Protection of UVB-Induced Photoaging by Fuzhuan-Brick Tea Aqueous Extract via MAPKs/Nrf2-Mediated Down-Regulation of MMP-1



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
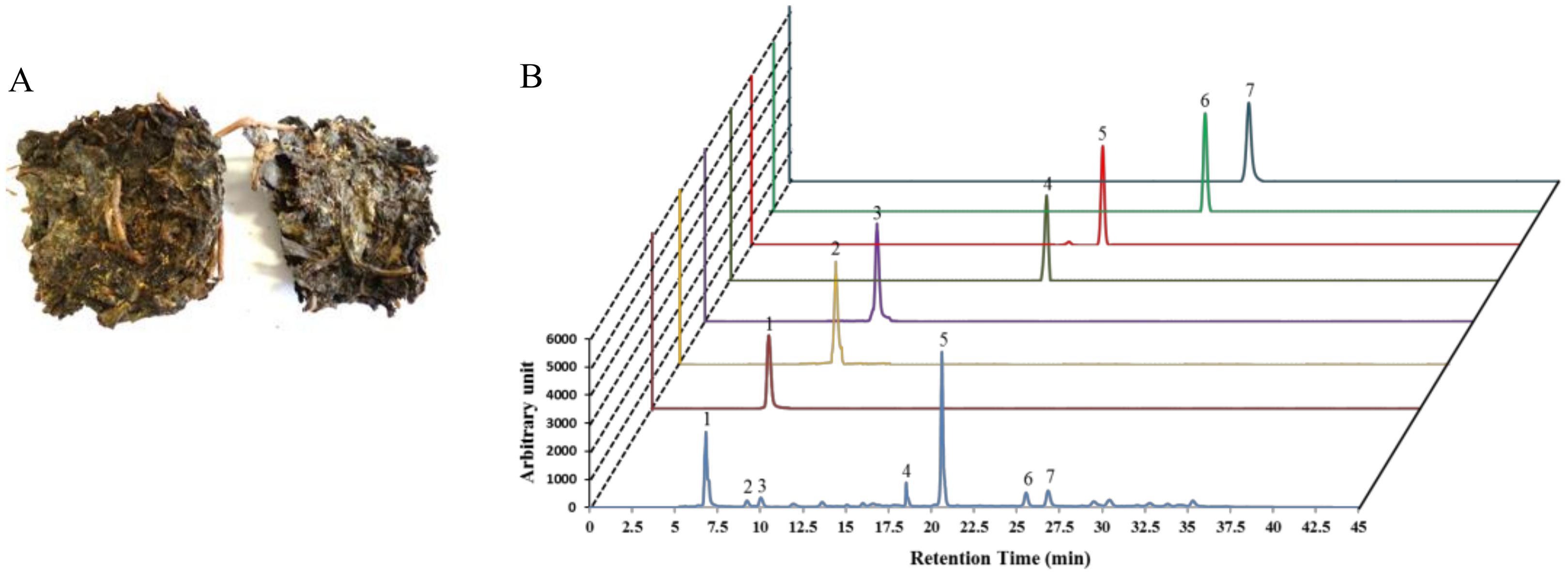
2.1. Plant Materials and Extraction
2.2. Radical-Scavenging Activity Assays
2.3. Cell Culture, UVB-Irradiation and Cell Viability Assay
2.4. Measurement of Cellular ROS Generation
2.5. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
2.6. Cell Lysates and Western Blotting
2.7. Statistical Analysis
3. Results
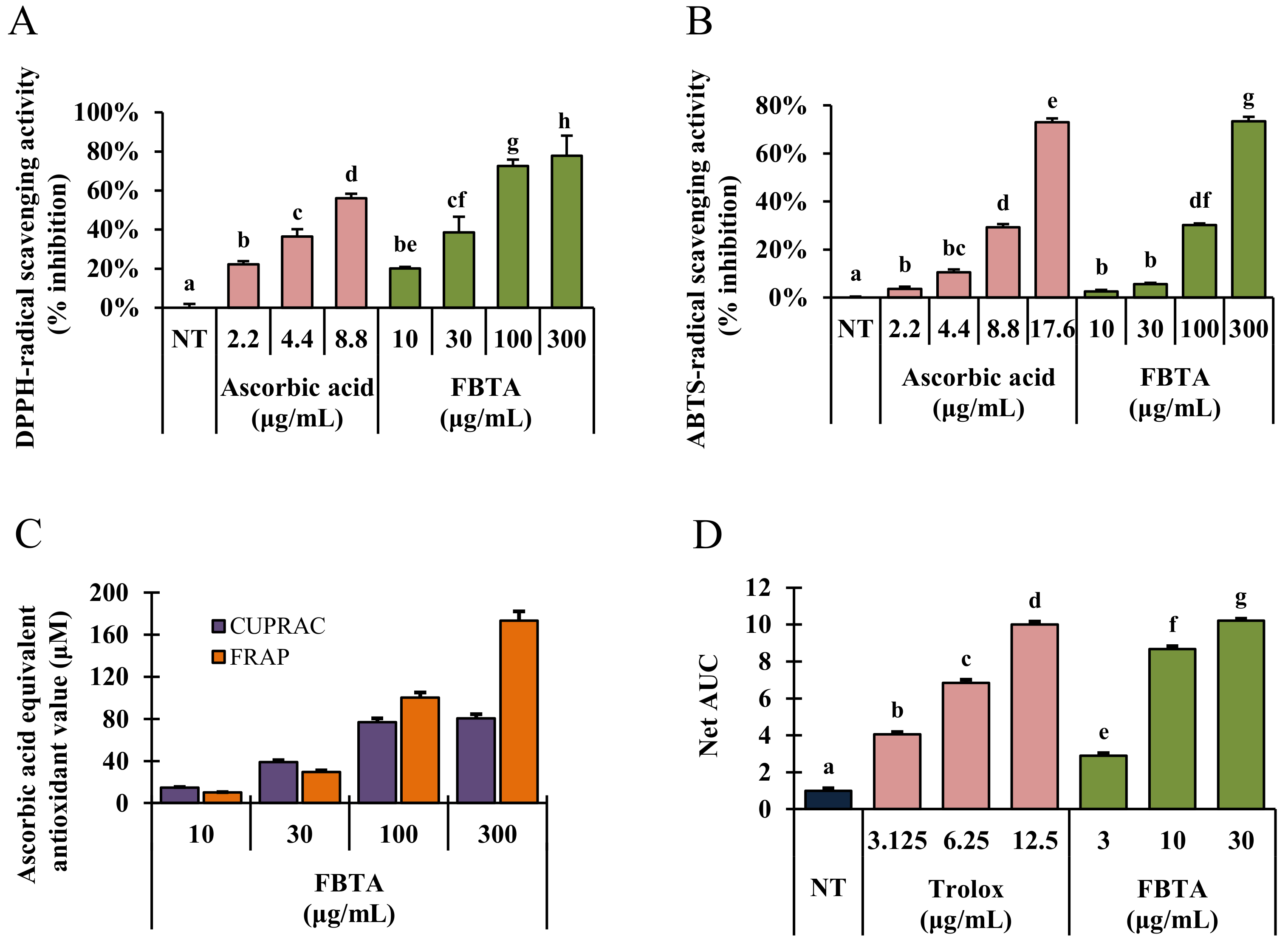
3.1. Radical Scavenging Abilities
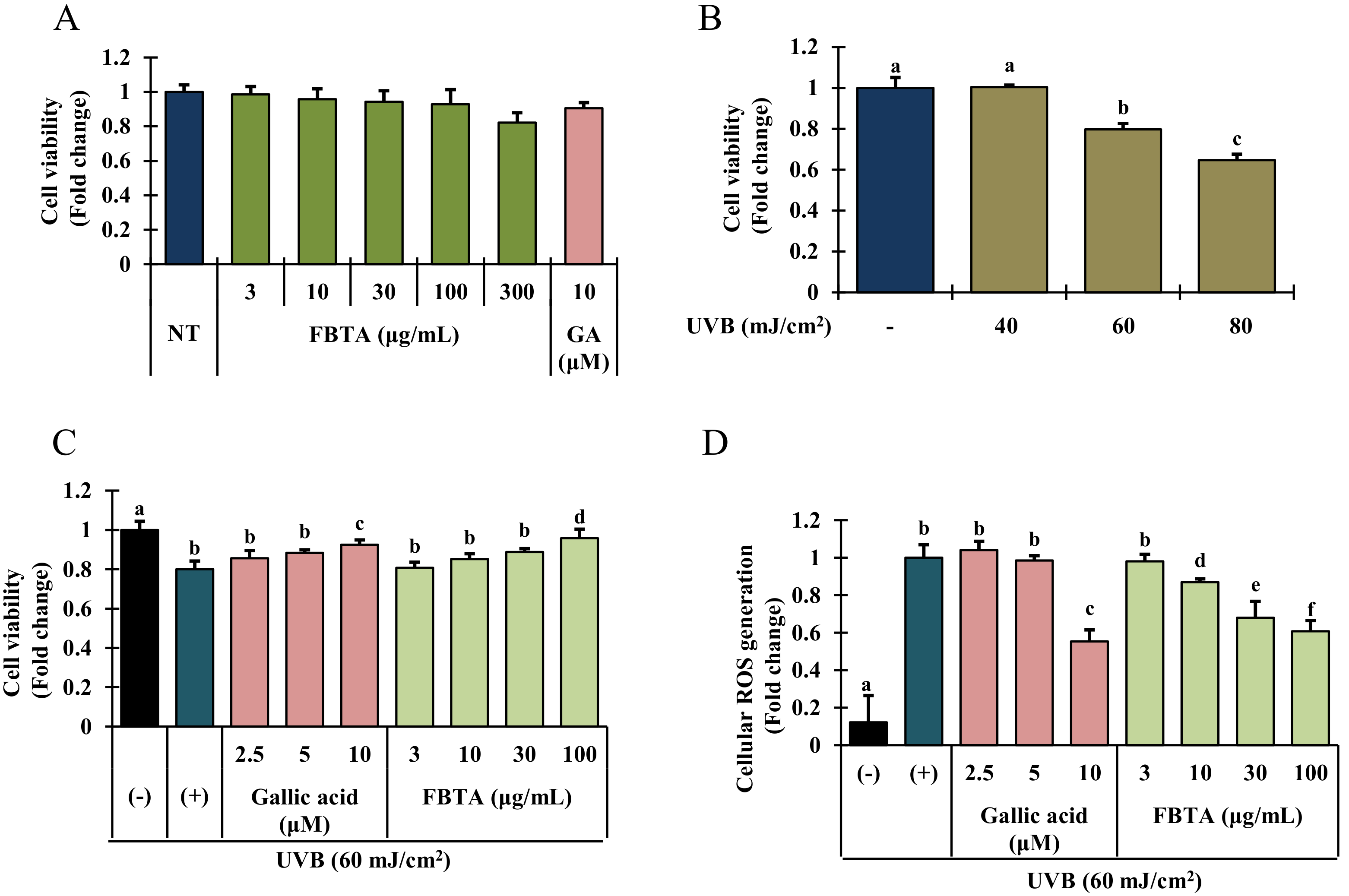
3.2. Assay of Cell Viability in UVB-Irradiated HaCaT Cells
3.3. Effects of FBTA on ROS Generation
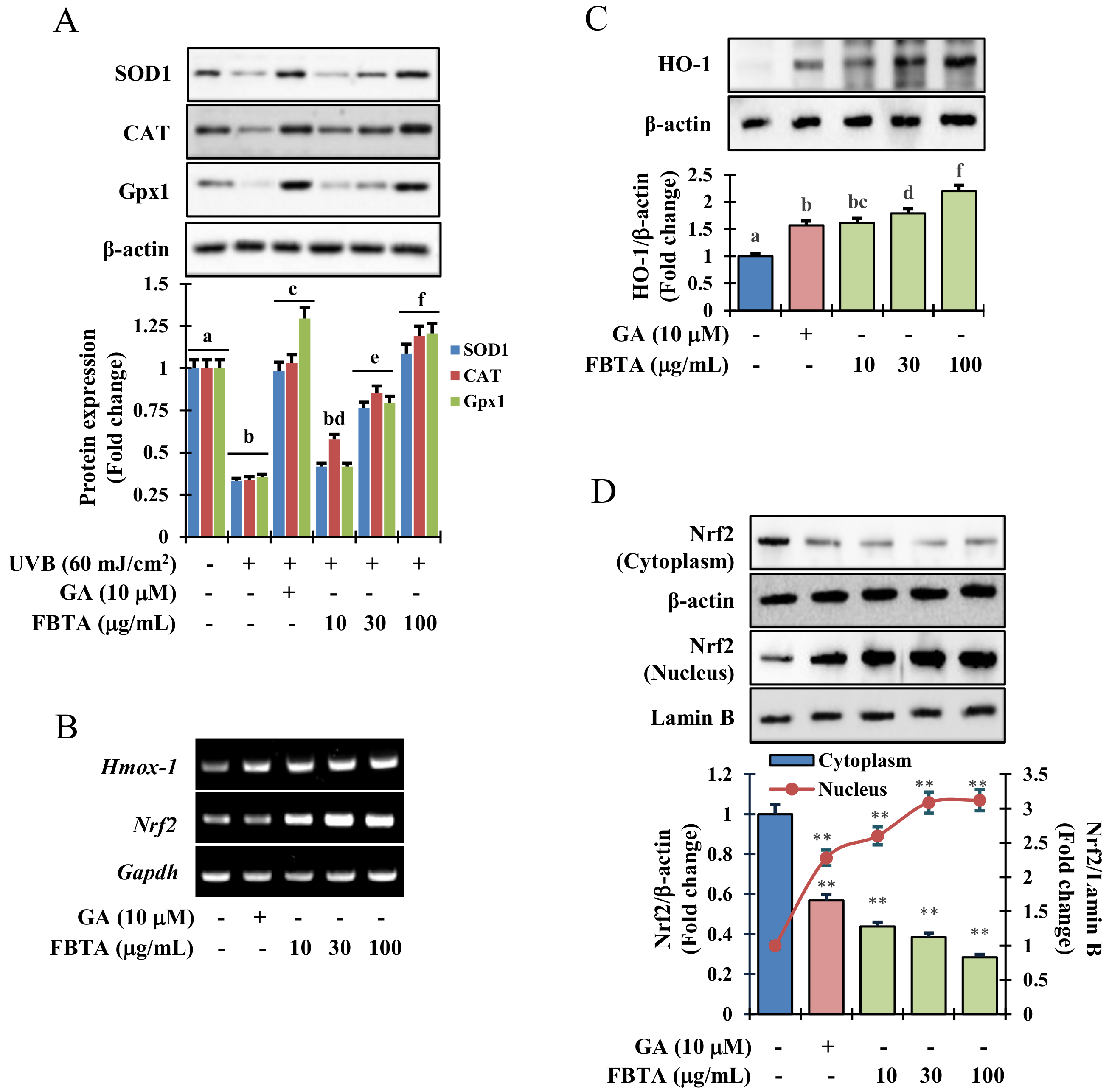
3.4. Effects of FBTA on Phase I and Phase II Antioxidant Enzyme Expression in HaCaT Cells
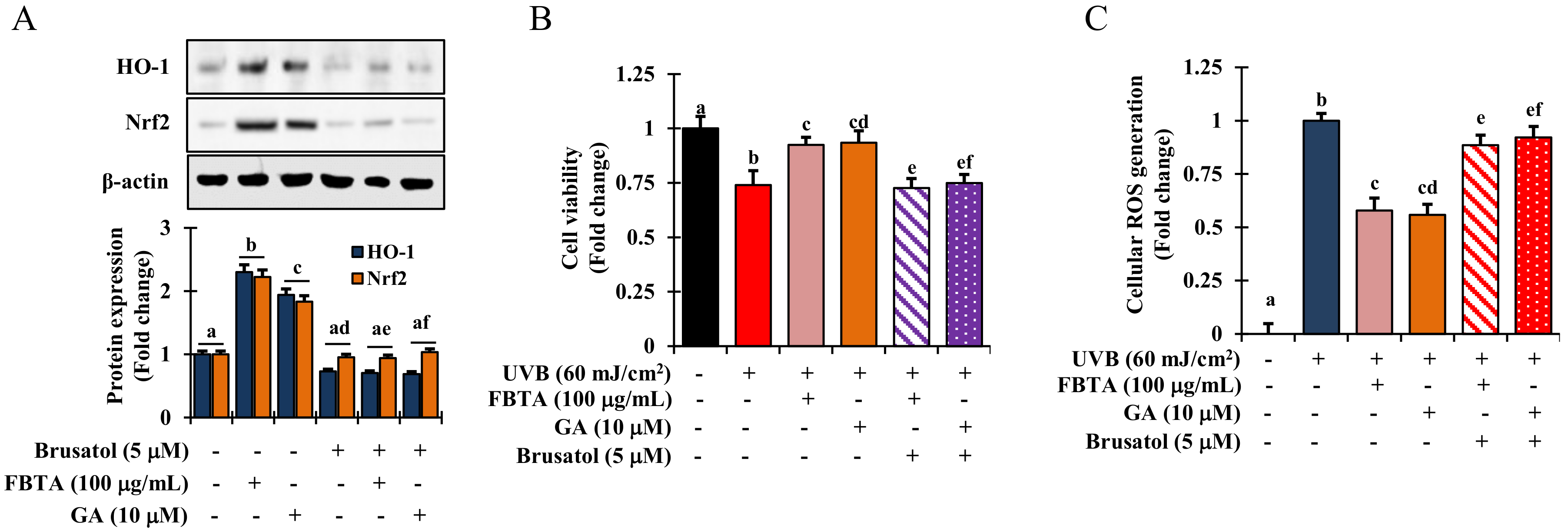
3.5. Acceleration of HO-1 Enzymes via Nrf2 Nuclear Translocation in HaCaT Cells
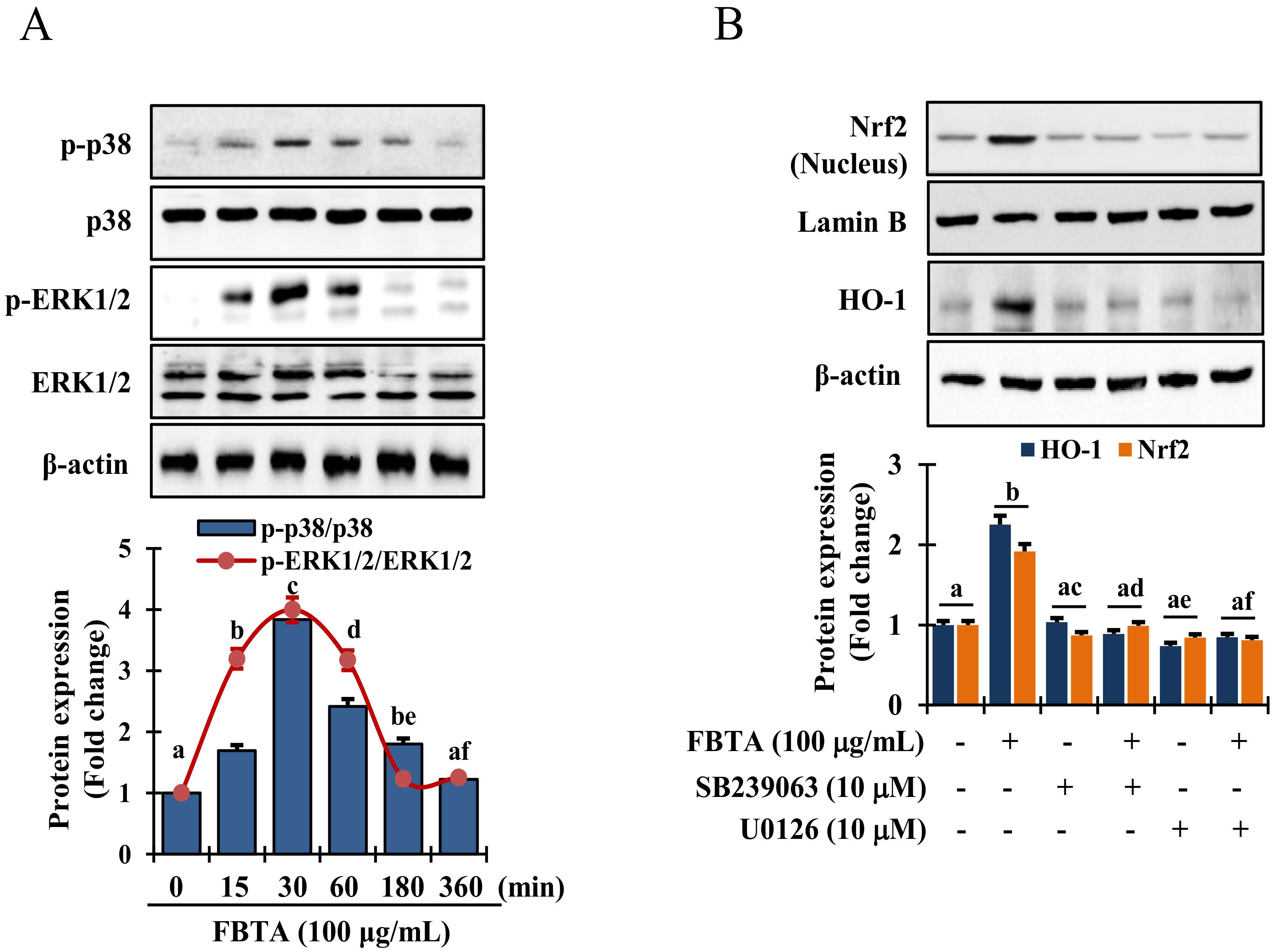
3.6. Effects of FBTA on the MAPK Signaling Pathway
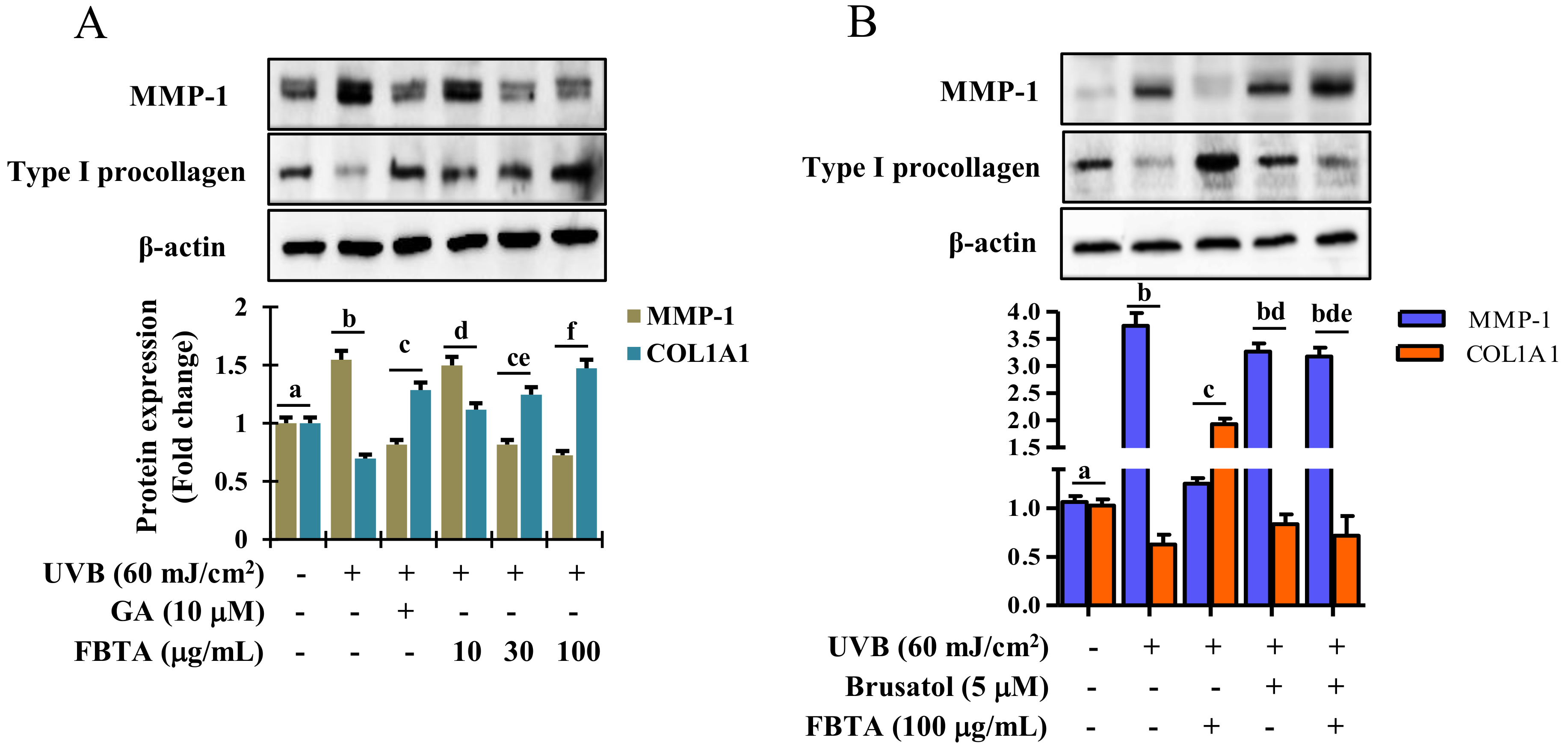
3.7. Effects of FBTA on the Expressions of MMP-1 and Procollagen Type I
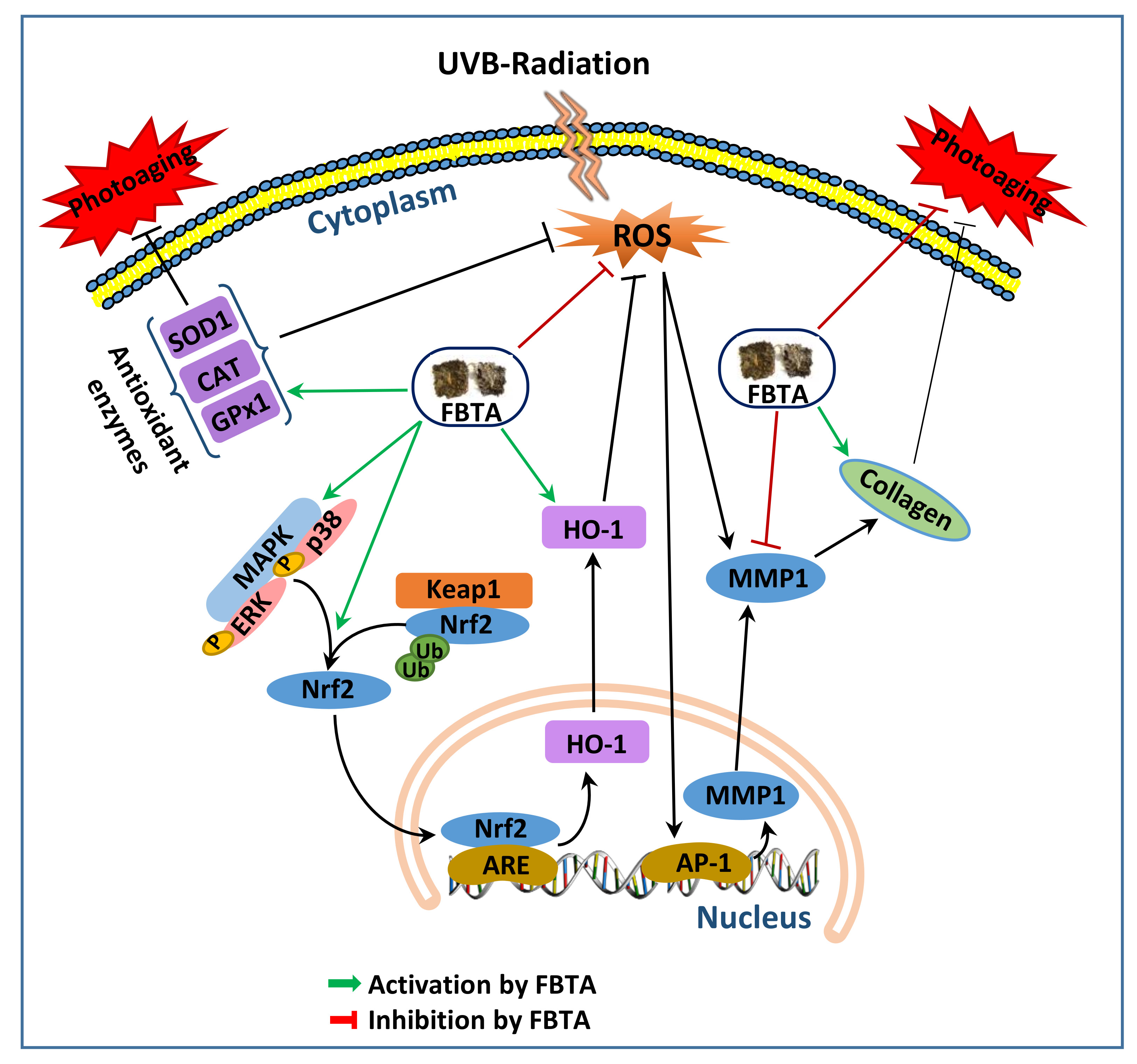
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rittie, L.; Fisher, G.J. Uv-light-induced signal cascades and skin aging. Ageing Res. Rev. 2002, 1, 705–720. [Google Scholar] [CrossRef]
- Hwang, B.M.; Noh, E.M.; Kim, J.S.; Kim, J.M.; Hwang, J.K.; Kim, H.K.; Kang, J.S.; Kim, D.S.; Chae, H.J.; You, Y.O.; et al. Decursin inhibits uvb-induced mmp expression in human dermal fibroblasts via regulation of nuclear factor-kappab. Int. J. Mol. Med. 2013, 31, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Jung, Y.R.; An, H.J.; Kim, D.H.; Jang, E.J.; Choi, Y.J.; Moon, K.M.; Park, M.H.; Park, C.H.; Chung, K.W.; et al. Anti-wrinkle and anti-inflammatory effects of active garlic components and the inhibition of mmps via nf-kappab signaling. PLoS ONE 2013, 8, e73877. [Google Scholar]
- Afaq, F.; Adhami, V.M.; Mukhtar, H. Photochemoprevention of ultraviolet b signaling and photocarcinogenesis. Mutat. Res. 2005, 571, 153–173. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.N.; Lee, Y.H.; Park, J.S.; Jun, W.J.; Kim, H.K.; Hong, B.S.; Shin, D.H.; Cho, H.Y. Protective effects of aucubin isolated from eucommia ulmoides against uvb-induced oxidative stress in human skin fibroblasts. Biol. Pharm. Bull. 2005, 28, 1244–1248. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Park, S.Y.; Hwang, E.; Zhang, M.; Jin, F.; Zhang, B.; Yi, T.H. Salvianolic acid b protects normal human dermal fibroblasts against ultraviolet b irradiation-induced photoaging through mitogen-activated protein kinase and activator protein-1 pathways. Photochem. Photobiol. 2015, 91, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Hu, J.Y.; Wang, S.Q. The role of antioxidants in photoprotection: A critical review. J. Am. Acad. Dermatol. 2012, 67, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Hseu, Y.C.; Lo, H.W.; Korivi, M.; Tsai, Y.C.; Tang, M.J.; Yang, H.L. Dermato-protective properties of ergothioneine through induction of nrf2/are-mediated antioxidant genes in uva-irradiated human keratinocytes. Free Radic. Biol. Med. 2015, 86, 102–117. [Google Scholar] [CrossRef]
- Marrot, L.; Jones, C.; Perez, P.; Meunier, J.R. The significance of nrf2 pathway in (photo)-oxidative stress response in melanocytes and keratinocytes of the human epidermis. Pigment Cell Melanoma Res. 2008, 21, 79–88. [Google Scholar] [CrossRef]
- Ben-Yehuda Greenwald, M.; Frusic-Zlotkin, M.; Soroka, Y.; Ben-Sasson, S.; Bianco-Peled, H.; Kohen, R. A novel role of topical iodine in skin: Activation of the nrf2 pathway. Free Radic. Biol. Med. 2017, 104, 238–248. [Google Scholar] [CrossRef]
- Sun, Z.; Park, S.Y.; Hwang, E.; Zhang, M.; Seo, S.A.; Lin, P.; Yi, T.H. Thymus vulgaris alleviates uvb irradiation induced skin damage via inhibition of mapk/ap-1 and activation of nrf2-are antioxidant system. J. Cell. Mol. Med. 2017, 21, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, K.; Motohashi, H.; Yamamoto, M. Molecular mechanisms of the keap1–nrf2 pathway in stress response and cancer evolution. Genes Cells 2011, 16, 123–140. [Google Scholar] [CrossRef]
- Bae, J.Y.; Choi, J.S.; Choi, Y.J.; Shin, S.Y.; Kang, S.W.; Han, S.J.; Kang, Y.H. (-)epigallocatechin gallate hampers collagen destruction and collagenase activation in ultraviolet-b-irradiated human dermal fibroblasts: Involvement of mitogen-activated protein kinase. Food Chem. Toxicol. 2008, 46, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of matrix metalloproteinases in photoaging and photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, A.; Liu, P.; Li, Z. Effects of fuzhuan brick-tea water extract on mice infected with e. Coli o157:H7. Nutrients 2015, 7, 5309–5326. [Google Scholar] [CrossRef] [PubMed]
- Mo, H.; Zhu, Y.; Chen, Z. Microbial fermented tea–a potential source of natural food preservatives. Trends Food Sci. Technol. 2008, 19, 124–130. [Google Scholar] [CrossRef]
- Keller, A.C.; Weir, T.L.; Broeckling, C.D.; Ryan, E.P. Antibacterial activity and phytochemical profile of fermented camellia sinensis (fuzhuan tea). Food Res. Int. 2013, 53, 945–949. [Google Scholar] [CrossRef]
- Luo, Z.M.; Ling, T.J.; Li, L.X.; Zhang, Z.Z.; Zhu, H.T.; Zhang, Y.J.; Wan, X.C. A new norisoprenoid and other compounds from fuzhuan brick tea. Molecules 2012, 17, 3539–3546. [Google Scholar] [CrossRef]
- Ling, T.J.; Wan, X.C.; Ling, W.W.; Zhang, Z.Z.; Xia, T.; Li, D.X.; Hou, R.Y. New triterpenoids and other constituents from a special microbial-fermented tea-fuzhuan brick tea. J. Agric. Food Chem. 2010, 58, 4945–4950. [Google Scholar] [CrossRef]
- Xu, X.; Mo, H.; Yan, M.; Zhu, Y. Analysis of characteristic aroma of fungal fermented fuzhuan brick-tea by gas chromatography/mass spectrophotometry. J. Sci. Food Agric. 2007, 87, 1502–1504. [Google Scholar] [CrossRef]
- Li, Q.; Liu, Z.; Huang, J.; Luo, G.; Liang, Q.; Wang, D.; Ye, X.; Wu, C.; Wang, L.; Hu, J. Anti-obesity and hypolipidemic effects of fuzhuan brick tea water extract in high-fat diet-induced obese rats. J. Sci. Food Agric. 2013, 93, 1310–1316. [Google Scholar] [CrossRef] [PubMed]
- Jie, G.; Lin, Z.; Zhang, L.; Lv, H.; He, P.; Zhao, B. Free radical scavenging effect of pu-erh tea extracts and their protective effect on oxidative damage in human fibroblast cells. J. Agric. Food Chem. 2006, 54, 8058–8064. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-C.; Yen, G.-C.; Wang, B.-S.; Chiu, C.-K.; Yen, W.-J.; Chang, L.-W.; Duh, P.-D. Antimutagenic and antimicrobial activities of pu-erh tea. LWT Food Sci. Technol. 2007, 40, 506–512. [Google Scholar] [CrossRef]
- Alam, M.B.; Ju, M.K.; Lee, S.H. DNA protecting activities of nymphaea nouchali (burm. F) flower extract attenuate t-bhp-induced oxidative stress cell death through nrf2-mediated induction of heme oxygenase-1 expression by activating map-kinases. Int. J. Mol. Sci. 2017, 18, 2069. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.B.; Kwon, K.R.; Lee, S.H.; Lee, S.H. Lannea coromandelica (houtt.) merr. Induces heme oxygenase 1 (ho-1) expression and reduces oxidative stress via the p38/c-jun n-terminal kinase-nuclear factor erythroid 2-related factor 2 (p38/jnk-nrf2)-mediated antioxidant pathway. Int. J. Mol. Sci. 2017, 18, 266. [Google Scholar] [CrossRef] [PubMed]
- Buechner, N.; Schroeder, P.; Jakob, S.; Kunze, K.; Maresch, T.; Calles, C.; Krutmann, J.; Haendeler, J. Changes of mmp-1 and collagen type ialpha1 by uva, uvb and ira are differentially regulated by trx-1. Exp. Gerontol. 2008, 43, 633–637. [Google Scholar] [CrossRef] [PubMed]
- Batchelder, R.J.; Calder, R.J.; Thomas, C.P.; Heard, C.M. In vitro transdermal delivery of the major catechins and caffeine from extract of camellia sinensis. Int. J. Pharm. 2004, 283, 45–51. [Google Scholar] [CrossRef]
- Chaiprasongsuk, A.; Lohakul, J.; Soontrapa, K.; Sampattavanich, S.; Akarasereenont, P.; Panich, U. Activation of nrf2 reduces uva-mediated mmp-1 upregulation via mapk/ap-1 signaling cascades: The photoprotective effects of sulforaphane and hispidulin. J. Pharmacol. Exp. Ther. 2017, 360, 388–398. [Google Scholar] [CrossRef]
- Hseu, Y.C.; Korivi, M.; Lin, F.Y.; Li, M.L.; Lin, R.W.; Wu, J.J.; Yang, H.L. Trans-cinnamic acid attenuates uva-induced photoaging through inhibition of ap-1 activation and induction of nrf2-mediated antioxidant genes in human skin fibroblasts. J. Dermatol. Sci. 2018, 90, 123–134. [Google Scholar] [CrossRef]
- Song, J.L.; Gao, Y. Effects of methanolic extract form fuzhuan brick-tea on hydrogen peroxide-induced oxidative stress in human intestinal epithelial adenocarcinoma caco-2 cells. Mol. Med. Rep. 2014, 9, 1061–1067. [Google Scholar] [CrossRef]
- Seo, S.A.; Park, B.; Hwang, E.; Park, S.Y.; Yi, T.H. Borago officinalis l. Attenuates uvb-induced skin photodamage via regulation of ap-1 and nrf2/are pathway in normal human dermal fibroblasts and promotion of collagen synthesis in hairless mice. Exp. Gerontol. 2018, 107, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.P.; Zhang, Y.; Shi, J.; Lin, Z. Phytochemical profiles and antioxidant activities of chinese dark teas obtained by different processing technologies. Food Res. Int. 2017, 100, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, D.; Chen, W.; Tan, X.; Wang, P. Impact of fermentation degree on the antioxidant activity of pu-erh tea in vitro. J. Food Biochem. 2012, 36, 262–267. [Google Scholar] [CrossRef]
- Shin, D.; Lee, S.; Huang, Y.H.; Lim, H.W.; Lee, Y.; Jang, K.; Cho, Y.; Park, S.J.; Kim, D.D.; Lim, C.J. Protective properties of geniposide against uv-b-induced photooxidative stress in human dermal fibroblasts. Pharm. Biol. 2018, 56, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wang, M.; Xie, M.; Wan, P.; Chen, D.; Hu, B.; Ye, H.; Zeng, X.; Liu, Z. Evaluation of chemical property, cytotoxicity and antioxidant activity in vitro and in vivo of polysaccharides from fuzhuan brick teas. Int. J. Biol. Macromol. 2018, 116, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Mishra, M.; Fomusi Ndisang, J. A critical and comprehensive insight on heme oxygenase and related products including carbon monoxide, bilirubin, biliverdin and ferritin in type-1 and type-2 diabetes. Curr. Pharm. Des. 2014, 20, 1370–1391. [Google Scholar] [CrossRef] [PubMed]
- Pullikotil, P.; Chen, H.; Muniyappa, R.; Greenberg, C.C.; Yang, S.; Reiter, C.E.; Lee, J.-W.; Chung, J.H.; Quon, M.J. Epigallocatechin gallate induces expression of heme oxygenase-1 in endothelial cells via p38 mapk and nrf-2 that suppresses proinflammatory actions of tnf-α. J. Nutr. Biochem. 2012, 23, 1134–1145. [Google Scholar] [CrossRef]
- Martín, M.Á.; Serrano, A.B.G.; Ramos, S.; Pulido, M.I.; Bravo, L.; Goya, L. Cocoa flavonoids up-regulate antioxidant enzyme activity via the erk1/2 pathway to protect against oxidative stress-induced apoptosis in hepg2 cells. J. Nutr. Biochem. 2010, 21, 196–205. [Google Scholar] [CrossRef]
- Hirota, A.; Kawachi, Y.; Yamamoto, M.; Koga, T.; Hamada, K.; Otsuka, F. Acceleration of uvb-induced photoageing in nrf2 gene-deficient mice. Exp. Dermatol. 2011, 20, 664–668. [Google Scholar] [CrossRef]
- Tao, S.; Park, S.L.; Rojo de la Vega, M.; Zhang, D.D.; Wondrak, G.T. Systemic administration of the apocarotenoid bixin protects skin against solar uv-induced damage through activation of nrf2. Free Radic. Biol. Med. 2015, 89, 690–700. [Google Scholar] [CrossRef]
- Tao, S.; Justiniano, R.; Zhang, D.D.; Wondrak, G.T. The nrf2-inducers tanshinone i and dihydrotanshinone protect human skin cells and reconstructed human skin against solar simulated uv. Redox Biol. 2013, 1, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Lv, L.; Lu, Q.; Li, Y.; Zhang, F.; Lin, M.; Gao, D.; Liu, K.; Tian, X.; Yao, J. Gallic acid attenuates dimethylnitrosamine-induced acute liver injury in mice through nrf2-mediated induction of heme oxygenase-1 and glutathione-s-transferase α 3. Med. Chem. 2014, 4, 663–669. [Google Scholar] [CrossRef]
- Chen, H.-H.; Wang, T.-C.; Lee, Y.-C.; Shen, P.-T.; Chang, J.-Y.; Yeh, T.-K.; Huang, C.-H.; Chang, H.-H.; Cheng, S.-Y.; Lin, C.-Y. Novel nrf2/are activator, trans-coniferylaldehyde, induces a ho-1-mediated defense mechanism through a dual p38α/mapkapk-2 and pk-n3 signaling pathway. Chem. Res. Toxicol. 2015, 28, 1681–1692. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.T.; Yen, G.C. Involvement of p38 mapk and nrf2 in phenolic acid-induced p-form phenol sulfotransferase expression in human hepatoma hepg2 cells. Carcinogenesis 2006, 27, 1008–1017. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Park, S.Y.; Hwang, E.; Park, B.; Seo, S.A.; Cho, J.G.; Zhang, M.; Yi, T.H. Dietary foeniculum vulgare mill extract attenuated uvb irradiation-induced skin photoaging by activating of nrf2 and inhibiting mapk pathways. Phytomedicine 2016, 23, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Park, Y.G.; Lee, H.J.; Lim, S.J.; Nho, C.W. Youngiasides a and c isolated from youngia denticulatum inhibit uvb-induced mmp expression and promote type i procollagen production via repression of mapk/ap-1/nf-kappab and activation of ampk/nrf2 in hacat cells and human dermal fibroblasts. J. Agric. Food Chem. 2015, 63, 5428–5438. [Google Scholar] [CrossRef]
- Hwang, E.; Park, S.Y.; Lee, H.J.; Lee, T.Y.; Sun, Z.W.; Yi, T.H. Gallic acid regulates skin photoaging in uvb-exposed fibroblast and hairless mice. Phytother. Res. 2014, 28, 1778–1788. [Google Scholar] [CrossRef]
- Choi, H.S.; Park, E.D.; Park, Y.; Han, S.H.; Hong, K.B.; Suh, H.J. Topical application of spent coffee ground extracts protects skin from ultraviolet b-induced photoaging in hairless mice. Photochem. Photobiol. Sci. 2016, 15, 779–790. [Google Scholar] [CrossRef]
- Clifford, M.N.; van der Hooft, J.J.; Crozier, A. Human studies on the absorption, distribution, metabolism, and excretion of tea polyphenols. Am. J. Clin. Nutr. 2013, 98, 1619s–1630s. [Google Scholar] [CrossRef]
- Hodgson, J.M.; Morton, L.W.; Puddey, I.B.; Beilin, L.J.; Croft, K.D. Gallic acid metabolites are markers of black tea intake in humans. J. Agric. Food Chem. 2000, 48, 2276–2280. [Google Scholar] [CrossRef]
- Shahrzad, S.; Aoyagi, K.; Winter, A.; Koyama, A.; Bitsch, I. Pharmacokinetics of gallic acid and its relative bioavailability from tea in healthy humans. J. Nutr. 2001, 131, 1207–1210. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, P.; Alam, M.B.; Lee, S.-H. Protection of UVB-Induced Photoaging by Fuzhuan-Brick Tea Aqueous Extract via MAPKs/Nrf2-Mediated Down-Regulation of MMP-1. Nutrients 2019, 11, 60. https://doi.org/10.3390/nu11010060
Zhao P, Alam MB, Lee S-H. Protection of UVB-Induced Photoaging by Fuzhuan-Brick Tea Aqueous Extract via MAPKs/Nrf2-Mediated Down-Regulation of MMP-1. Nutrients. 2019; 11(1):60. https://doi.org/10.3390/nu11010060
Chicago/Turabian StyleZhao, Peijun, Md Badrul Alam, and Sang-Han Lee. 2019. "Protection of UVB-Induced Photoaging by Fuzhuan-Brick Tea Aqueous Extract via MAPKs/Nrf2-Mediated Down-Regulation of MMP-1" Nutrients 11, no. 1: 60. https://doi.org/10.3390/nu11010060
APA StyleZhao, P., Alam, M. B., & Lee, S. -H. (2019). Protection of UVB-Induced Photoaging by Fuzhuan-Brick Tea Aqueous Extract via MAPKs/Nrf2-Mediated Down-Regulation of MMP-1. Nutrients, 11(1), 60. https://doi.org/10.3390/nu11010060