M.; Park, M.H.; Choi, D.Y.; Yuk, D.Y.; Lee, Y.M.; Lee, J.M.; Jeong, J.H.; Oh, K.W.; Lee, M.S.; Han, S.B.; et al. Neuroprotective effect of L-theanine on A β-induced neurotoxicity through anti-oxidative mechanisms in SK-N-SH and SK-N-MC cells. Biomol. Ther. 2011, 19, 288–295. [Google Scholar] [CrossRef]
- Chastain, S.E.; Moss, M. Green and black tea polyphenols mechanistically inhibit the aggregation of amyloid-β in Alzheimer’s disease. Biophys. J. 2015, 108, 357a. [Google Scholar] [CrossRef]
- Sinha, S.; Du, Z.M.; Maiti, P.; Klärner, F.G.; Schrader, T.; Wang, C.Y.; Bitan, G. Comparison of three amyloid assembly inhibitors: The sugar scyllo-inositol, the polyphenol epigallocatechingallate, and the molecular Tweezer CLR01. ACS Chem. Neurosci. 2012, 3, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Grelle, G.; Otto, A.; Lorenz, M.; Frank, R.F.; Wanker, E.E.; Bieschke, J. Black tea theaflavins inhibit formation of toxic amyloid-β and α-synuclein fibrils. Biochemistry 2011, 50, 10624–10636. [Google Scholar] [CrossRef] [PubMed]
- Levites, Y.; Amit, T.; Mandel, S.; Youdim, M.B.H. Neuroprotection and neurorescue against A beta toxicity and PKC-dependent release of non-amyloidogenic soluble precursor protein by green tea polyphenol (−)-epigallocatechin-3-gallate. FASEB J. 2003, 17, 952–954. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Wu, M.M.; Lu, F.; Luo, N.; He, Z.P.; Yang, H. Involvement of α7 nAChR signaling cascade inepigallocatechingallate suppression of β-amyloid-induced apoptotic cortical neuronal insults. Mol. Neurobiol. 2014, 49, 66–67. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Y.; Chen, F.J.; Sha, L.; Wang, S.; Tao, L.; Yao, L.T.; He, M.; Yao, Z.M.; Liu, H.; Zhu, Z.; et al. (−)-Epigallocatechin-3-gallate ameliorates learning and memory deficits by adjusting the balance of TrkA/p75(NTR) signaling in APP/PS1 transgenic Mice. Mol. Neurobiol. 2014, 49, 1350–1363. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Shibata, M.; Ishikuro, R.; Tanida, M.; Taniguchi, Y.; Ikeda-Matsuo, Y.; Sobue, K. Epigallocatechingallate induces extracellular degradation of amyloid beta-protein by increasing neprilysin secretion from astrocytes through activation of ERK and PI3K pathways. Neuroscience 2017, 362, 70–78. [Google Scholar] [CrossRef] [PubMed]
- II Kim, T.; Lee, Y.K.; Park, S.G.; Choi, I.S.; Ban, J.O.; Park, H.K.; Nam, S.Y.; Yun, Y.W.; Han, S.B.; Oh, K.W.; et al. L-Theanine, an amino acid in green tea, attenuates beta-amyloid-induced cognitive dysfunction and neurotoxicity: Reduction in oxidative damage and inactivation of ERK/p38 kinase and NF-kappa B pathways. Free Radic. Biol. Med. 2009, 47, 1601–1610. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.F.; Zhu, Y.S.; Cao, X.S.; Sun, S.F.; Zhao, B.L. Mitochondrial toxic effects of a beta through mitofusins in the early pathogenesis of Alzheimer’s disease. Mol. Neurobiol. 2014, 50, 986–996. [Google Scholar] [CrossRef] [PubMed]
- Dragicevic, N.; Smith, A.; Lin, X.Y.; Yuan, F.; Copes, N.; Delic, V.; Tan, J.; Cao, C.H.; Shytle, R.D.; Bradshaw, P.C. Green tea epigallocatechin-3-gallate (EGCG) and other flavonoids reduce Alzheimer’s amyloid-induced mitochondrial dysfunction. J. Alzheimers Dis. 2011, 26, 507–521. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Jung, J.Y.; Kim, E.C.; Kim, H.J.; Kim, W.J.; Lee, E.J.; Kim, S.H. Inhibition of amyloid β peptide-induced neuronal cytotoxicity by EGCG. Korean J. Phys. Anthropol. 2005, 18, 139–147. [Google Scholar] [CrossRef]
- Wang, S.W.; Wang, Y.J.; Su, Y.J.; Zhou, W.W.; Yang, S.G.; Zhang, R.; Zhao, M.; Li, Y.N.; Zhang, Z.P.; Zhan, D.W.; et al. Rutin inhibits beta-amyloid aggregation and cytotoxicity, attenuates oxidative stress, and decreases the production of nitric oxide and proinflammatory cytokines. Neurotoxicology 2012, 33, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, A.; Zarb, C.; Caruana, M.; Ostermeier, U.; Ghio, S.; Högen, T.; Schmidt, F.; Giese, A.; Vassallo, N. Mitochondrial membrane permeabilisation by amyloid aggregates and protection by polyphenols. BBA-Biomembr. 2013, 1828, 2532–2543. [Google Scholar] [CrossRef] [PubMed]
- Chesser, A.S.; Ganeshan, V.; Yang, J.; Johnson, G.V.W. Epigallocatechin-3-gallateenhancesclearanceofphosphorylated tau in primary neurons. Nutr. Neurosci. 2016, 19, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.F.; Zhao, Y.; Nan, Y.; Wang, X.; Chen, Y.L.; Wang, S. (−)-Epigallocatechin-3- gallate ameliorates memory impairment and rescues the abnormal synaptic protein levels in the frontal cortex and hippocampus in a mouse model of Alzheimer’s disease. Neuroreport 2017, 28, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Wobst, H.J.; Sharma, A.; Diamond, M.I.; Wanker, E.E.; Bieschke, J. The green tea polyphenol (−)-epigallocatechingallate prevents the aggregation of tau protein into toxic oligomers at substoichiometric ratios. FEBS Lett. 2015, 589, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.P.; Wang, A.; Ye, J.H.; Zheng, X.Q.; Polito, C.A.; Lu, J.L.; Li, Q.S.; Liang, Y.R. Suppressive effects of tea catechins on breast cancer. Nutrients 2016, 8, 458. [Google Scholar] [CrossRef] [PubMed]
- Du, L.L.; Fu, Q.Y.; Xiang, L.P.; Zheng, X.Q.; Lu, J.L.; Ye, J.H.; Li, Q.S.; Polito, C.A.; Liang, Y.R. Tea polysaccharides and their bioactivities. Molecules 2016, 21, 1449. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.Y.; Li, Q.S.; Lin, X.M.; Qiao, R.Y.; Yang, R.; Li, X.M.; Dong, Z.B.; Xiang, L.P.; Zheng, X.Q.; Lu, J.L.; et al. Antidiabetic effects of tea. Molecules 2017, 22, 849. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Shi, Y.L.; Li, X.M.; Yang, R.; Cai, Z.Y.; Li, Q.S.; Ma, S.C.; Ye, J.H.; Lu, J.L.; Liang, Y.R.; et al. Food-grade encapsulation systems for (−)-epigallocatechingallate. Molecules 2018, 23, 445. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.H.; Augustin, M.A. Nano- and micro-particles for delivery of catechins: Physical and biological performance. Crit. Rev. Food Sci. 2018. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Type of Study | Country | Number of Subjects | Main Results | Reference |
|---|---|---|---|---|
| Six-year follow up longitudinal study | U.K. | Nine community-dwelling men and women. | The action-based memory of people with dementia of AD can be judged by looking at the process of preparing a cup of tea. | Rusted et al., 2002 [12] |
| Cross-sectional study | Japan | 1003 Japanese subjects aged 70 or older. | Consumption of ≥2 cups/day green tea was associated with a lower prevalence of CoI (OR: 0.46 (95% CI: 0.30, 0.72; p = 0.0006), compared to reference (≤3 cups/week) | Kuriyama et al., 2006 [13] |
| Prospective cohort study | Japan | 13,988 Japanese subjects aged 65 or older. | Green tea consumption was significantly associated with a lower risk of incident functional disability, even after adjustment for possible confounding factors. | Tomata et al., 2012 [14] |
| Population-based prospective study | Japan | 490 Japanese residents over 60 years old. | The multiple adjusted OR for the incidence of dementia was 0.26 (95% CI: 0.06, 1.06) among individuals who consumed green tea every day compared with those who did not consume green tea at all. No association was found between coffee or black tea consumption and the incidence of dementia or MCI. | Noguchi-Shinohara et al., 2014 [15] |
| Cross-sectional study | Japan | 1143 subjects. | Low green tea consumption (p = 0.032) were independently associated with a higher prevalence of CoI. The OR for drinking tea every day was 0.65 (95% CI: 0.47, 0.89) | Kitamura et al., 2016 [16] |
| A double-blind, randomized controlled study | Japan | 33 nursing home residents, consumed 2 g/day of green tea powder for 12 months. | Cognitive disfunction was not significantly different compared with that of the placebo group (OR: −0.61 (95% CI: −2.97, 1.74), p = 0.59). | Ide et al., 2016 [17] |
| Cross-sectional study | Singapore | 2501 adults aged 55 or older. | Cognitive decline ORs were 0.74 (95% CI: 0.54, 1.00) for low level, 0.78 (95% CI: 0.55, 1.11) for medium level, and 0.57 (95% CI: 0.32, 1.03) for high level tea intake. | Ng et al., 2008 [18] |
| Cross-sectional study | Singapore | 716 adults aged 55 or older. | Total tea consumption was independently associated with better performance on global cognition, memory, executive function, and information processing speed. | Feng et al., 2010 [19] |
| Longitudinal aging study | Singapore | 1615 adults aged 55 to 93. | The ORs were 0.79 (95% CI: 0.42, 1.48) for low tea consumption participants, 0.47 (95% CI: 0.25, 0.88) for medium tea consumption participants and 0.27 (95% CI: 0.11, 0.63) for high tea consumption participants (p = 0.01). | Feng et al., 2012 [20] |
| Cohort study | Singapore | 614 elderly aged 60 or older who were free of dementia and cognitive impairment. | Long-term tea consumption was associated with reduced depressive and anxiety symptoms among community-living elderly. | Chan et al., 2017 [21] |
| Cohort study | China | 681 unrelated Chinese nonagenarians/centenarians (67.25% women). | Habits of tea drinking had a significantly positive impact on CoI in men, but no association of CoI with tea drinking in women. | Huang et al., 2009 [22] |
| Population-based, nest case-control study | China | 5691 elderly residents aged 65 or older (1489 cognitive decline and 4822 normal cognitive function). | An inverse association between tea drinking and cognitive decline was found (OR: 0.82; 95% CI: 0.69, 1.00, p = 0.0468). | Chen et al., 2012 [23] |
| Population-based survey | China | 4579 elders aged 60 or older from the town of Weitang in Suzhou, China. | An inverse association was found between tea consumption (of any type) and prevalence of CoI (OR: 0.74, 95%CI: 0.57–0.98, p = 0.032). | Gu et al., 2017 [24] |
| Population-based study | China | 1368 rural community-dwelling individuals aged 60 or older (59.3% female). | Daily tea consumption was associated with a lower likelihood of depressive symptoms in older Chinese people living in a rural community. The association appears to be independent of cerebrovascular disease and atherosclerosis. | Feng et al., 2013 [25] |
| Cross-sectional Study | China | 9375 adults aged 60 or older. | An inverse correlation was found between tea consumption and prevalence of CoI. | Shen et al., 2015 [26] |
| Cross-sectional study | China | 2015 adults aged 65 or older (42.2% men). | Tea consumption was associated with low prevalence of AD. | Yang et al., 2016 [27] |
| Longitudinal Healthy Longevity Survey | China | 32,606 subjects aged 65 or older (13,429 men and 19,177 women). | High frequency of tea consumption was significantly associated with reduced OR of CoI. | Qiu et al., 2012 [28] |
| Longitudinal Healthy Longevity Survey | China | 7139 participants aged 80 to 115 years. | Regular tea drinking was associated with better cognitive function in oldest-old Chinese, with regression coefficient 0.72 (p < 0.0001) for daily drinking and 0.41(p = 0.01) for occasional drinking. | Feng et al., 2012 [29] |
| Prevalence survey | China | 1000 residents aged ≥60 years old. | Drinking tea reduced the incidence of MCI (p < 0.05) | Yang et al., 2017 [30] |
| Cross-sectional study | China | 870 elders aged ≥90 years old. | Among the Chinese nonagenarians and centenarians, no significant correlation between tea consumption and the prevalence of MCI. | Wang et al., 2010 [31] |
| Cross-sectional study | Norway | 2031 adults aged 70–74 years (55% women). | The associations between intake of tea and cognition were approximately linearly dose-dependent. | Nurk et al., 2009 [32] |
| Population-based study | 23 developed countries | Adults from 23 developed countries and given different genetic backgrounds. | A significant inverse correlation was found between dietary consumption of flavonoids and rate of AD or related dementias. | Beking et al., 2010 [33] |
| Meta-analyses | Asia, Europe, Australia, and North America. | 52,503 participants distributed in Asia, Europe, Australia, and America. | Daily tea drinking was associated with decreased risk of CoI, MCI andcognitive decline in the elderly. However, the association between tea intake and AD remained elusive. | Ma et al., 2016 [34] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polito, C.A.; Cai, Z.-Y.; Shi, Y.-L.; Li, X.-M.; Yang, R.; Shi, M.; Li, Q.-S.; Ma, S.-C.; Xiang, L.-P.; Wang, K.-R.; et al. Association of Tea Consumption with Risk of Alzheimer’s Disease and Anti-Beta-Amyloid Effects of Tea. Nutrients 2018, 10, 655. https://doi.org/10.3390/nu10050655
Polito CA, Cai Z-Y, Shi Y-L, Li X-M, Yang R, Shi M, Li Q-S, Ma S-C, Xiang L-P, Wang K-R, et al. Association of Tea Consumption with Risk of Alzheimer’s Disease and Anti-Beta-Amyloid Effects of Tea. Nutrients. 2018; 10(5):655. https://doi.org/10.3390/nu10050655
Chicago/Turabian StylePolito, Curt Anthony, Zhuo-Yu Cai, Yun-Long Shi, Xu-Min Li, Rui Yang, Meng Shi, Qing-Sheng Li, Shi-Cheng Ma, Li-Ping Xiang, Kai-Rong Wang, and et al. 2018. "Association of Tea Consumption with Risk of Alzheimer’s Disease and Anti-Beta-Amyloid Effects of Tea" Nutrients 10, no. 5: 655. https://doi.org/10.3390/nu10050655
APA StylePolito, C. A., Cai, Z. -Y., Shi, Y. -L., Li, X. -M., Yang, R., Shi, M., Li, Q. -S., Ma, S. -C., Xiang, L. -P., Wang, K. -R., Ye, J. -H., Lu, J. -L., Zheng, X. -Q., & Liang, Y. -R. (2018). Association of Tea Consumption with Risk of Alzheimer’s Disease and Anti-Beta-Amyloid Effects of Tea. Nutrients, 10(5), 655. https://doi.org/10.3390/nu10050655