Bioavailability of Vitamin B12 from Dairy Products Using a Pig Model


Abstract
:1. Introduction
2. Methods
3. Initial Analysis and Selection of Dairy Products
4. Preliminary Animal Trial
5. Description of Treatments
6. Experimental Animals and Palatability Test
7. Surgery
8. Post-Operatory Procedures and Experimental Days
9. Sample Handling and Analyses
10. Calculations and Statistical Analysis
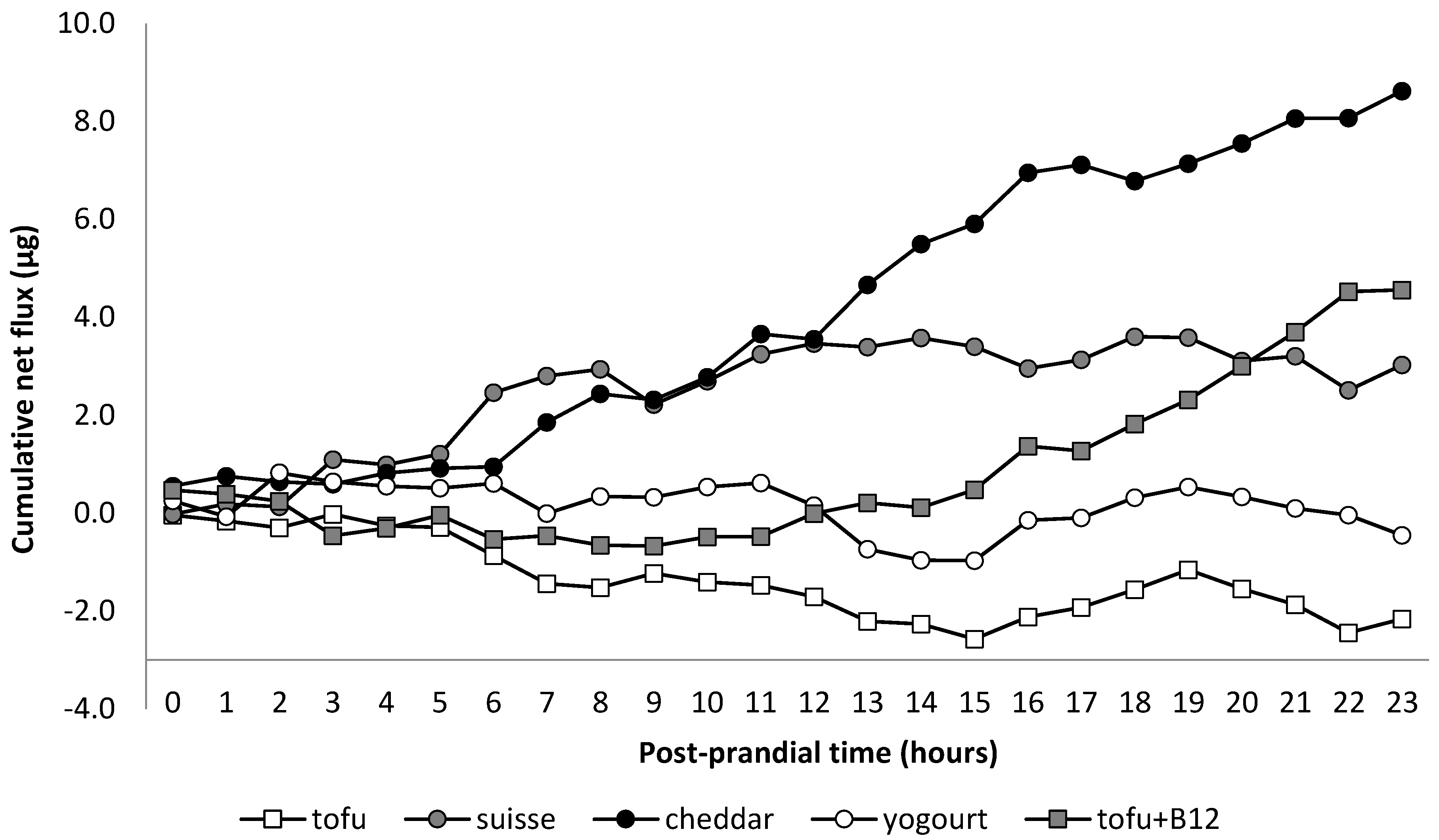
11. Results
12. Discussion
13. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miller, D.R.; Specker, B.L.; Ho, M.L.; Norman, E.J. Vitamin B12 status in a macrobiotic community. Am. J. Clin. Nutr. 1991, 53, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Tucker, K.L.; Rich, S.; Rosenberg, I.H.; Jacques, P.; Dallal, G.; Wilson, P.W.; Selhub, J. Plasma vitamin B12 concentrations relate to intake source in the Framingham offspring study. Am. J. Clin. Nutr. 2000, 71, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Vogiatzoglou, A.; Smith, A.D.; Nurk, E.; Berstad, P.; Drevon, C.A.; Ueland, P.M.; Vollset, S.E.; Tell, G.S.; Refsum, H. Dietary sources of vitamin B12 and their association with plasma vitamin B12 concentrations in the general population: The Hordaland Homocysteine Study. Am. J. Clin. Nutr. 2009, 89, 1078–1087. [Google Scholar] [CrossRef] [PubMed]
- Matte, J.J.; Guay, F.; Christiane, L. Bioavailability of vitamin B12 in cows’ milk. Brit. J. Nutr. 2012, 107, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Farquharson, J.; Adams, J.F. The forms of vitamin B12 in foods. Br. J. Nutr. 1976, 36, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Ball, G.F.M. Vitamins in Foods: Analysis, Bioavailability, and Stability; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Gardner, N.; Champagne, C.P. Production of Propionibacterium shermanii biomass and vitamin B12 on spent media. J. Appl. Microbiol. 2005, 99, 1236–1245. [Google Scholar] [CrossRef] [PubMed]
- Arkbage, K.; Witthoft, C.; Fondén, R.; Jägerstad, M. Retention of vitamin B12 during manufacture of six fermented dairy products using a validated radio protein-binding assay. Int. Dairy J. 2003, 13, 101–109. [Google Scholar] [CrossRef]
- Canadian Council on Animal Care. Guide to the Care and Use of Experimental Animals; Canadian Council on Animal Care: Ottawa, ON, Canada, 2009. [Google Scholar]
- National Farm Animal Care Council. Code of Practice for the Care and Handling of Pigs; Agriculture Canada: Ottawa, ON, Canada, 2014.
- Martens, J.-H.; Barg, H.; Warren, M.J.; Jahn, D. Microbial production of vitamin B12. Appl. Microbiol. Biotechnol. 2002, 58, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Matte, J.J.; Guay, F.; Le Floc’h, N.; Girard, C.L. Bioavailability of dietary cyanocobalamin (vitamin B12) in growing pigs. J. Anim. Sci. 2010, 88, 3936–3944. [Google Scholar] [CrossRef] [PubMed]
- Flohr, J.R.; DeRouchey, J.M.; Woodworth, J.C.; Tokach, M.D.; Goodband, R.D.; Dritz, S.S. A survey of current feeding regimens for vitamins and trace minerals in the US swine industry. J. Swine Health Prod. 2016, 24, 290–303. [Google Scholar]
- Hooda, S.; Matte, J.J.; Wilkinson, C.W.; Zijlstra, R.T. Technical note: An improved surgical model for the long-term studies of kinetics and quantification of nutrient absorption in swine. J. Anim. Sci. 2009, 87, 2013–2019. [Google Scholar] [CrossRef] [PubMed]
- Manet, L. Techniques usuelles de biologie clinique. In Hématologie; Editions Medicales Flammarion: Paris, France, 1969. [Google Scholar]
- Girard, C.L.; Lapierre, H.; Desrochers, A.; Benchaar, C.; Matte, J.J.; Rémond, D. Net flux of folates and vitamin B12 through the gastrointestinal tract and the liver of lactating dairy cows. Br. J. Nutr. 2001, 86, 707–715. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute. SAS/STAT User’s Guide; SAS Inst. Inc.: Cary, NC, USA, 2004. [Google Scholar]
- Guilloteau, P.; Zabielski, R.; Hammon, H.M.; Metges, C.C. Nutritional programming of gastrointestinal tract development. Is pig a good model for man? Nutr. Res. Rev. 2010, 23, 4–22. [Google Scholar] [CrossRef] [PubMed]
- Schneider, Z.; Stroinski, A. Comprehensive B12: Chemistry, Biochemistry, Nutrition, Ecology, Medicine; Walter de Gruyter: Berlin, Germany, 1987. [Google Scholar]
- Combs, G.F., Jr. The Vitamins: Fundamental Aspects in Nutrition and Health, 4th ed.; Academic Press: San Diego, CA, USA, 2012. [Google Scholar]
- Wardener, H.E.; He, F.; Macgregor, G.A. Plasma sodium and hypertension. Kidney Int. 2004, 66, 2454–2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siregar, H.; Chou, C.C. Relative contribution of fat, protein, carbohydrate, and ethanol to intestinal hyperemia. Am. J. Physiol. 1982, 242, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.C.; Coatney, R.W. Nutrient-induced changes in intestinal blood flow in the dog. Br. Vet. J. 1994, 150, 423–437. [Google Scholar] [CrossRef]
- Greibe, E. Nutritional and biochemical aspects of cobalamin throughout life. In Vitamin B12: Advances and Insights; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Artegoitia, V.M.; De Veth, M.J.; Harte, F.; Ouellet, D.R.; Girard, C.L. Casein hydrolysate and whey proteins as excipients for cyancobalamin to increase intestinal absorption in the lactating dairy cow. J. Dairy Sci. 2015, 98, 8128–8132. [Google Scholar] [CrossRef] [PubMed]
- Burn-Murdoch, R.A.; Fisher, M.A.; Hunt, J.H. The slowing of gastric emptying by proteins in test meals. J. Physiol. 1978, 274, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Gizis, E.; Kim, Y.P.; Brunner, J.R.; Schweigert, B.S. Vitamin B12 content and binding capacity of cow’s milk proteins. J. Nutr. 1965, 87, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Dalziel, J.E.; Young, W.; McKenzie, C.M.; Haggarty, N.W.; Roy, N.C. Gastric emptying and gastrointestinal transit compared among native and hydrolyzed whey and casein milk proteins in an aged rat model. Nutrients 2017, 9, 1351. [Google Scholar] [CrossRef] [PubMed]
- Bender, D.A. Nutritional Biochemistry of the Vitamins; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Adams, J.F.; Ross, S.K.; Mervyn, L.; Boddy, K.; King, P. Absorption of cyanocobalamin, coenzyme B12, methylcobalamin, and hydroxocobalamin at different dose levels. Scand. J. Gastroenter. 1971, 6, 249–252. [Google Scholar] [CrossRef]
- Webb, B.H.; Johnson, A.H.; Alford, J.A. Fundamentals of Dairy Chemistry, 2nd ed.; Avi Publishing CO Inc.: Westport, MA, USA, 1974. [Google Scholar]
- Jalan, K.N.; Mahalanabis, D.; Maitra, T.K.; Agarwal, S.K. Gastric acid secretion rate and buffer content of the stomach after a rice and a wheat-based meal in normal subjects and patients with duodenal ulcer. Gut 1979, 20, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Fedosov, S.N.; Nexo, E.; Heegaard, C.W. Binding of aquocobalamin to bovine casein and its peptides via coordination to histidine residues. Int. Dairy J. 2018, 76, 30–39. [Google Scholar] [CrossRef]
- Rioux, L.E.; Turgeon, S.L. The ratio of casein to whey protein impacts yogurt digestion in vitro. Food Dig. 2012, 3, 25–35. [Google Scholar] [CrossRef]
- Richardson, B.C.; Pearce, K.N. The determination of plasmin in dairy products. J. Dairy Sci. Technol. 1981, 16, 209–220. [Google Scholar]


{kind=link}
| Item | Tofu | Swiss Cheese | Cheddar Cheese | Yogurt 2 |
|---|---|---|---|---|
| Composition | ||||
| Dry matter, % | 34.60 | 62.40 | 52.90 | 12.70 (23.7) |
| Protein, g/g | 0.17 | 0.27 | 0.23 | 0.06 (0.11) |
| Fat, g/g | 0.05 | 0.27 | 0.33 | 0.02 (0.04) |
| Sodium, mg/g | 0.10 | 5.33 | 5.00 | 0.49 (0.91) |
| Vitamin B12, ng/g | 0.12 | 31.88 | 14.87 | 3.77 (6.79) |
| Calculated provision per meal | ||||
| Dry matter, g | 692.0 | 833.0 | 883.0 | 865.1 |
| Protein, g | 340.0 | 380.6 | 384.1 | 401.5 |
| Fat, g | 100.0 | 260.6 | 551.1 | 146.0 |
| Sodium, g | 0.2 | 4.26 | 8.35 | 3.32 |
| Vitamin B12, ng | 0.2 | 25.1 | 24.8 | 24.8 |
| Item | Tofu | Swiss Cheese | Cheddar Cheese | Yogurt | Tofu + B12 | p Value |
|---|---|---|---|---|---|---|
| Arterial B12, ng/L | 173.2 ± 14.2 | 177.2 ± 13.0 | 145.4 ± 143.0 | 187.7 ± 15.2 | 194.6 ± 16.4 | 0.18 |
| PDV plasma flow, L/min | 0.93 c ± 0.08 | 1.31 a ± 0.08 | 1.34 a ± 0.08 | 1.19 ab ± 0.09 | 1.06 bc ± 0.08 | 0.01 |
| Porto-arterial difference, ng/L 1 | −1.36 b ± 1.56 | 1.58 ab ± 1.46 | 4.68 a ± 1.53 | −0.21 b ± 1.68 | 4.78 a ± 1.81 | 0.03 |
| Net PDV flux of B12, ng/min 2,3 | −1.50 c ± 1.84 | 2.10 abc ± 1.73 | 5.99 a ± 1.81 | −0.31 bc ± 1.98 | 3.17 ab ± 2.14 | 0.06 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bueno Dalto, D.; Audet, I.; Girard, C.L.; Matte, J.-J. Bioavailability of Vitamin B12 from Dairy Products Using a Pig Model. Nutrients 2018, 10, 1134. https://doi.org/10.3390/nu10091134
Bueno Dalto D, Audet I, Girard CL, Matte J-J. Bioavailability of Vitamin B12 from Dairy Products Using a Pig Model. Nutrients. 2018; 10(9):1134. https://doi.org/10.3390/nu10091134
Chicago/Turabian StyleBueno Dalto, Danyel, Isabelle Audet, Christiane L. Girard, and Jean-Jacques Matte. 2018. "Bioavailability of Vitamin B12 from Dairy Products Using a Pig Model" Nutrients 10, no. 9: 1134. https://doi.org/10.3390/nu10091134
APA StyleBueno Dalto, D., Audet, I., Girard, C. L., & Matte, J. -J. (2018). Bioavailability of Vitamin B12 from Dairy Products Using a Pig Model. Nutrients, 10(9), 1134. https://doi.org/10.3390/nu10091134