Effects of Vitamin K2 on the Expression of Genes Involved in Bile Acid Synthesis and Glucose Homeostasis in Mice with Humanized PXR


 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Cell Culture
2.4. High-Performance Liquid Chromatography (HPLC) for Determination of MK-4 Levels in the Liver
2.5. RNA Extraction and mRNA Quantification
2.6. DNA Microarray
2.7. Statistical Analysis
3. Results
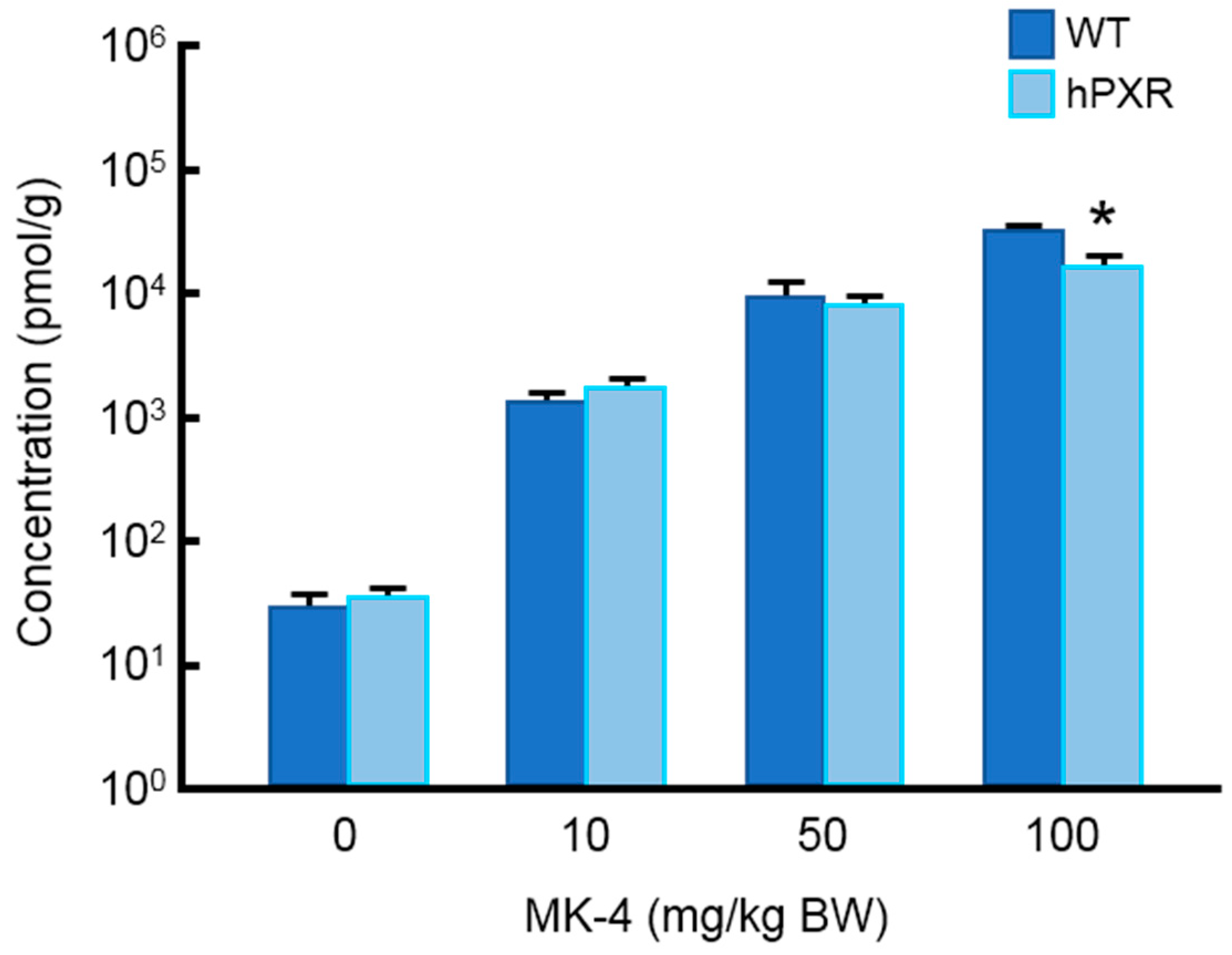
3.1. MK-4 Concentration in the Liver of hPXR and WT Mice
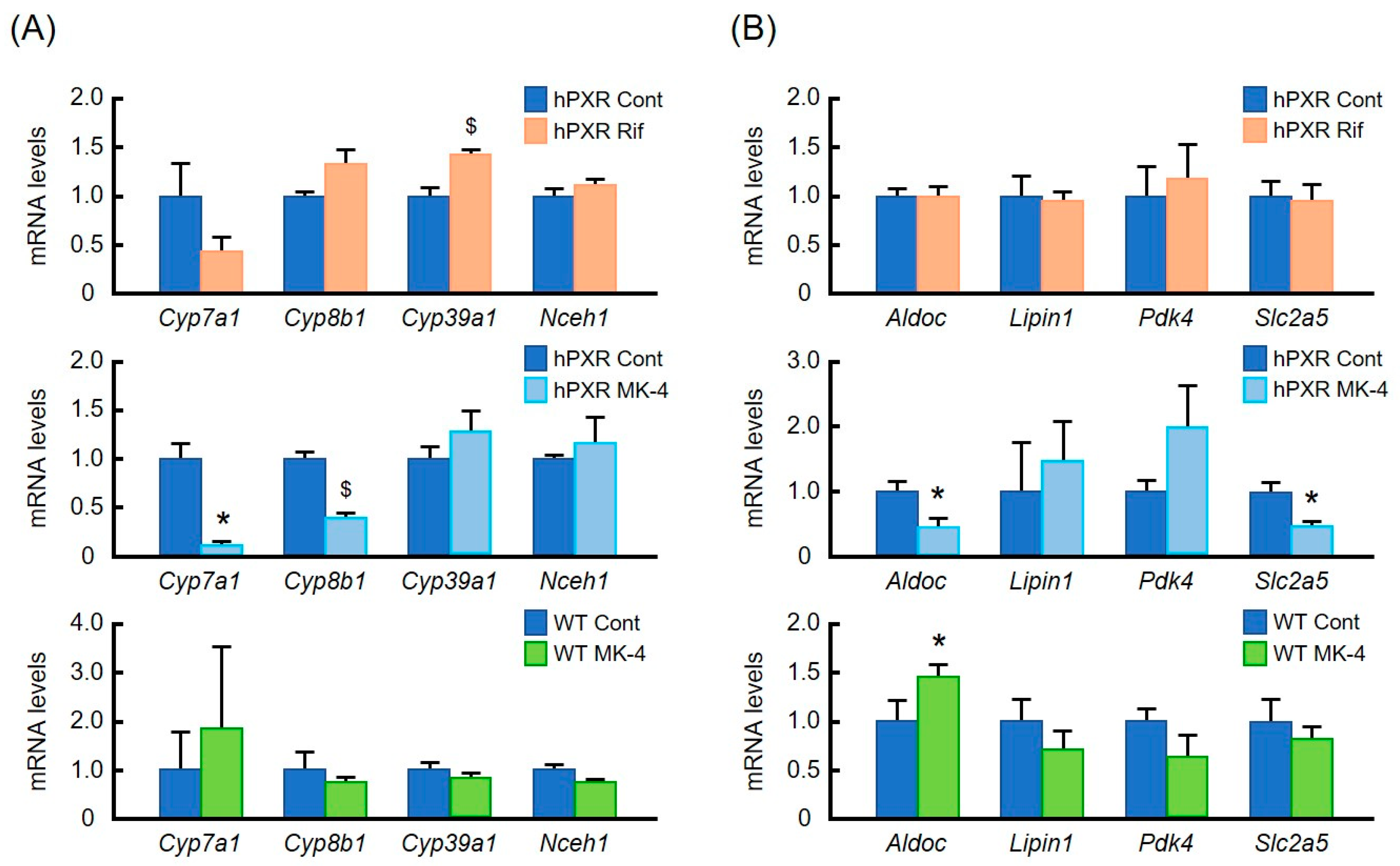
3.2. Effects of MK-4 and Rif on mRNA Levels of Typical PXR Target Genes
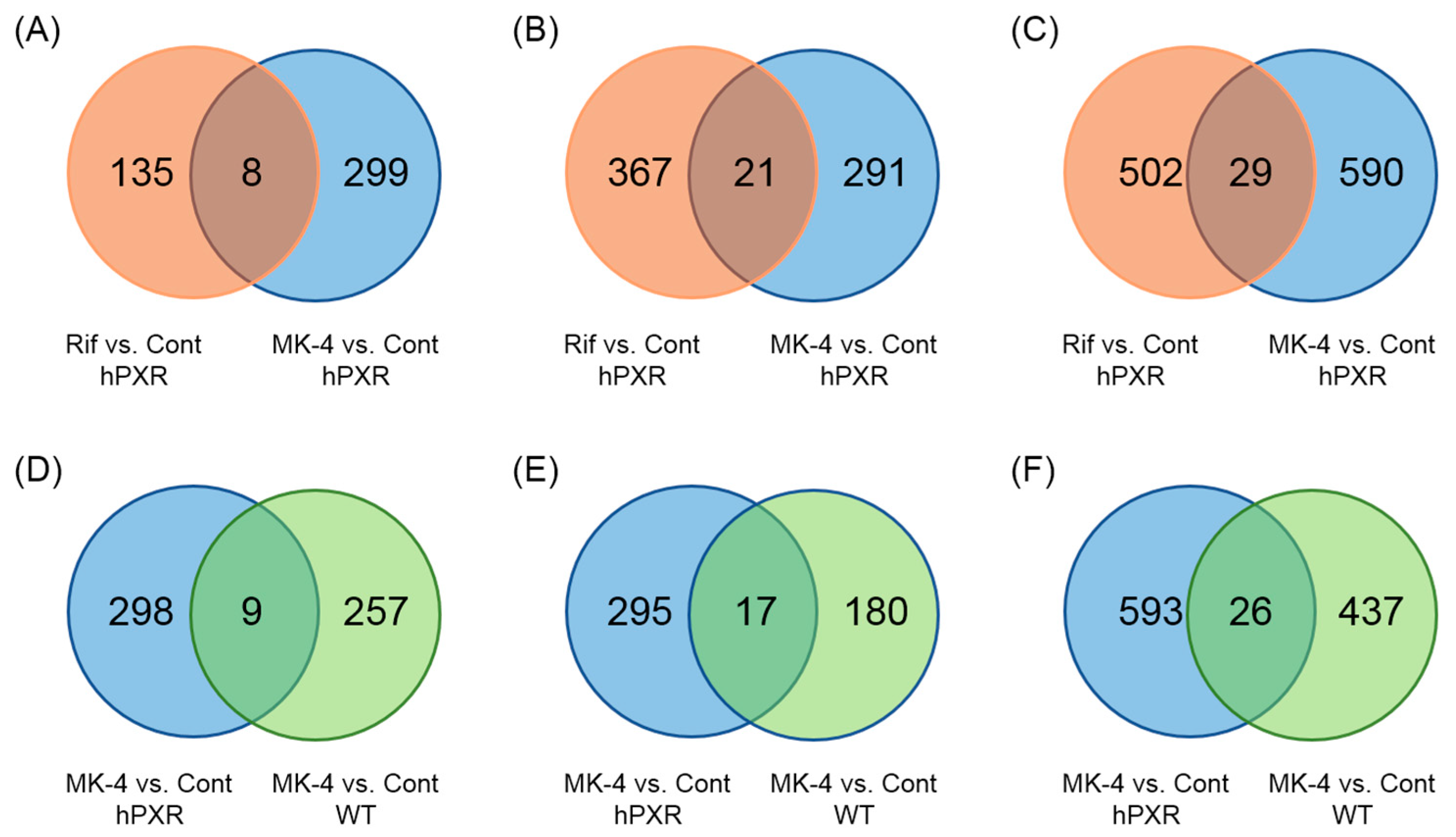
3.3. MK-4 and Rif Induced Different Changes in Gene Expression in hPXR and WT Mice
3.4. Validation of the Data Obtained from DNA Microarray Analysis by Quantitative RT-PCR
3.5. Effects of Lower Doses of MK-4 on Gene Expression
3.6. Effects of MK-4 on mRNA Levels of Bile Acid Synthesis Genes in HepG2 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kliewer, S.A.; Moore, J.T.; Wade, L.; Staudinger, J.L.; Watson, M.A.; Jones, S.A.; McKee, D.D.; Oliver, B.B.; Willson, T.M.; Zetterstrom, R.H.; et al. An Orphan Nuclear Receptor Activated by Pregnanes Defines a Novel Steroid Signaling Pathway. Cell 1998, 92, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Bertilsson, G.; Heidrich, J.; Svensson, K.; Asman, M.; Jendeberg, L.; Sydow-Backman, M.; Ohlsson, R.; Postlind, H.; Blomquist, P.; Berkenstam, A. Identification of a human nuclear receptor defines a new signaling pathway for CYP3A induction. Proc. Natl. Acad. Sci. USA 1998, 95, 12208–12213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blumberg, B.; Sabbagh, W., Jr.; Juguilon, H.; Bolado, J., Jr.; van Meter, C.M.; Ong, E.S.; Evans, R.M. SXR, a novel steroid and xenobiotic-sensing nuclear receptor. Genes Dev. 1998, 12, 3195–3205. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, K.; Kitajima, S.; Aisaki, K.; Tanemura, K.; Taquahashi, Y.; Moriyama, N.; Ikeno, E.; Matsuda, N.; Saga, Y.; Blumberg, B.; et al. Development of humanized steroid and xenobiotic receptor mouse by homologous knock-in of the human steroid and xenobiotic receptor ligand binding domain sequence. J. Toxicol. Sci. 2012, 37, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Cytochrome P-450 3A4: Regulation and role in drug metabolism. Ann. Rev. Pharmacol. Toxicol. 1999, 39, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Watkins, R.E.; Davis-Searles, P.R.; Lambert, M.H.; Redinbo, M.R. Coactivator Binding Promotes the Specific Interaction Between Ligand and the Pregnane X Receptor. J. Mol. Biol. 2003, 331, 815–828. [Google Scholar] [CrossRef]
- Watkins, R.E.; Wisely, G.B.; Moore, L.B.; Collins, J.L.; Lambert, M.H.; Williams, S.P.; Willson, T.M.; Kliewer, S.A.; Redinbo, M.R. The Human Nuclear Xenobiotic Receptor PXR: Structural Determinants of Directed Promiscuity. Science 2001, 292, 2329–2333. [Google Scholar] [CrossRef] [PubMed]
- Watkins, R.E.; Maglich, J.M.; Moore, L.B.; Wisely, G.B.; Noble, S.M.; Davis-Searles, P.R.; Lambert, M.H.; Kliewer, S.A.; Redinbo, M.R. 2.1 Å Crystal Structure of Human PXR in Complex with the St. John’s Wort Compound Hyperforin. Biochemistry 2003, 42, 1430–1438. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Verma, S.; Blumberg, B. The steroid and xenobiotic receptor (SXR), beyond xenobiotic metabolism. Nucl. Recept. Signal. 2009, 7, e001. [Google Scholar] [CrossRef] [PubMed]
- Staudinger, J.L.; Goodwin, B.; Jones, S.A.; Hawkins-Brown, D.; MacKenzie, K.I.; LaTour, A.; Liu, Y.; Klaassen, C.D.; Brown, K.K.; Reinhard, J.; et al. The nuclear receptor PXR is a lithocholic acid sensor that protects against liver toxicity. Proc. Natl. Acad. Sci. USA 2001, 98, 3369–3374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohsaki, Y.; Shirakawa, H.; Miura, A.; Giriwono, P.E.; Sato, S.; Ohashi, A.; Iribe, M.; Goto, T.; Komai, M. Vitamin K suppresses the lipopolysaccharide-induced expression of inflammatory cytokines in cultured macrophage-like cells via the inhibition of the activation of nuclear factor κB through the repression of IKKα/β phosphorylation. J. Nutr. Biochem. 2010, 21, 1120–1126. [Google Scholar] [CrossRef] [PubMed]
- Ito, A.; Shirakawa, H.; Takumi, N.; Minegishi, Y.; Ohashi, A.; Howlader, Z.H.; Ohsaki, Y.; Sato, T.; Goto, T.; Komai, M. Menaquinone-4 enhances testosterone production in rats and testis-derived tumor cells. Lipids Health Dis. 2011, 10, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamson, D.W.; Plaza, S.M. The Anticancer Effects of Vitamin K. Altern. Med. Rev. 2003, 8, 303–318. [Google Scholar]
- Tabb, M.M.; Sun, A.; Zhou, C.; Grun, F.; Errandi, J.; Romero, K.; Pham, H.; Inoue, S.; Mallick, S.; Lin, M.; et al. Vitamin K2 Regulation of Bone Homeostasis Is Mediated by the Steroid and Xenobiotic Receptor SXR. J. Biol. Chem. 2003, 278, 43919–43927. [Google Scholar] [CrossRef] [PubMed]
- Beulens, J.W.; van der A, D.L.; Grobbee, D.E.; Sluijs, I.; Spijkerman, A.M.; van der Schouw, Y.T. Dietary Phylloquinone and Menaquinones Intakes and Risk of Type 2 Diabetes. Diabetes Care 2010, 33, 1699–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neogi, T.; Booth, S.L.; Zhang, Y.Q.; Jacques, P.F.; Terkeltaub, R.; Aliabadi, P.; Felson, D.T. Low Vitamin K Status Is Associated With Osteoarthritis in the Hand and Knee. Arthritis Rheum. 2006, 54, 1255–1261. [Google Scholar] [CrossRef] [PubMed]
- Presse, N.; Shatenstein, B.; Kergoat, M.J.; Ferland, G. Low Vitamin K Intakes in Community-Dwelling Elders at an Early Stage of Alzheimer’s Disease. J. Am. Diet. Assoc. 2008, 108, 2095–2099. [Google Scholar] [CrossRef] [PubMed]
- Geleijnse, J.M.; Vermeer, C.; Grobbee, D.E.; Schurgers, L.J.; Knapen, M.H.; van der Meer, I.M.; Hofman, A.; Witteman, J.C. Dietary Intake of Menaquinone Is Associated with a Reduced Risk of Coronary Heart Disease: The Rotterdam Study. J. Nutr. 2004, 134, 3100–3105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azuma, K.; Inoue, S. Vitamin K, SXR, and GGCX. In Vitamin K2—Vital for Health and Wellbeing; InTechOpen: London, UK, 2017; pp. 21–32. ISBN 978-953-51-3020-8. [Google Scholar]
- Ichikawa, T.; Horie-Inoue, K.; Ikeda, K.; Blumberg, B.; Inoue, S. Steroid and Xenobiotic Receptor SXR Mediates Vitamin K2-activated Transcription of Extracellular Matrix-related Genes and Collagen Accumulation in Osteoblastic Cells. J. Biol. Chem. 2006, 281, 16927–16934. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Urano, T.; Ouchi, Y.; Inoue, S. Vitamin K2 Suppresses Proliferation and Motility of Hepatocellular Carcinoma Cells by Activating Steroid and Xenobiotic Receptor. Endocr. J. 2009, 56, 843–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, S.A.; Moore, L.B.; Shenk, J.L.; Wisely, G.B.; Hamilton, G.A.; McKee, D.D.; Tomkinson, N.C.; LeCluyse, E.L.; Lambert, M.H.; Willson, T.M.; et al. The Pregnane X Receptor: A Promiscuous Xenobiotic Receptor That Has Diverged during evolution. Mol. Endocrinol. 2000, 1, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Barwick, J.L.; Downes, M.; Blumberg, B.; Simon, C.M.; Nelson, M.C.; Neuschwander-Tetri, B.A.; Brunt, E.M.; Guzelian, P.S.; Evans, R.M. Humanized xenobiotic response in mice expressing nuclear receptor SXR. Nature 2000, 406, 435–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Shah, Y.; Cheung, C.; Guo, G.L.; Feigenbaum, L.; Krausz, K.W.; Idle, J.R.; Gonzalez, F.J. The Pregnane X Receptor Gene-Humanized mouse: A Model for Investigating drug-drug Interactions Mediated by Cytochromes P450 3A. Drug Metab. Dispos. 2007, 35, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Scheer, N.; Ross, J.; Rode, A.; Zevnik, B.; Niehaves, S.; Faust, N.; Wolf, C.R. A novel panel of mouse models to evaluate the role of human pregnane X receptor and constitutive androstane receptor in drug response. J. Clin. Investig. 2008, 118, 3228–3239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohsaki, Y.; Shirakawa, H.; Hiwatashi, K.; Furukawa, Y.; Mizutani, T.; Komai, M. Vitamin K Suppresses Lipopolysaccharide Induced Inflammation in the Rat. Biosci. Biotechnol. Biochem. 2006, 70, 926–932. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Shirakawa, H.; Tomita, S.; Tohkin, M.; Gonzalez, F.J.; Komai, M. The aryl hydrocarbon receptor and glucocorticoid receptor interact to activate human metallothionein 2A. Toxicol. Appl. Pharmacol. 2013, 273, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Mitro, N.; Godio, C.; De Fabiani, E.; Scotti, E.; Galmozzi, A.; Gilardi, F.; Caruso, D.; Chacon, A.B.V.; Crestani, M. Insights in the Regulation of Cholesterol 7α-Hydroxylase Gene Reveal a Target for Modulating Bile Acid Synthesis. Hepatology 2007, 46, 885–897. [Google Scholar] [CrossRef] [PubMed]
- Masuyama, H.; Hiramatsu, Y.; Kunitomi, M.; Kudo, T.; MacDonald, P.N. Endocrine Disrupting Chemicals, Phthalic Acid and Nonylphenol, Activate Pregnane X Receptor-Mediated Transcription. Mol. Endocrinol. 2000, 14, 421–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavek, P. Pregnane X Receptor (PXR)-Mediated Gene Repression and Cross-Talk of PXR with Other Nuclear Receptors via Coactivator Interactions. Front. Pharmacol. 2016, 7, 456. [Google Scholar] [CrossRef] [PubMed]
- Tolson, A.H.; Wang, H. Regulation of drug-metabolizing enzymes by xenobiotic receptors: PXR and CAR. Adv. Drug Deliv. Rev. 2010, 62, 1238–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benson, E.A.; Eadon, M.T.; Desta, Z.; Liu, Y.; Lin, H.; Burgess, K.S.; Segar, M.W.; Gaedigk, A.; Skaar, T.C. Rifampin Regulation of Drug Transporters Gene Expression and the Association of MicroRNAs in Human Hepatocytes. Front. Pharmacol. 2016, 7, 111. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Robertson, L.W.; Wang, K.; Ludewig, G. Species difference in the regulation of cytochrome P450 2S1: Lack of induction in rats by the aryl hydrocarbon receptor agonist PCB126. Xenobiotica 2011, 41, 1031–1043. [Google Scholar] [CrossRef] [PubMed]
- Maglich, J.M.; Stoltz, C.M.; Goodwin, B.; Hawkins-Brown, D.; Moore, J.T.; Kliewer, S.A. Nuclear Pregnane X Receptor and Constitutive Androstane Receptor Regulate Overlapping but Distinct Sets of Genes Involved in Xenobiotic Detoxification. Mol. Pharmacol. 2002, 62, 638–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, J.Y. Bile acids: Regulation of synthesis. J. Lipid Res. 2009, 50, 1955–1966. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chiang, J.Y. Mechanism of rifampicin and pregnane X receptor inhibition of human cholesterol 7α-hydroxylase gene transcription. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G74–G84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhalla, S.; Ozalp, C.; Fang, S.; Xiang, L.; Kemper, J.K. Ligand-activated Pregnane X Receptor Interferes with HNF-4 Signaling by Targeting a Common Coactivator PGC-1α. Functional implications in hepatic cholesterol and glucose metabolism. J. Biol. Chem. 2004, 279, 45139–45147. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.; Alhousseini, A.; Awadalla, M.; Idler, J.; Welch, R.; Puder, K.; Patwardhan, M.; Gonik, B. Intrahepatic Cholestasis of Pregnancy Leading to Severe Vitamin K Deficiency and Coagulopathy. Case Rep. Obstet. Gynecol. 2017, 2017, 5646247. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Gao, J.; Xie, W. PXR and CAR in energy metabolism. Trends Endocrinol. Metab. 2009, 20, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhai, Y.; Mu, Y.; Gong, H.; Uppal, H.; Toma, D.; Ren, S.; Evans, R.M.; Xie, W. A Novel Pregnane X Receptor-mediated and Sterol Regulatory Element-binding Protein-independent Lipogenic Pathway. J. Biol. Chem. 2006, 281, 15013–15020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakkola, J.; Rysa, J.; Hukkanen, J. Regulation of hepatic energy metabolism by the nuclear receptor PXR. Biochim. Biophys. Acta 2016, 1859, 1072–1082. [Google Scholar] [CrossRef] [PubMed]
- Prasenjit Manna, P.; Kalita, J. Beneficial role of vitamin K supplementation on insulin sensitivity, glucose metabolism, and the reduced risk of type 2 diabetes: A review. Nutrition 2016, 32, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Rutter, W.J. Evolution of Aldolase. Fed. Proc. 1964, 23, 1248–1257. [Google Scholar] [PubMed]
- Burant, C.F.; Takeda, J.; Brot-Laroche, E.; Bell, G.I.; Davidson, N.O. Fructose Transporter in Human Spermatozoa and Small Intestine is GLUT5. J. Biol. Chem. 1992, 267, 14523–14526. [Google Scholar] [PubMed]
- Hartley, D.P.; Dai, X.; He, Y.D.; Carlini, E.J.; Wang, B.; Huskey, S.W.; Ulrich, R.G.; Rushmore, T.H.; Evers, R.; Evans, D.C. Activators of the Rat Pregnane X Receptor Differentially Modulate Hepatic and Intestinal Gene Expression. Mol. Pharmacol. 2004, 65, 1159–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Forward Primer | Reverse Primer |
|---|---|---|
| Abca3 | TCCTTTGCCTTCATAGCACAGTT | GACCAAGCCAGGGATGCTAA |
| Aldh1b1 | TCAGCCTGAGTTCCCAGTGA | GGTTGACACTTCTTGTAACACTTCA |
| Aldoc | TCATCTCACATCTCACATGCCTT | GGTCTCGGGTACAGCAATGT |
| Ces2a | GTATCGCTTGGGGGTCCTTG | GCCACTTGGTCCAGGTATCC |
| Cyp1a1 | TGCATCAGCTCTTGGTCTCTC | CAAATGCATAAGCAAAATACAGTCCAT |
| Cyp2s1 | AGCTGACCTCTTTGTGACTTGA | TTTATTGCAAAACGGGAACCTTCA |
| Cyp39a1 | GAAGGTGGGGAACGGAAACT | CCTGGCACAAAACCAGATGC |
| Cyp3a11 | CCTGGGTGCTCCTAGCAATC | GGCCCAGGAATTCCCTGTTT |
| Cyp7a1 | AAAACAAGTTTTATGACTCCCTGAAC | GGAAGGTATGTGGATACATTCAGTT |
| Cyp8b1 | AAACCTGGAAAAAGACGGCATAAG | ACTGAAGCATGTAGCCTAACCAA |
| Eef1al | GATGGCCCCAAATTCTTGAAG | GGACCATGTCAATGGCAG |
| Gstp1 | CAGGGCCAGAGCTGGAAG | AGCCTTGCATCCAGGTATCTATG |
| Lipin1 | TGGGTACCATACATTTCAAAGTTGC | CTATGGAGCAAGTCGCTCATTTC |
| Mdr1 | ATGCTGAGACAGGATGTGAGC | AGACCCTGTAGCCCCTTTCA |
| Nceh1 | TTTTTCATATGTGTCCATGTCTGGG | ACCAGTACTCATATGCACATACCC |
| Pdk4 | ATGAACCCATGGGAGACTTTAA | GCCTGGGCATTTAGCATCTAT |
| Slc2a5 | CCATTTGCGAAGACACACTGAG | CGCTTACAGTTAATAATCCACGCTC |
| Sult1b1 | GACCCACAATCTGATACAACCTC | AAATTAGTCTAGGTCACAGATGCTT |
| hCYP3A4 | TGGTGATGATTCCAAGCTATGCT | AATGCAGTTTCTGGGTCCACTTC |
| hCYP7A1 * | AAACGGGTGAACCACCTCTAGA | AACTCAAGAGGATTGGCACCA |
| hCYP8B1 | AAACGGGTGAACCACCTCTAGA | AACTCAAGAGGATTGGCACCA |
| hMDR1 | CCCATCATTGCAATAGCAGG | TGTTCAAACTTCTGCTCCTGA |
| Groups | Body Weight (g) | Liver Weight (g) | Ratio (Liver/Body Weight) | |
|---|---|---|---|---|
| Wild type | Control | 19.7 ± 0.440 * | 0.740 ± 0.0386 | 0.0375 ± 0.00160 |
| 10 mg/kg BW | 20.1 ± 0.614 | 0.7875± 0.0175 | 0.0392 ± 0.00072 | |
| 50 mg/kg BW | 20.6 ± 0.795 | 0.770 ± 0.0409 | 0.0377 ± 0.00169 | |
| 100 mg/kg BW | 19.4 ± 0.290 | 0.807 ± 0.0169 | 0.0417 ± 0.00030 | |
| hPXR | Control | 18.0 ± 0.422 | 0.776 ± 0.0281 | 0.0436 ± 0.0025 |
| 10 mg/kg BW | 17.3 ± 0.414 | 0.798 ± 0.0253 | 0.0461 ± 0.00081 | |
| 50 mg/kg BW | 17.4 ± 0.232 | 0.808 ± 0.0317 | 0.0465 ± 0.00218 | |
| 100 mg/kg BW | 18.7 ± 0.396 | 0.822 ± 0.0348 | 0.0439 ± 0.00091 | |
| Function | Gene Symbol | WT MK-4 * | hPXR MK-4 | hPXR Rif |
|---|---|---|---|---|
| Bile homeostasis | Cyp7a1 | 2.24 | 0.07 | 0.39 |
| Cyp8b1 | 1.11 | 0.45 | 0.99 | |
| Cyp39a1 | 0.93 | 1.82 | 1.12 | |
| Nceh1 | 0.97 | 1.83 | 1.91 | |
| Energy homeostasis | Aldoc | 1.44 | 0.44 | 0.73 |
| Lipin1 | 1.03 | 2.00 | 0.75 | |
| Pdk4 | 0.61 | 2.12 | 0.96 | |
| Slc2a5 | 0.99 | 0.51 | 0.71 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sultana, H.; Watanabe, K.; Rana, M.M.; Takashima, R.; Ohashi, A.; Komai, M.; Shirakawa, H. Effects of Vitamin K2 on the Expression of Genes Involved in Bile Acid Synthesis and Glucose Homeostasis in Mice with Humanized PXR. Nutrients 2018, 10, 982. https://doi.org/10.3390/nu10080982
Sultana H, Watanabe K, Rana MM, Takashima R, Ohashi A, Komai M, Shirakawa H. Effects of Vitamin K2 on the Expression of Genes Involved in Bile Acid Synthesis and Glucose Homeostasis in Mice with Humanized PXR. Nutrients. 2018; 10(8):982. https://doi.org/10.3390/nu10080982
Chicago/Turabian StyleSultana, Halima, Kimika Watanabe, Md Masud Rana, Rie Takashima, Ai Ohashi, Michio Komai, and Hitoshi Shirakawa. 2018. "Effects of Vitamin K2 on the Expression of Genes Involved in Bile Acid Synthesis and Glucose Homeostasis in Mice with Humanized PXR" Nutrients 10, no. 8: 982. https://doi.org/10.3390/nu10080982
APA StyleSultana, H., Watanabe, K., Rana, M. M., Takashima, R., Ohashi, A., Komai, M., & Shirakawa, H. (2018). Effects of Vitamin K2 on the Expression of Genes Involved in Bile Acid Synthesis and Glucose Homeostasis in Mice with Humanized PXR. Nutrients, 10(8), 982. https://doi.org/10.3390/nu10080982