Taurine Supplementation Alleviates Puromycin Aminonucleoside Damage by Modulating Endoplasmic Reticulum Stress and Mitochondrial-Related Apoptosis in Rat Kidney

 ,
,  ,
,  , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animal Treatment
2.3. Histopathology and Immunohistochemistry
2.4. Transmission Electron Microscopy
2.5. Statistical Analysis
3. Results
3.1. Metabolical and Histopathological Effects of TAU in Nephrotic Rats
3.2. TAU Changes Podocytes Markers Distributions in Nephrotic Rats
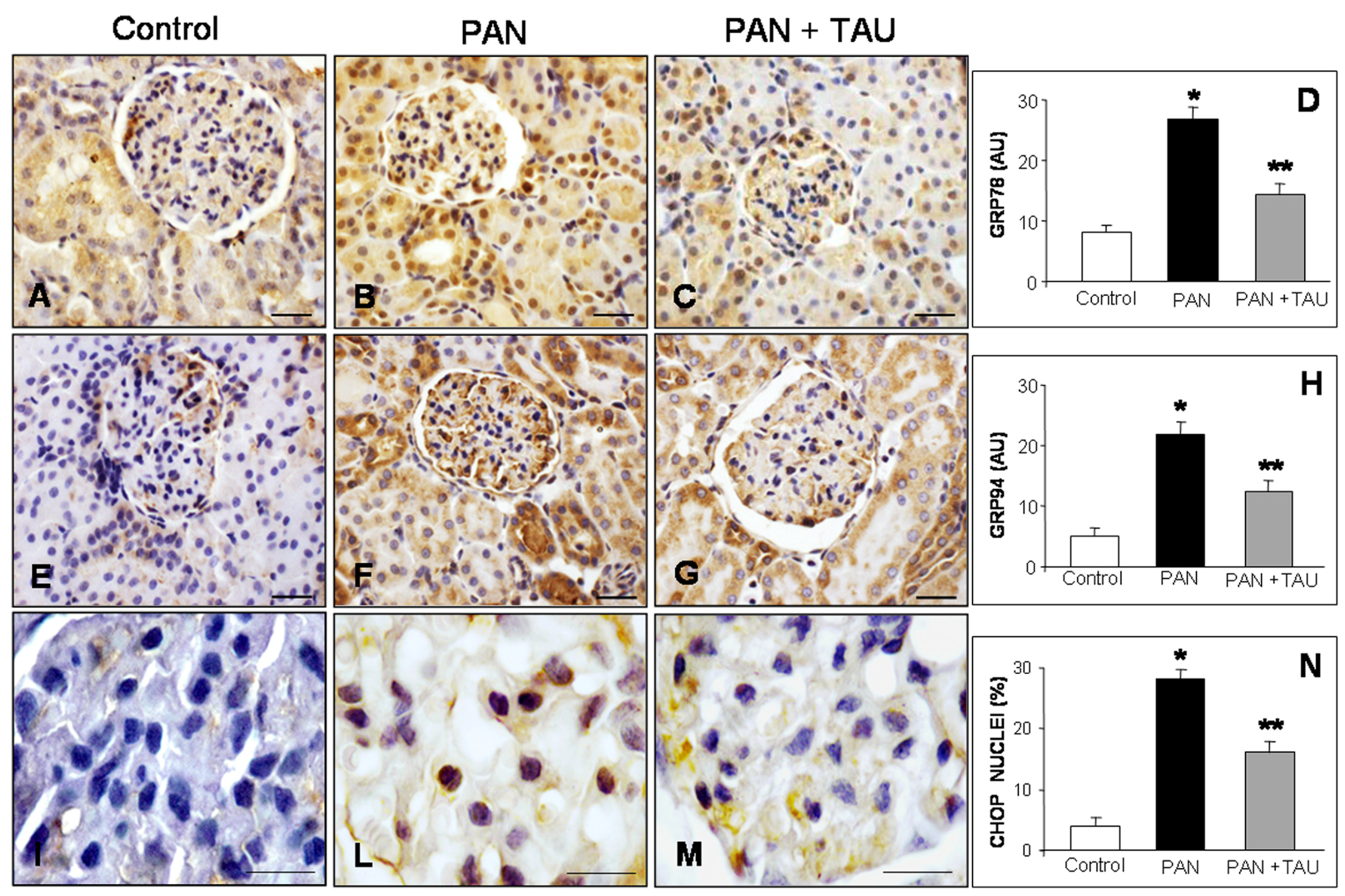
3.3. TAU Reduces Glomerular ER Chaperones and Apoptosis
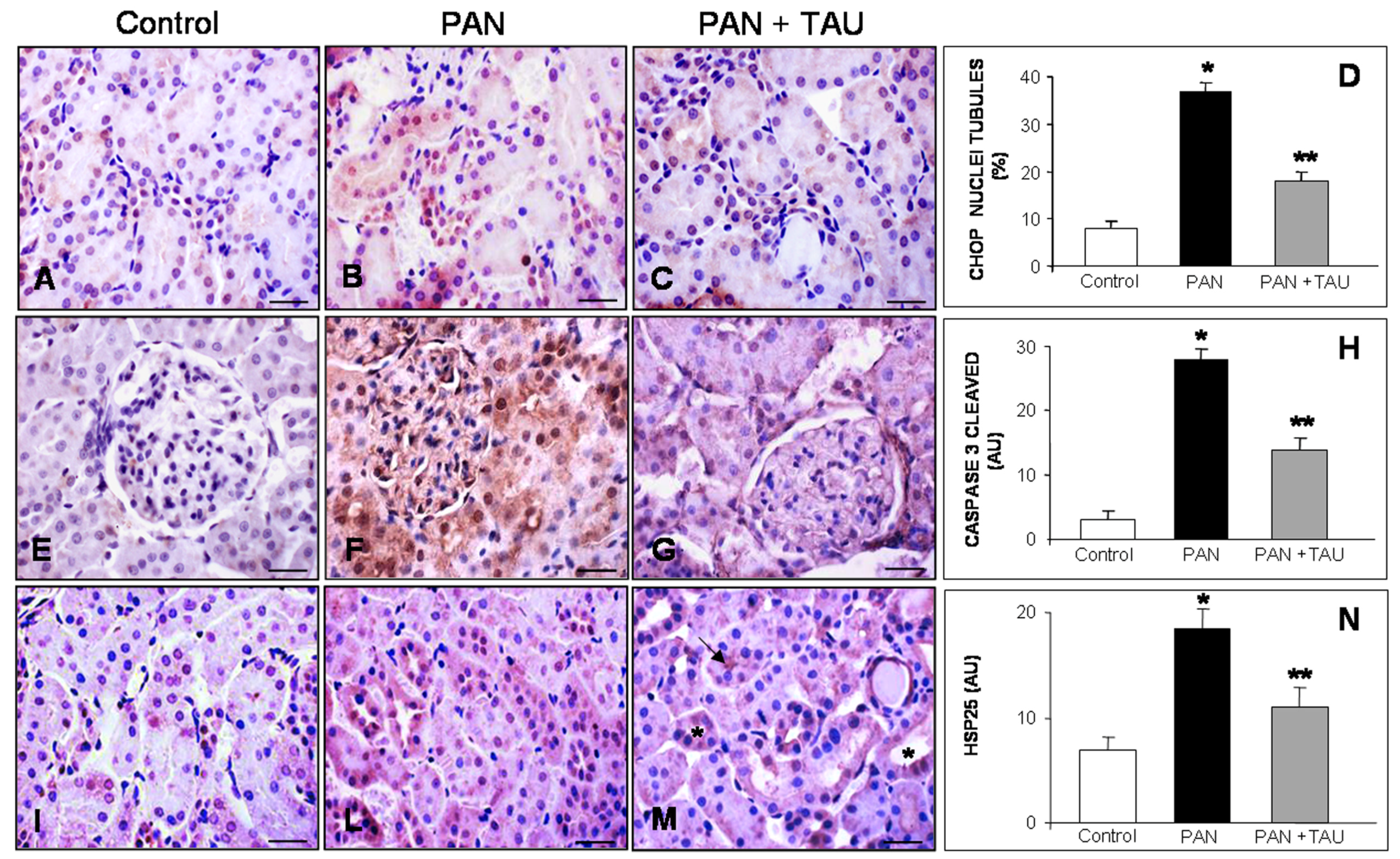
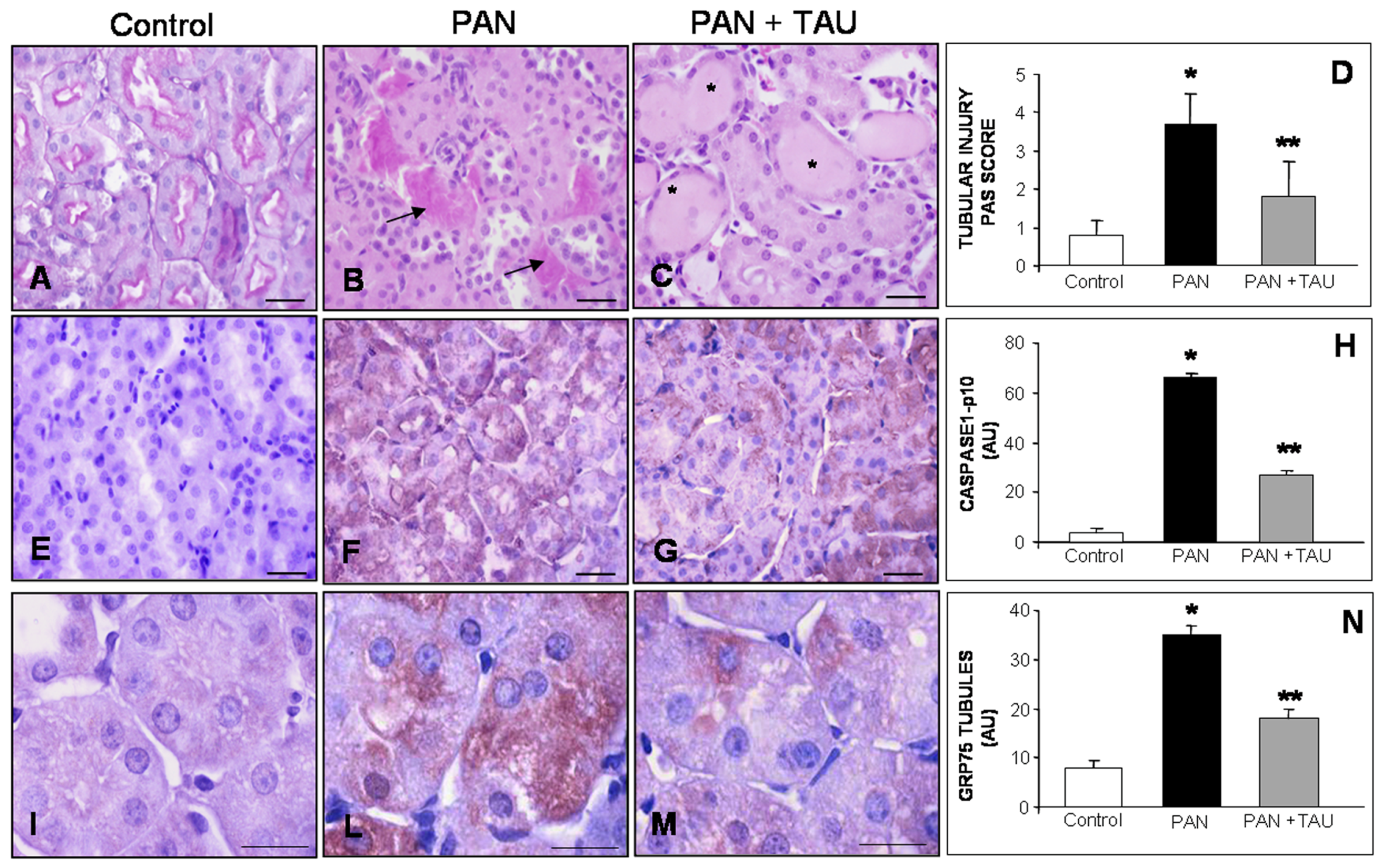
3.4. TAU Decreases Tubular Apoptosis and Oxidative Damage
3.5. TAU Improves Tubular Inflammation and Mitochondrial Stress in PAN Nephrosis
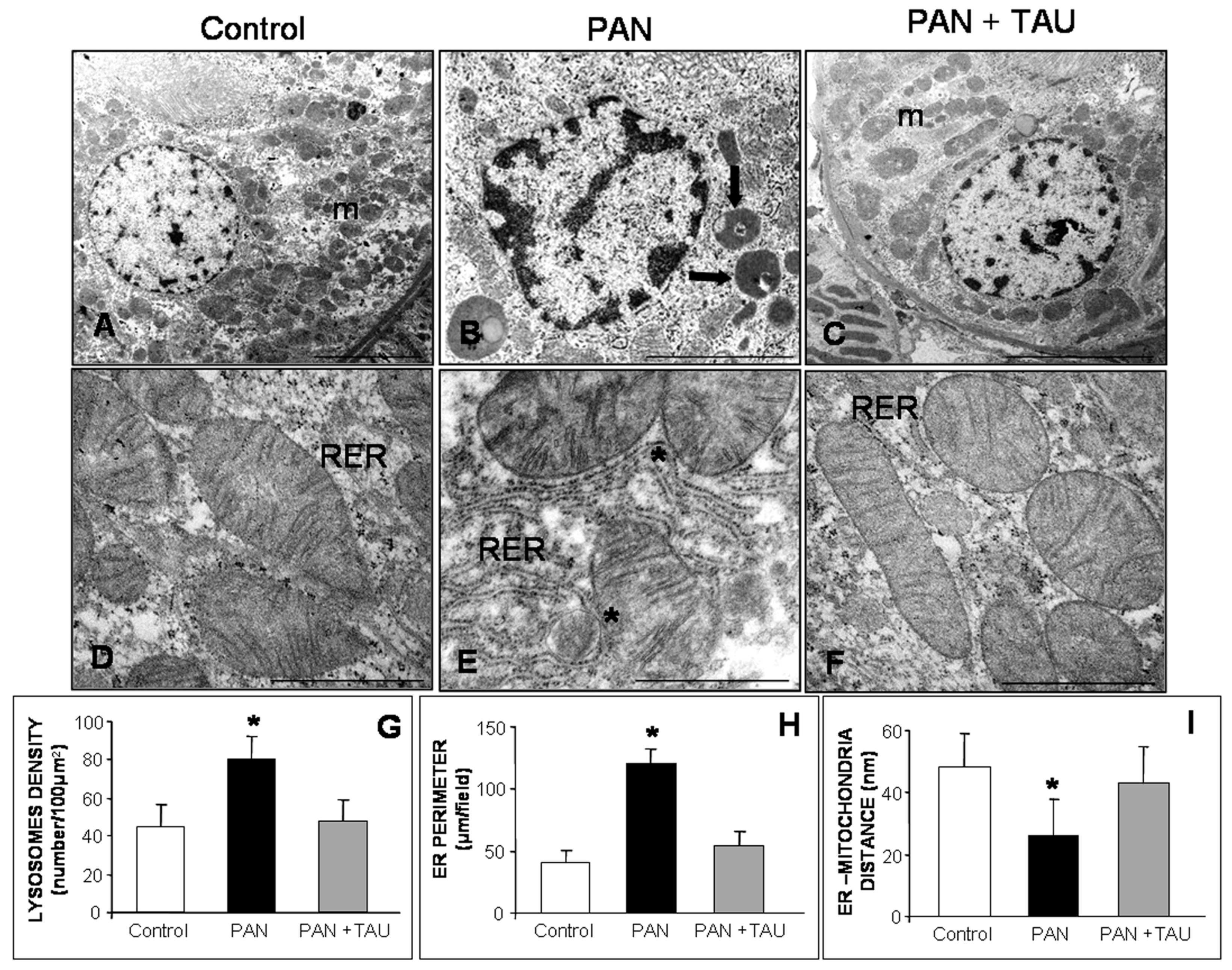
3.6. Effect of TAU on Mitochondria and Rough ER in Nephrotic Proximal Tubules
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pal, A.; Kastel, F. History of nephrotic syndrome and evolution of its treatment. Front. Pediatr. 2016, 4, 56. [Google Scholar] [CrossRef] [PubMed]
- Barisoni, L.; Schnaper, H.; Kopp, J. A proposed taxonomy for the podocytopathies: A reassessment of the primary nephritic diseases. Clin. J. Am. Soc. Nephrol. 2007, 2, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Vivarelli, M.; Massella, L.; Ruggiero, B.; Emma, F. Minimal Change Disease. Clin. J. Am. Soc. Nephrol. 2017, 12, 332–345. [Google Scholar] [CrossRef] [PubMed]
- Teeninga, N.; Kist van Holthe, K.; Nauta, J. Extending prednisolone treatment does not reduce relapses in childhood nephrotic syndrome. J. Am. Soc. Nephrol. 2012, 24, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Hogan, J.; Radhakrishnan, J. The treatment of minimal change disease in adults. J. Am. Soc. Nephrol. 2013, 24, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Mathieson, P. Minimal change nephropathy and focal segmental glomerulosclerosis. Semin. Immunopathol. 2007, 29, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Greka, A. Human genetics of nephrotic syndrome and the quest for precision medicine. Curr. Opin. Nephrol. Hypertens. 2016, 25, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Jiang, H.; Ying, M.; Xie, Z.; Li, X.; Wang, H.; Zhao, J.; Lin, C.; Wang, Y.; Feng, S.; et al. Calcineurin inhibitors cyclosporine A and tacrolimus protect against podocyte injury induced by puromycin aminonucleoside in rodent models. Sci. Rep. 2016, 6, 32087. [Google Scholar] [CrossRef] [PubMed]
- Pippin, J.; Brinkkoetter, P.; Cormack-Aboud, F.; Durvasula, R.; Hauser, P.; Kowalewska, J.; Krofft, R.; Logar, C.; Marshall, C.; Ohse, T.; et al. Inducible rodent models of acquired podocyte diseases. Am. J. Physiol. Ren. Physiol. 2009, 296, F213–F229. [Google Scholar] [CrossRef] [PubMed]
- Haraldsson, B.; Nystrom, J.; Deen, W. Properties of the glomerular barrier and mechanisms of proteinuria. Physiol. Rev. 2008, 88, 451–487. [Google Scholar] [CrossRef] [PubMed]
- Kriz, K.; Shirato, I.; Nagata, M.; LeHir, M.; Lemiev, K. The podocyte’s response to stress: The enigma of foot process effacement. Am. J. Physiol. Ren. Physiol. 2013, 304, F333–F347. [Google Scholar] [CrossRef] [PubMed]
- Burford, J.; Gyarmati, G.; Shirato, I.; Kriz, W.; Lemiev, K.; Peti-Peterdi, J. Combined use of electron microscopy and intravital imaging captures morphological and functional features of podocyte detachment. Pflugers Arch. Eur. J. Physiol. 2017, 469, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Welsh, G.; Saleem, S. The podocyte cytoskeleton-key to a functioning glomerulus in health and disease. Nat. Rev. Nephrol. 2011, 8, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Yan, K.; Khoshnoodi, J.; Ruotsalainen, V.; Tryggvason, K. N-linked glycosylation is critical for the plasma membrane localization of nephrin. J. Am. Soc. Nephrol. 2002, 13, 1385–1389. [Google Scholar] [CrossRef] [PubMed]
- Swiatecka-Urban, A. Endocytic trafficking at the mature podocyte slit diaphragm. Front. Pediatr. 2017, 5, 32. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Sunq, A.; Roth, R.; Hou, J. Inducible expression of claudin-1 in glomerular podocytes generates aberrant tight junctions and proteinuria through slit diaphragm destabilization. J. Am. Soc. Nephrol. 2017, 28, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Greka, A.; Mundel, P. Regulation of podocyte actin dynamics by calcium. Semin. Nephrol. 2012, 32, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Li, X.; Li, B.; Guo, J.; Zhang, Y.; Ding, J. Calpain-mediated cleavage of calcineurin in puromycin aminonucleoside-induced podocyte injury. PLoS ONE 2016, 11, e0155504. [Google Scholar] [CrossRef] [PubMed]
- Inagi, R.; Ishimoto, Y.; Nangaku, M. Proteostasis in endoplasmic reticulum-new mechanisms in kidney disease. Nat. Rev. Nephrol. 2014, 10, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, C.; Brodsky, J. The delicate balance between secreted protein folding and endoplasmic reticulum-associated degradation in human physiology. Physiol. Rev. 2012, 92, 537–576. [Google Scholar] [CrossRef] [PubMed]
- Krebs, J.; Agellon, L.; Michalak, M. Ca2+ homeostasis and endoplasmic reticulum (ER) stress: An integrated view of calcium signaling. Biochem. Biophys. Res. Commun. 2015, 460, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, M.; Yoshida, H. Endoplasmic reticulum stress in kidney function and disease. Curr. Opin. Nephrol. Hypertens. 2015, 24, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Inagi, R. Endoplasmic reticulum stress as a progression factor for kidney injury. Curr. Opin. Pharmacol. 2010, 10, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Cybulsky, A. Endoplasmic reticulum stress, the unfolded protein response and autophagy in kidney diseases. Nat. Rev. Nephrol. 2017, 13, 681–696. [Google Scholar] [CrossRef] [PubMed]
- El Karoui, K.; Vian, A.; Dellis, O.; Bagattin, A.; Nguyen, C.; Baron, W.; Burtin, M.; Broueilh, M.; Heidet, L.; Mollet, G.; et al. Endoplasmic reticulum stress drives proteinuria-induced kidney lesions via lipocalin 2. Nat. Commun. 2016, 7, 10330. [Google Scholar] [CrossRef] [PubMed]
- Lindenmeyer, M.; Rastaldi, M.; Ikehata, M.; Neusser, M.; Kretzler, M.; Cohen, C.; Schlondorff, D. Proteinuria and hyperglycemia induce endoplasmic reticulum stress. J. Am. Soc. Nephrol. 2008, 19, 2225–2236. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Lee, A. Role of the unfolded protein response, GRP78 and GRP94 in organ homeostasis. J. Cell Physiol. 2015, 230, 1413–1420. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Jin, J.; Ogawa, M.; Aoe, T. Dysfunction of the ER chaperone BiP accelerates the renal tubular injury. Biochem. Biophys. Res. Commun. 2008, 366, 1048–1053. [Google Scholar] [CrossRef] [PubMed]
- Shore, G.; Papa, F.; Oakes, S. Signalling cell death from the endoplasmic reticulum stress response. Curr. Opin. Cell Biol. 2011, 23, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Bijian, K.; Cybulsky, A. Stress proteins in glomerular epithelial cell injury. Contrib. Nephrol. 2005, 148, 8–20. [Google Scholar] [PubMed]
- Sreedharan, R.; Van Why, S. Heat shock proteins in the kidney. Pediatr. Nephrol. 2016, 31, 1561–1570. [Google Scholar] [CrossRef] [PubMed]
- Lanneu, D.; Brunet, M.; Frisan, E.; Solary, E.; Fontenay, M.; Garrido, C. Heat shock proteins: Essential proteins for apoptosis regulation. J. Cell. Mol. Med. 2008, 12, 743–761. [Google Scholar] [CrossRef] [PubMed]
- Smoyer, W.; Gupta, A.; Mundel, P.; Ballew, J.; Welsh, M. Altered expression of glomerular heat shock protein 27 in experimental nephrotic syndrome. J. Clin. Investig. 1996, 97, 2697–2704. [Google Scholar] [CrossRef] [PubMed]
- Smoyer, W.; Ransom, R. Hsp27 regulates podocyte cytoskeletal changes in an in vitro model of podocyte process retraction. FASEB J. 2002, 16, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Wadhwa, P.; Taira, K.; Kaul, S. An Hsp70 family chaperone, mortalin/mthsp70/PBP74/Grp75: What, when, where? Cell Stress Chaperones 2002, 7, 309–316. [Google Scholar] [CrossRef]
- Imasawa, T.; Rossignol, R. Podocyte energy metabolism and glomerular diseases. Int. J. Biochem. Cell Biol. 2013, 45, 2109–2118. [Google Scholar] [CrossRef] [PubMed]
- Che, R.; Yuan, Y.; Huang, Y.; Zhang, A. Mitochondrial dysfunction in the pathophysiology of renal diseases. Am. J. Physiol. Ren. Physiol. 2014, 306, F367–F378. [Google Scholar] [CrossRef] [PubMed]
- Granquist, A.; Nilsson, U.; Ebefors, K.; Haraldsson, B.; Nystrom, J. Impaired glomerular and tubular antioxidative defense mechanisms in nephrotic syndrome. Am. J. Physiol. Ren. Physiol. 2010, 299, F898–F904. [Google Scholar] [CrossRef] [PubMed]
- Tian, N.; Arany, I.; Waxman, D.; Baliga, R. Cytochrome P450 2B1 gene silencing attenuates puromycin aminonucleoside-induced cytotoxicity in glomerular epithelial cells. Kidney Int. 2010, 78, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Gaull, G. Taurine as a conditionally essential nutrient in man. J. Am. Coll. Nutr. 1986, 5, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Yamori, Y.; Taguchi, T.; Hamada, A.; Kunimasia, K.; Mori, H.; Mori, M. Taurine in health and diseases: Consistent evidence from experimental and epidemiological studies. J. Biomed. Sci. 2010, 17, S6. [Google Scholar] [CrossRef] [PubMed]
- Jong, C.; Ito, T.; Prentice, H.; Wu, J.; Schaffer, S. Role of mitochondria and endoplasmic reticulum in taurine-deficiency-mediated apoptosis. Nutrients 2017, 9, 795. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, S.; Kim, H.W. Effects and mechanisms of taurine as a therapeutic agent. Biomol. Ther. 2018, 26, 225–241. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Chesney, R. The role of taurine in renal disorders. Amino Acids 2012, 43, 2249–2263. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Schaffer, S.; Azuma, J. The potential usefulness of taurine on diabetes mellitus and its complications. Amino Acids 2012, 42, 1529–1539. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, P.; Basak, P.; Gosh, S.; Kundu, M.; Sil, P. Prophylactic role of taurine and its derivatives against diabetes mellitus and its related complications. Food Chem. Toxicol. 2017, 110, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Guo, J.; Zhang, Y.; Zhang, J. The beneficial effects of taurine in preventing metabolic syndrome. Food Funct. 2017, 7, 1849–1863. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, N.; Rao, P.; Arumugam, V. Inhibitory effect of taurine on puromycin aminonucleoside-induced hyperlipidemia in rats. J. Clin. Biochem. Nutr. 1993, 15, 203–210. [Google Scholar] [CrossRef]
- Feng, Y.; Li, J.; Yang, Y.; Yang, Q.; Lv, Q.; Gao, Y.; Hu, J. Synergistic effects of taurine and l-arginine on attenuating insulin resistance hypertension. Adv. Exp. Med. Biol. 2013, 775, 427–435. [Google Scholar] [PubMed]
- Kerlin, B.; Waller, A.; Sharma, R.; Chanley, M.; Nieman, M.; Smoyer, W. Disease severity correlates with thrombotic capacity in experimental nephrotic syndrome. J. Am. Soc. Nephrol. 2015, 26, 3009–3019. [Google Scholar] [CrossRef] [PubMed]
- Moloney, M.A.; Casey, R.G.; O’Donnell, D.H.; Fitzgerald, P.; Thompson, C.; Bouchier-Hayes, D.J. Two weeks taurine supplementation reverses endothelial dysfunction in young male type 1 diabetics. Diabetes Vasc. Dis. Res. 2010, 7, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Stacchiotti, A.; Lavazza, A.; Rezzani, R.; Borsani, E.; Rodella, L.; Bianchi, R. Mercuric-chloride induced alterations in stress proteins distribution in rat kidney. Histol. Histopathol. 2004, 19, 1209–1218. [Google Scholar] [PubMed]
- Sitrin, J.; Suto, E.; Wuster, A.; Eastham-Anderson, J.; Kim, J.; Austin, C.; Lee, W.; Beherens, T. The Ox40/Ox40 ligand pathway promotes pathogenic Th cell responses, plasmablast accumulation, and lupus nephritis in NZB/W F1 mice. J. Immunol. 2017, 109, 1238–1249. [Google Scholar] [CrossRef] [PubMed]
- Stacchiotti, A.; Ricci, F.; Rezzani, R.; Li Volti, G.; Borsani, E.; Lavazza, A.; Bianchi, R.; Rodella, L.F. Tubular stress proteins and nitric oxide synthase expression in rat kidney exposed to mercuric chloride and melatonin. J. Histochem. Cytochem. 2006, 54, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Imai, M.; Song, H.; Quigg, R.; Tomlinson, S. Complement inhibitors targeted to the proximal tubule prevent injury in experimental nephrotic syndrome and demonstrate a key role for C5b-9. J. Immunol. 2005, 174, 5750–5757. [Google Scholar] [CrossRef] [PubMed]
- Stacchiotti, A.; Favero, G.; Giugno, L.; Lavazza, A.; Reiter, R.; Rodella, L.F.; Rezzani, R. Mitochondrial and metabolic dysfunction in renal convoluted tubules of obese mice: Protective role of melatonin. PLoS ONE 2014, 9, e111141. [Google Scholar] [CrossRef] [PubMed]
- Stacchiotti, A.; Favero, G.; Lavazza, A.; Golic, I.; Aleksic, M.; Korac, A.; Rodella, L.F.; Rezzani, R. Hepatic macrosteatosis is partially converted to microsteatosis by melatonin supplementation in ob/ob mice nonalcoholic fatty liver disease. PLoS ONE 2016, 11, e0148115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patrakka, J.; Lahdenkari, A.; Koskimies, O.; Holmberg, C.; Wartiovaara, J.; Jalanko, H. The number of podocyte-slit diaphragms is decreased in minimal change nephrotic syndrome. Pediatr. Res. 2002, 52, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Sanz, A.; Santamaria, B.; Ruiz-Ortega, J.; Egido, J.; Ortiz, A. Mechanisms of renal apoptosis in health and disease 2. J. Am. Soc. Nephrol. 2008, 19, 1634–1642. [Google Scholar] [CrossRef] [PubMed]
- Patergnani, S.; Suski, J.; Agnoletto, C.; Bononi, A.; Bonora, M.; De Marchi, E.; Giorgi, C.; Marchi, S.; Missiroli, S.; Poletti, F.; et al. Calcium signaling around mitochondria associated membranes (MAMs). Cell Commun. Signal. 2011, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- De Zeeuw, D.; Remuzzi, G.; Parving, H.; Keane, W.; Zhang, Z.; Shahinfar, S.; Snapinn, S.; Cooper, M.; Mitch, W.; Brenner, B. Proteinuria, a target for renoprotection in patients with type 2 diabetic nephropathy: Lessons from RENAAL. Kidney Int. 2004, 65, 2309–2320. [Google Scholar] [CrossRef] [PubMed]
- Abbate, M.; Zoja, C.; Remuzzi, G. How does proteinuria cause progressive renal damage? J. Am. Soc. Nephrol. 2006, 17, 2974–2984. [Google Scholar] [CrossRef] [PubMed]
- Cybulsky, A. Endoplasmic reticulum stress in proteinuric kidney disease. Kidney Int. 2010, 77, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, S.; Hiromura, K.; Tomioka, M.; Takahashi, S.; Sakairi, T.; Maeshima, A.; Kaneko, Y.; Kuroiwa, T.; Nojima, Y. The immunosuppressive drug mizoribine directly prevents podocyte injury in puromycinaminonucleoside nephrosis. Nephron Exp. Nephrol. 2010, 116, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Hammadi, M.; Oulidi, A.; Gackiere, F.; Katsogiannou, M.; Slomianny, C.; Roudbaraki, M.; Dewailly, E.; Delcourt, P.; Lepage, G.; Lotteau, S.; et al. Modulation of ER stress and apoptosis by endoplasmic reticulum calcium leak via translocon during unfolded protein response: Involvement of GRP78. FASEB J. 2013, 27, 1600–1609. [Google Scholar] [CrossRef] [PubMed]
- Rovetta, F.; Stacchiotti, A.; Consiglio, A.; Cadei, M.; Grigolato, P.; Lavazza, A.; Rezzani, R.; Aleo, M.F. ER signaling regulation drives the switch between autophagy and apoptosis in NRK-52E cells exposed to cisplatin. Exp. Cell Res. 2012, 318, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ren, Y.; Liu, Y.; Gao, K.; Liu, Z.; Zhang, Z. Inhibition of starvation-triggered endoplasmic reticulum stress, autophagy, and apoptosis in ARPE-19 cells by taurine through modulating the expression of calpain-1 and calpain-2. Int. J. Mol. Sci. 2017, 18, 2146. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Zhan, Y.; Liu, H.; Zhao, T.; Yang, L.; Zhang, H.; Dong, X.; Li, P. Yi Qi Qing Re Gao formula ameliorates puromycin aminonucleoside-induced nephrosis by suppressing inflammation and apoptosis. BMC Complement. Altern. Med. 2015, 15, 155. [Google Scholar] [CrossRef] [PubMed]
- Mallipattu, S.; He, H. The beneficial role of retinoids in glomerular disease. Front. Med. 2015, 2, 16. [Google Scholar] [CrossRef] [PubMed]
- Prentice, H.; Modi, J.; Wu, J. Mechanisms of neuronal protection against excitotoxicity, endoplasmic reticulum stress, and mitochondrial dysfunction in stroke and neurodegenerative diseases. Oxid. Med. Cell. Longev. 2015, 2015, 964518. [Google Scholar] [CrossRef] [PubMed]
- Cocchiaro, P.; de Pasquale, V.; Della Morte, D.; Taffuri, S.; Avallone, L.; Pizard, A.; Moles, A.; Pavone, L. The multifaceted role of the lysosomal protease cathepsins in kidney disease. Front. Cell Dev. Biol. 2017, 5, 114. [Google Scholar] [CrossRef] [PubMed]
- Mozaffari, M.S.; Abdelsayed, R.; Patel, C.; Wimborne, H.; Liu, J.Y.; Schaffer, S.W. Differential effects of taurine treatment and taurine deficiency on the outcome of renal ischemia reperfusion injury. J. Biomed. Sci. 2010, 17 (Suppl. 1), S32. [Google Scholar] [CrossRef] [PubMed]
- Shimada, K.; Jong, C.; Takahashi, K.; Schaffer, S. Role of ROS production and turnover in the antioxidant activity of taurine. Adv. Exp. Med. Biol. 2015, 803, 581–596. [Google Scholar] [PubMed]
- Chebotareva, N.; Bobkova, I.; Shilou, E. Heat shock proteins and kidney disease: Perspectives of HSP therapy. Cell Stress Chaperones 2017, 22, 319–343. [Google Scholar] [CrossRef] [PubMed]
- Bravo, R.; Vicencio, J.; Parra, V.; Troncoso, R.; Munoz, J.; Bui, M.; Quiroga, C.; Rodriguez, A.; Verdejo, H.; Ferreira, J.; et al. Increased ER-mitochondrial coupling promotes mitochondrial respiration and bioenergetics during early phases of ER stress. J. Cell Sci. 2011, 124, 2143–2152. [Google Scholar] [CrossRef] [PubMed]
- El Idrissi, A. Taurine increases mitochondrial buffering of calcium: Role of neuroprotection. Amino Acids 2008, 34, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Xie, D.; Xian, W.; Cao, H.; Su, W.; Yang, J. Involvement of endoplasmic reticulum stress in albuminuria induced inflammasome activation in renal proximal tubular cells. PLoS ONE 2013, 8, e72344. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Ding, G.; Zhao, M.; Bai, M.; Yang, L.; Ni, J.; Wang, R.; Jia, Z.; Huang, S.; Zhang, A. NLRP3 inflammasome mediates albumin-induced renal tubular injury through impaired mitochondrial function. J. Biol. Chem. 2014, 289, 25101–25111. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, M.; Haynes, C.; Pellegrino, M. The mitochondrial unfolded protein response: Signaling from the powerhouse. J. Biol. Chem. 2017, 292, 13500–13506. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (N = 4) | Taurine (N = 4) | PAN (N = 6) | PAN + Taurine (N = 6) | |
|---|---|---|---|---|
| Body weight (g) | 248.5 ± 10.0 | 240.8 ± 10.0 | 200.6 ± 12.00 * | 230.5 ± 10.0 ** |
| Urinary Volume (mL/day) | 5.5 ± 0.9 | 5.3 ± 0.9 | 18.8 ± 10.0 * | 9.5 ± 8.5 ** |
| Proteinuria | ND | ND | 3+ | 1+ |
| Urobilinogen (mg/dL) | 0.2 | 0.2 | 0.2 | 0.2 |
| Hemoglobin (mg/dL) | 0.025 | 0.025 | 0.025 | 0.025 |
| Leukocytes (N/dL) | absent | absent | 125 (2+) | 70 (2+) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stacchiotti, A.; Favero, G.; Lavazza, A.; Monsalve, M.; Rodella, L.F.; Rezzani, R. Taurine Supplementation Alleviates Puromycin Aminonucleoside Damage by Modulating Endoplasmic Reticulum Stress and Mitochondrial-Related Apoptosis in Rat Kidney. Nutrients 2018, 10, 689. https://doi.org/10.3390/nu10060689
Stacchiotti A, Favero G, Lavazza A, Monsalve M, Rodella LF, Rezzani R. Taurine Supplementation Alleviates Puromycin Aminonucleoside Damage by Modulating Endoplasmic Reticulum Stress and Mitochondrial-Related Apoptosis in Rat Kidney. Nutrients. 2018; 10(6):689. https://doi.org/10.3390/nu10060689
Chicago/Turabian StyleStacchiotti, Alessandra, Gaia Favero, Antonio Lavazza, Maria Monsalve, Luigi Fabrizio Rodella, and Rita Rezzani. 2018. "Taurine Supplementation Alleviates Puromycin Aminonucleoside Damage by Modulating Endoplasmic Reticulum Stress and Mitochondrial-Related Apoptosis in Rat Kidney" Nutrients 10, no. 6: 689. https://doi.org/10.3390/nu10060689
APA StyleStacchiotti, A., Favero, G., Lavazza, A., Monsalve, M., Rodella, L. F., & Rezzani, R. (2018). Taurine Supplementation Alleviates Puromycin Aminonucleoside Damage by Modulating Endoplasmic Reticulum Stress and Mitochondrial-Related Apoptosis in Rat Kidney. Nutrients, 10(6), 689. https://doi.org/10.3390/nu10060689