Hypnotic Effects of Lactobacillus fermentum PS150TM on Pentobarbital-Induced Sleep in Mice

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain, Media, and Growth Conditions
2.2. Animals
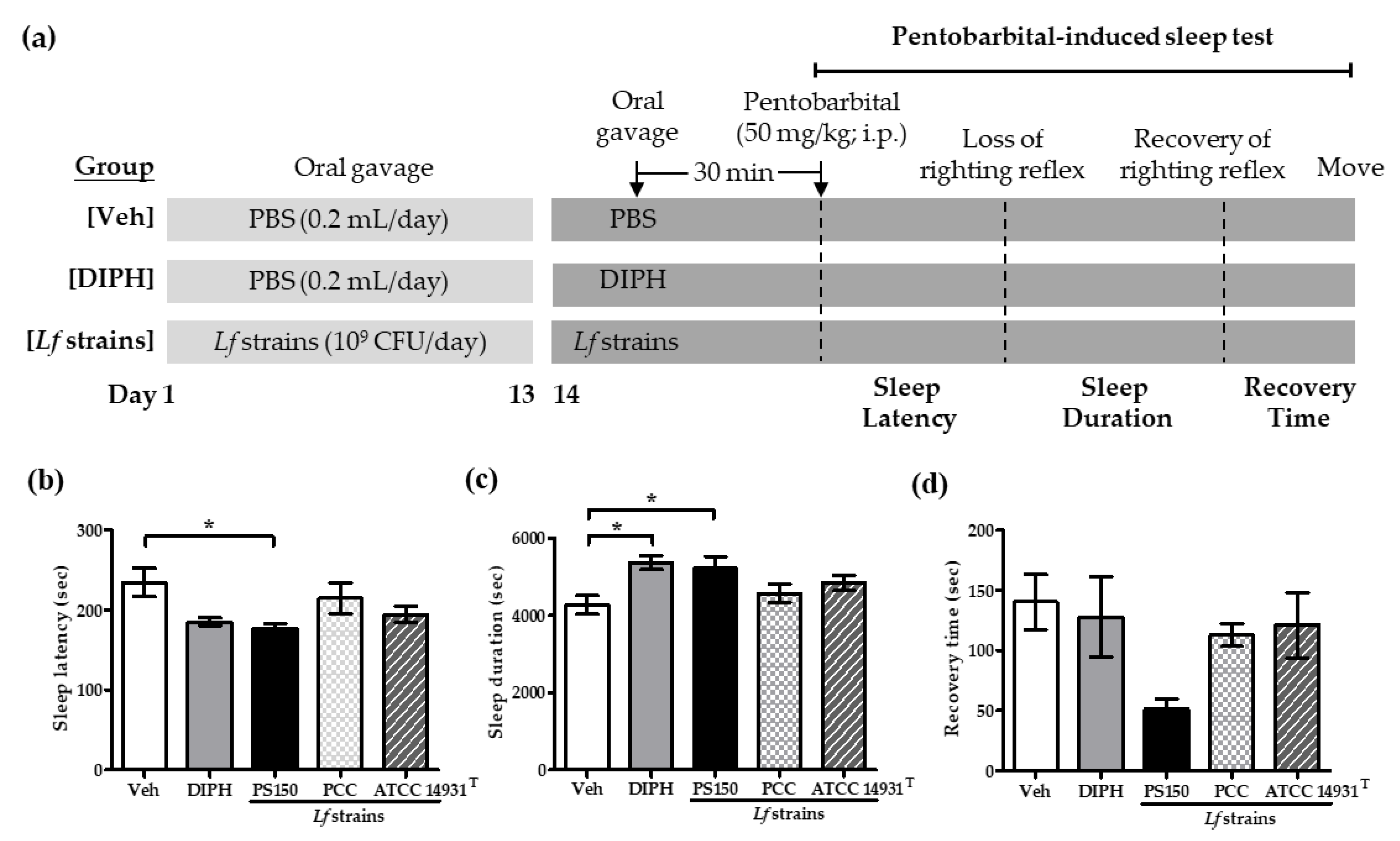
2.3. Pentobarbital-Induced Sleep Test
2.4. Open Field Test
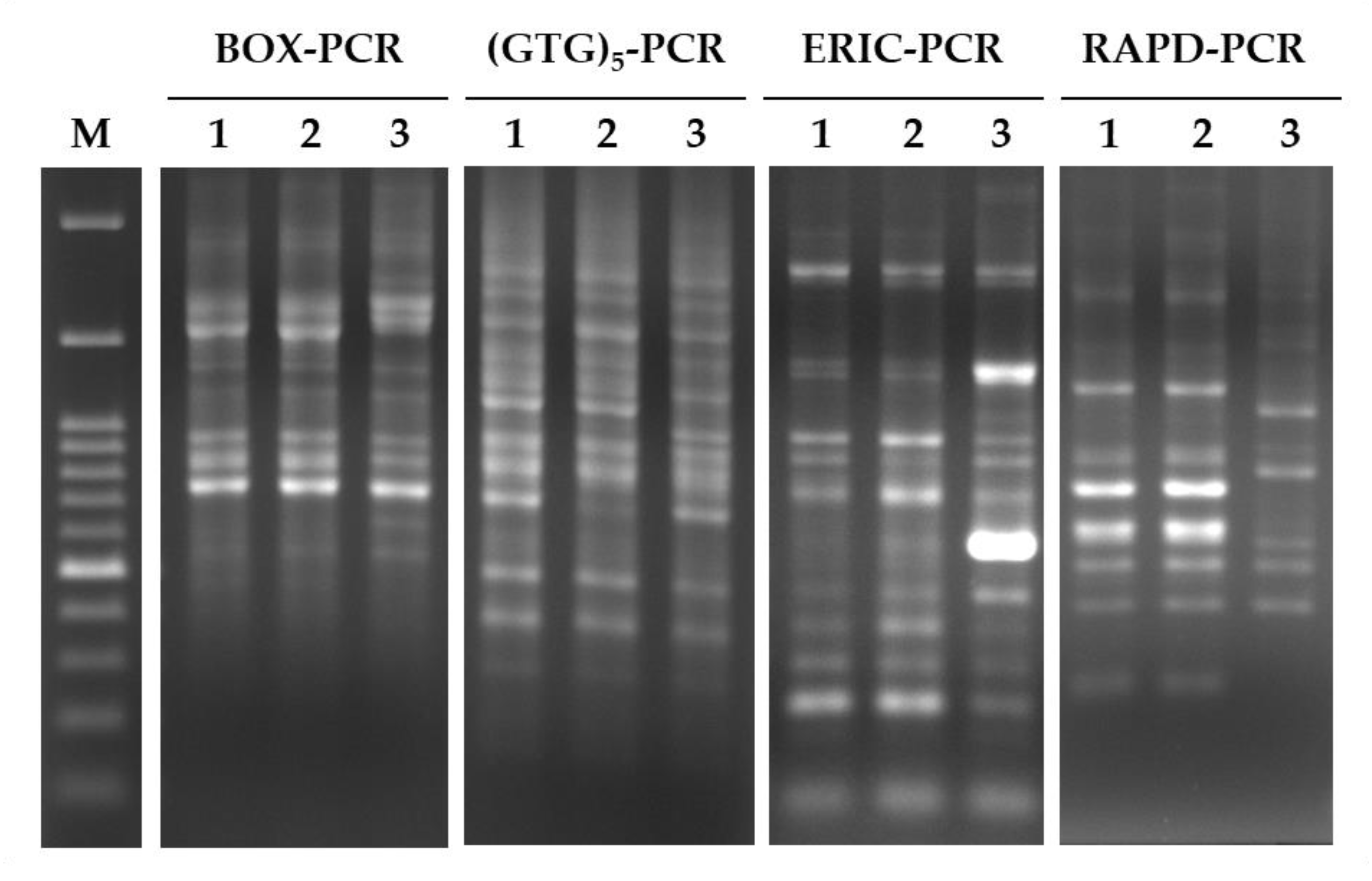
2.5. Genomic DNA Extraction and PCR Analysis of L. fermentum Strains
2.6. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
2.7. Statistical Analysis
3. Results
3.1. Strain-Specific Effects of PS150TM on Pentobarbital-Induced Sleep in Mice
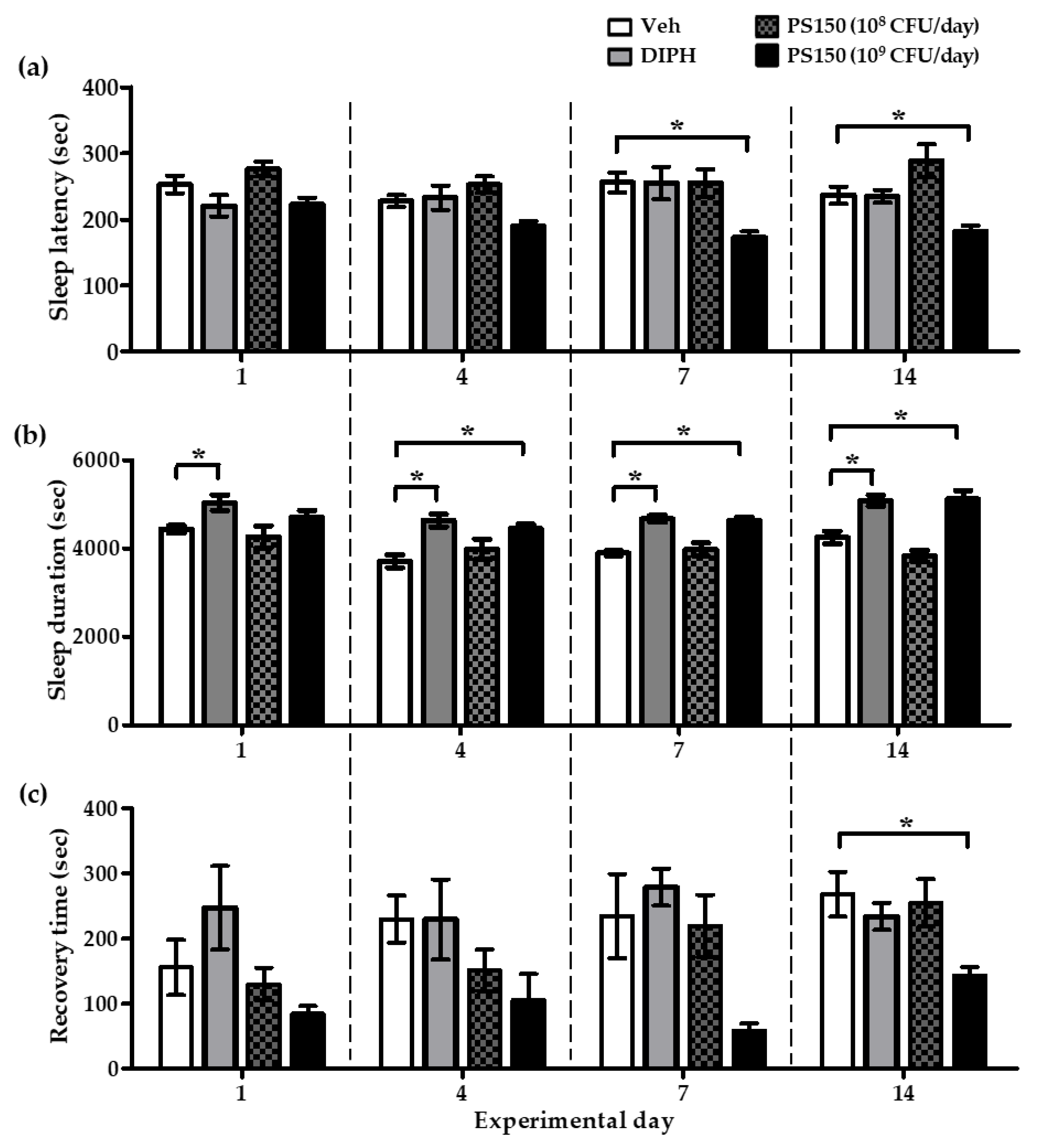
3.2. PS150TM Improved Pentobarbital-Induced Sleep in Mice in a Time- and Dose-Dependent Manner
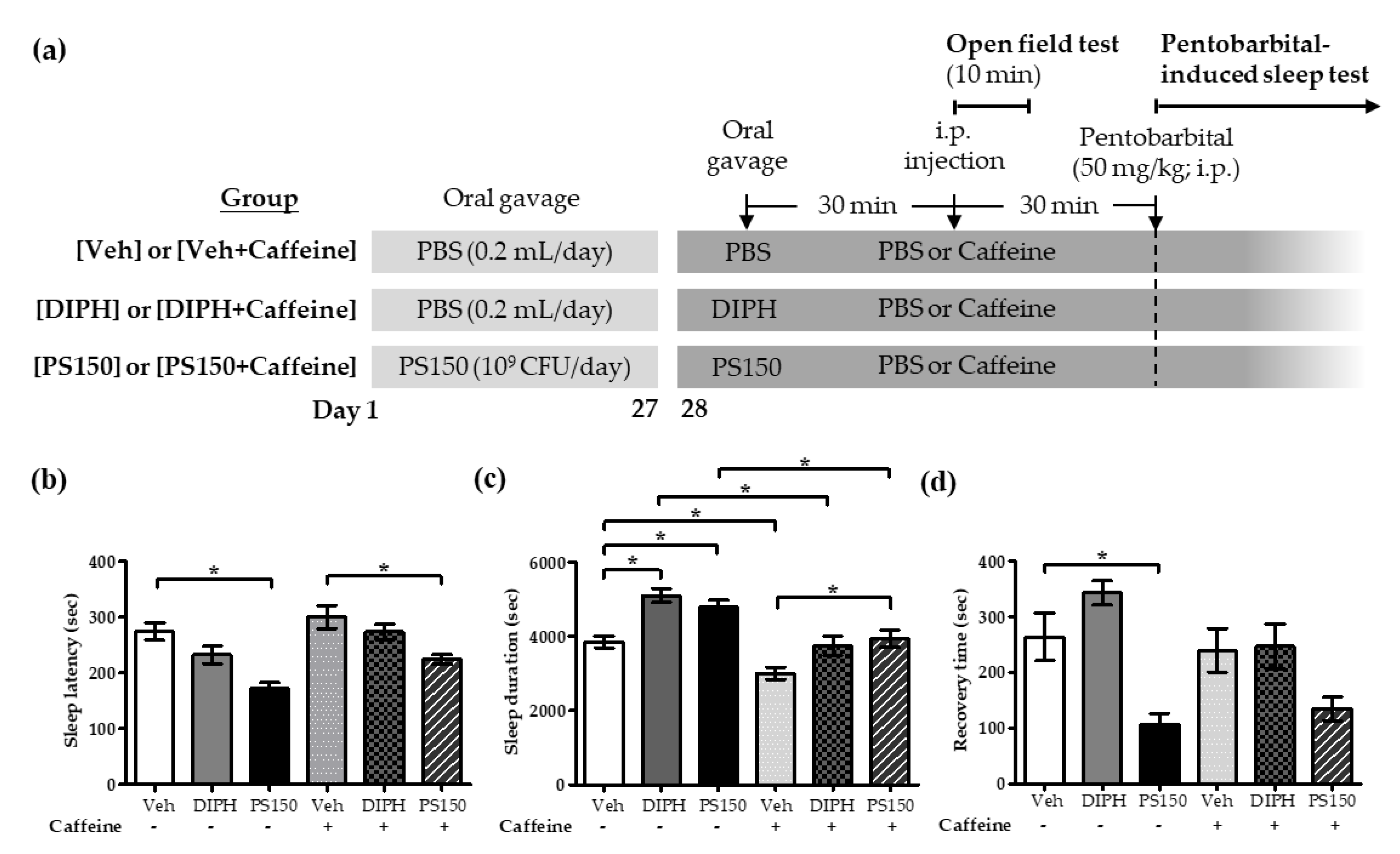
3.3. Effects of PS150TM on Caffeine-Induced Sleep Disruption in Mice
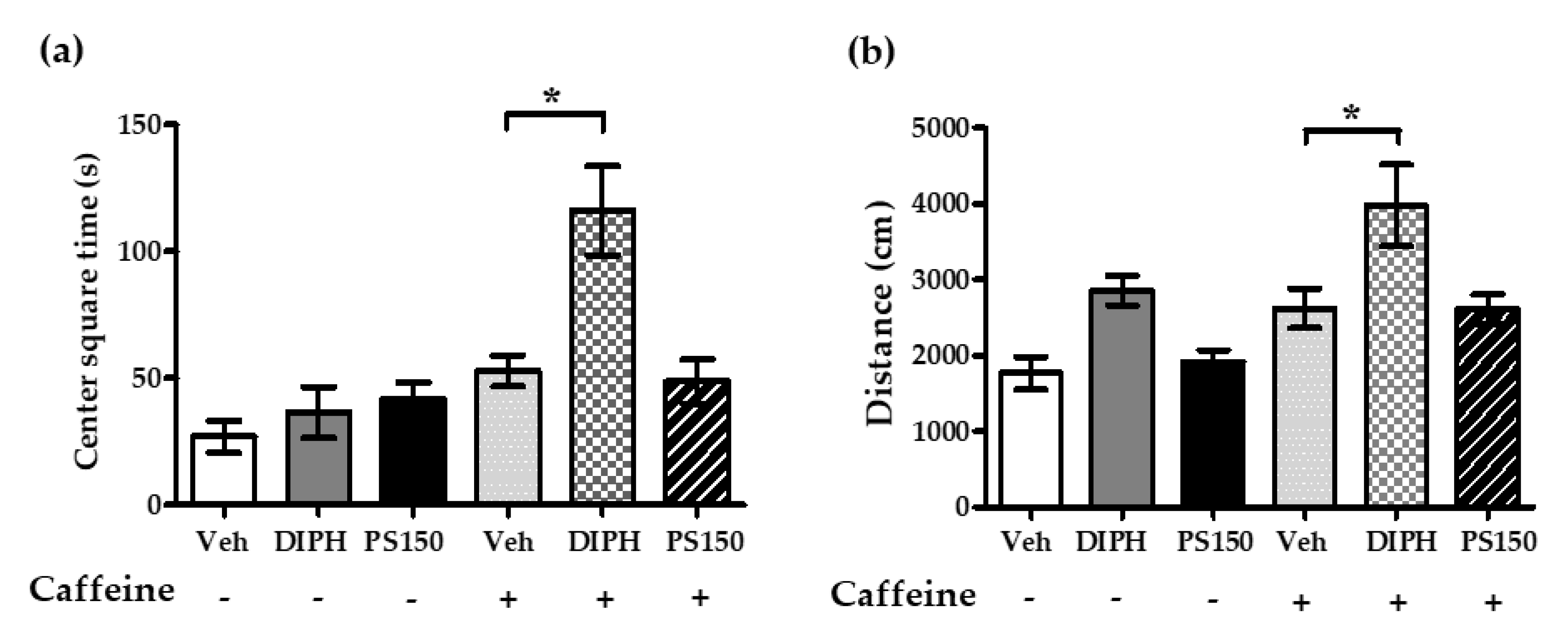
3.4. PS150TM Did Not Alter the Spontaneous Locomotor Activity of Mice in the Open Field Test
3.5. Effects of PS150TM on the Expression of Genes Related to Adenosine Signaling in the Basal Forebrain and Hypothalamus
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Morin, C.M.; Benca, R. Chronic insomnia. Lancet 2012, 379, 1129–1141. [Google Scholar] [CrossRef]
- Krystal, A.D.; Benca, R.M.; Kilduff, T.S. Understanding the sleep-wake cycle: Sleep, insomnia, and the orexin system. J. Clin. Psychiatry 2013, 74, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.M.; Pietrone, R.; Cashmere, J.D.; Begley, A.; Miewald, J.M.; Germain, A.; Buysse, D.J. EEG power during waking and NREM sleep in primary insomnia. J. Clin. Sleep Med. 2013, 9, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Parekh, P.J.; Oldfield, E.C.T.; Johnson, D.A. The Effects of Sleep on the Commensal Microbiota: Eyes Wide Open? J. Clin. Gastroenterol. 2018, 52, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Buysse, D.J. Insomnia. JAMA 2013, 309, 706–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orr, W.C.; Chen, C.L. Sleep and the gastrointestinal tract. Neurol. Clin. 2005, 23, 1007–1024. [Google Scholar] [CrossRef] [PubMed]
- Parekh, P.J.; Oldfield Iv, E.C.; Challapallisri, V.; Ware, J.C.; Johnson, D.A. Sleep disorders and inflammatory disease activity: Chicken or the egg? Am. J. Gastroenterol. 2015, 110, 484–488. [Google Scholar] [CrossRef]
- Voigt, R.M.; Forsyth, C.B.; Green, S.J.; Mutlu, E.; Engen, P.; Vitaterna, M.H.; Turek, F.W.; Keshavarzian, A. Circadian Disorganization Alters Intestinal Microbiota. PLoS ONE 2014, 21, e97500. [Google Scholar] [CrossRef]
- FAO; WHO. Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic: Report of a Joint FAO/WHO Expert Consultation on Acid Bacteria Expert Consultation on Evaluation of Health and Nutritional Properties of Probitics in Food Including Powder Milk with Live Lactic acid Bacteria; FAO: Rome, Italy; WHO: Geneva, Switzerland, 2001. [Google Scholar]
- Weizman, Z. The role of probiotics and prebiotics in the prevention of infections in child day-care centres. Benef. Microbes 2015, 6, 181–183. [Google Scholar] [CrossRef]
- Takeda, K.; Suzuki, T.; Shimada, S.I.; Shida, K.; Nanno, M.; Okumura, K. Interleukin-12 is involved in the enhancement of human natural killer cell activity by Lactobacillus casei Shirota. Clin. Exp. Immunol. 2006, 146, 109–115. [Google Scholar] [CrossRef]
- Baldassarre, M.E.; Palladino, V.; Amoruso, A.; Pindinelli, S.; Mastromarino, P.; Fanelli, M.; Di Mauro, A.; Laforgia, N. Rationale of Probiotic Supplementation during Pregnancy and Neonatal Period. Nutrients 2018, 10, 1693. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Cryan, J.F. The Microbiome-Gut-Brain Axis in Health and Disease. Gastroenterol. Clin. North Am. 2017, 46, 77–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyte, M. Probiotics function mechanistically as delivery vehicles for neuroactive compounds: Microbial endocrinology in the design and use of probiotics. Bioessays 2011, 33, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cao, S.; Zhang, X. Modulation of Gut Microbiota-Brain Axis by Probiotics, Prebiotics, and Diet. J. Agric. Food Chem. 2015, 63, 7885–7895. [Google Scholar] [CrossRef] [PubMed]
- Abi, S.A.; Marques, A.M. Louis Gardan and Régine Samson. BOX-PCR-based identification of bacterial species belonging to Pseudomonas syringae—P. viridiflava group. Genet. Mol. Biol. 2008, 31, 106–115. [Google Scholar]
- Cox, A.J.; Pyne, D.B.; Saunders, P.U.; Fricker, P.A. Oral administration of the probiotic Lactobacillus fermentum VRI-003 and mucosal immunity in endurance athletes. Br. J. Sports Med. 2010, 44, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Farías, M.E.; de Saad AM, S.; de Ruiz Holgado AA, P.; Oliver, G. Purification and properties of L-serine dehydratase from Lactobacillus fermentum ATCC 14931. Curr. Microbiol. 1991, 22, 205–211. [Google Scholar] [CrossRef]
- Yang, H.; Yoon, M.; Um, M.Y.; Lee, J.; Jung, J.; Lee, C.; Kim, Y.T.; Kwon, S.; Kim, B.; Cho, S. Sleep-Promoting Effects and Possible Mechanisms of Action Associated with a Standardized Rice Bran Supplement. Nutrients 2017, 9, 512. [Google Scholar] [CrossRef]
- Hong, K.B.; Park, Y.; Suh, H.J. Two combined amino acids promote sleep activity in caffeine-induced sleepless model systems. Nutr. Res. Pract. 2018, 12, 208–214. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, L.; Wang, G.; He, Z.; Zhao, Y.; Xu, Y.; Gao, Y.; Zhang, L. Sedative and hypnotic effects of supercritical carbon dioxide fluid extraction from Schisandra chinensis in mice. J. Food Drug Anal. 2016, 24, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Ko, Y.H.; Shim, K.Y.; Lee, S.Y.; Jang, C.G. Evodiamine Reduces Caffeine-Induced Sleep Disturbances and Excitation in Mice. Biomol. Ther. 2018, 26, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.H.; Chuang, H.L.; Huang, Y.T.; Wu, C.C.; Chou, G.T.; Wang, S.; Tsai, Y.C. Alteration of behavior and monoamine levels attributable to Lactobacillus plantarum PS128 in germ-free mice. Behav. Brain Res. 2016, 298, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Liu, W.H.; Wu, C.C.; Juan, Y.C.; Wu, Y.C.; Tsai, H.P.; Wang, S.; Tsai, Y.C. Psychotropic effects of Lactobacillus plantarum PS128 in early life-stressed and naive adult mice. Brain Res. 2016, 1631, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chao, S.H.; Tomii, Y.; Watanabe, K.; Tsai, Y.C. Diversity of lactic acid bacteria in fermented brines used to make stinky tofu. Int. J. Food Microbiol. 2008, 123, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Hulton, C.S.; Higgins, C.F.; Sharp, P.M. ERIC sequences: A novel family of repetitive elements in the genomes of Escherichia coli, Salmonella typhimurium and other enterobacteria. Mol. Microbiol. 1991, 5, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, N.C.; Ficoseco, C.A.; Mansilla, F.I.; Melián, C.; Hébert, E.M.; Vignolo, G.M.; Nader-Macías, M.E.F. Identification, characterization and selection of autochthonous lactic acid bacteria as probiotic for feedlot cattle. Livest. Sci. 2018, 212, 99–110. [Google Scholar] [CrossRef]
- Tannock, G.W.; Wilson, C.M.; Loach, D.; Cook, G.M.; Eason, J.; O’Toole, P.W.; Holtrop, G.; Lawley, B. Resource partitioning in relation to cohabitation of Lactobacillus species in the mouse forestomach. ISME J. 2012, 6, 927–938. [Google Scholar] [CrossRef]
- Jhaveri, K.A.; Reichensperger, J.; Toth, L.A.; Sekino, Y.; Ramkumar, V. Reduced basal and lipopolysaccharide-stimulated adenosine A1 receptor expression in the brain of nuclear factor-kappaB p50-/- mice. Neuroscience 2007, 146, 415–426. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Takata, Y.; Li, R.; Zhang, Z.; Zhang, M.Q.; Urade, Y.; Qu, W.M.; Huang, Z.L. Doxepin and diphenhydramine increased non-rapid eye movement sleep through blockade of histamine H1 receptors. Pharmacol. Biochem. Behav. 2015, 129, 56–64. [Google Scholar] [CrossRef]
- Park, E.; Younes, M.; Liu, H.; Liu, X.; Horner, R.L. Systemic vs. central administration of common hypnotics reveals opposing effects on genioglossus muscle activity in rats. Sleep 2008, 31, 355–365. [Google Scholar] [CrossRef]
- Basheer, R.; Strecker, R.E.; Thakkar, M.M.; McCarley, R.W. Adenosine and sleep-wake regulation. Prog. Neurobiol. 2004, 73, 379–396. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.W.; Gao, X.B. Adenosine inhibits activity of hypocretin/orexin neurons by the A1 receptor in the lateral hypothalamus: A possible sleep-promoting effect. J. Neurophysiol. 2007, 97, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Szymusiak, R.; Gvilia, I.; McGinty, D. Hypothalamic control of sleep. Sleep Med. 2007, 8, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Parkar, S.G.; Kalsbeek, A.; Cheeseman, J.F. Potential Role for the Gut Microbiota in Modulating Host Circadian Rhythms and Metabolic Health. Microorganisms 2019, 7, 41. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hao, Y.; Fan, F.; Zhang, B. The Role of Microbiome in Insomnia, Circadian Disturbance and Depression. Front. Psychiatry 2018, 9, 669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Han, Y.; Du, J.; Liu, R.; Jin, K.; Yi, W. Microbiota-gut-brain axis and the central nervous system. Oncotarget 2017, 8, 53829–53838. [Google Scholar] [CrossRef]
- Sheth, R.U.; Cabral, V.; Chen, S.P.; Wang, H.H. Manipulating Bacterial Communities by in situ Microbiome Engineering. Trends Genet. 2016, 32, 189–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.T.; Walsh, R.F.L.; Sheehan, A.E. Prebiotics and probiotics for depression and anxiety: A systematic review and meta-analysis of controlled clinical trials. Neurosci. Biobehav. Rev. 2019, 102, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, K.; Itoh, N.; Yamamoto, S.; Higo-Yamamoto, S.; Nakakita, Y.; Kaneda, H.; Shigyo, T.; Oishi, K. Dietary heat-killed Lactobacillus brevis SBC8803 promotes voluntary wheel-running and affects sleep rhythms in mice. Life Sci. 2014, 111, 47–52. [Google Scholar] [CrossRef]
- Nakamura, Y.; Midorikawa, T.; Monoi, N.; Kimura, E.; Murata-Matsuno, A.; Sano, T.; Oka, K.; Sugafuji, T.; Uchiyama, A.; Murakoshi, M.; et al. Oral administration of Japanese sake yeast (Saccharomyces cerevisiae sake) promotes non-rapid eye movement sleep in mice via adenosine A2A receptors. J. Sleep Res. 2016, 25, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Takada, M.; Nishida, K.; Gondo, Y.; Kikuchi-Hayakawa, H.; Ishikawa, H.; Suda, K.; Kawai, M.; Hoshi, R.; Kuwano, Y.; Miyazaki, K.; et al. Beneficial effects of Lactobacillus casei strain Shirota on academic stress-induced sleep disturbance in healthy adults: A double-blind, randomised, placebo-controlled trial. Benef. Microbes 2017, 8, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Nishida, K.; Sawada, D.; Kawai, T.; Kuwano, Y.; Fujiwara, S.; Rokutan, K. Para-psychobiotic Lactobacillus gasseri CP2305 ameliorates stress-related symptoms and sleep quality. J. Appl. Microbiol. 2017, 123, 1561–1570. [Google Scholar] [CrossRef] [PubMed]
- Rakhshandeh, H.; Sadeghnia, H.R.; Ghorbani, A. Sleep-prolonging effect of Coriandrum sativum hydro-alcoholic extract in mice. Nat. Prod. Res. 2012, 26, 2095–2098. [Google Scholar] [CrossRef] [PubMed]
- Rakhshandah, H.; Shakeri, M.T.; Ghasemzadeh, M.R. Comparative Hypnotic Effect of Rosa damascena Fractions and Diazepam in Mice. Iran. J. Pharm. Res. 2007, 6, 193–197. [Google Scholar]
- Amin, J.; Weiss, D.S. GABAA receptor needs two homologous domains of the beta-subunit for activation by GABA but not by pentobarbital. Nature 1993, 366, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Mou, N.; Duan, Z.; Ma, P.; Fu, R.; Fan, D. Study on the hypnotic effect of rare protopanaxadiol-type and protopanaxatriol-type ginsenosides. RSC Adv. 2019, 9, 20483–20491. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.M.; Shimizu, M.; Lee, C.J.; Han, D.S.; Jung, C.K.; Jo, J.H.; Kim, Y.M. Hypnotic effects and binding studies for GABA(A) and 5-HT(2C) receptors of traditional medicinal plants used in Asia for insomnia. J. Ethnopharmacol. 2010, 132, 225–232. [Google Scholar] [CrossRef]
- Kuribara, H.; Asahi, T.; Tadokoro, S. Ethanol enhances, but diazepam and pentobarbital reduce the ambulation-increasing effect of caffeine in mice. Arukoru Kenkyu Yakubutsu Izon 1992, 27, 528–539. [Google Scholar]
- Forrest, W.H., Jr.; Bellville, J.W.; Brown, B.W., Jr. The interaction of caffeine with pentobarbital as a nighttime hypnotic. Anesthesiology 1972, 36, 37–41. [Google Scholar] [CrossRef]
- Suez, J.; Zmora, N.; Segal, E.; Elinav, E. The pros, cons, and many unknowns of probiotics. Nat. Med. 2019, 25, 716–729. [Google Scholar] [CrossRef] [PubMed]
- Luppi, P.H.; Fort, P. Sleep-wake physiology. Handb. Clin. Neurol. 2019, 160, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Yunes, R.A.; Poluektova, E.U.; Dyachkova, M.S.; Klimina, K.M.; Kovtun, A.S.; Averina, O.V.; Orlova, V.S.; Danilenko, V.N. GABA production and structure of gadB/gadC genes in Lactobacillus and Bifidobacterium strains from human microbiota. Anaerobe 2016, 42, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Porkka-Heiskanen, T.; Alanko, L.; Kalinchuk, A.; Stenberg, D. Adenosine and sleep. Sleep Med. Rev. 2002, 6, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Oishi, Y.; Huang, Z.L.; Fredholm, B.B.; Urade, Y.; Hayaishi, O. Adenosine in the tuberomammillary nucleus inhibits the histaminergic system via A1 receptors and promotes non-rapid eye movement sleep. Proc. Natl. Acad. Sci. USA 2008, 105, 19992–19997. [Google Scholar] [CrossRef] [PubMed]
- Cun, Y.; Tang, L.; Yan, J.; He, C.; Li, Y.; Hu, Z.; Xia, J. Orexin A attenuates the sleep-promoting effect of adenosine in the lateral hypothalamus of rats. Neurosci. Bull. 2014, 30, 877–886. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.R.; Sun, Y.; Luo, Y.J.; Zhao, X.; Chen, J.F.; Yanagawa, Y.; Qu, W.M.; Huang, Z.L. Paeoniflorin Promotes Non-rapid Eye Movement Sleep via Adenosine A1 Receptors. J. Pharmacol. Exp. Ther. 2016, 356, 64–73. [Google Scholar] [CrossRef]
- Neubauer, D.N.; Pandi-Perumal, S.R.; Spence, D.W.; Buttoo, K.; Monti, J.M. Pharmacotherapy of Insomnia. J. Cent. Nerv. Syst. Dis. 2018, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bambury, A.; Sandhu, K.; Cryan, J.F.; Dinan, T.G. Finding the needle in the haystack: Systematic identification of psychobiotics. Br. J. Pharmacol. 2018, 175, 4430–4438. [Google Scholar] [CrossRef]
- Herman, L.; Chemaly, M.; Cocconcelli, P.S.; Fernandez, P.; Klein, G.; Peixe, L.; Prieto, M.; Querol, A.; Suarez, J.E.; Sundh, I.; et al. The qualified presumption of safety assessment and its role in EFSA risk evaluations: 15 years past. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′→3′) | Reference | |
|---|---|---|---|
| ERIC | ATGTAAGCTCCTGGGGATTCAC | [26] | |
| AAGTAAGTGACTGGGGTGAGCG | |||
| BOX | CTACGGCAAGGCGACGCTGACG | [16] | |
| (GTG)5 | GTGGTGGTGGTGGTG | [27] | |
| RAPD | CTCAGGTCGC | Present work | |
| For qRT-PCR | Sequence (5′→3′) | Size (bp) | Accession number |
| A1R-F | AGAACCACCTCCACCCTTCT | 227 | XM_006529079.2 |
| A1R-R | TACTCTGGGTGGTGGTCACA | ||
| A2AR-F | AACCTGCAGAACGTCAC | 245 | XM_006513093.3 |
| A2AR-R | GTCACCAAGCCATTGTACCG | ||
| NT5e-F | TTACTAAAGCATGACTCTGGTGATCAA | 84 | NM_011851.4 |
| NT5e-R | AACGGCTGGGTAAACTACTTTCATT | ||
| GAPDH-F | CAATGTGTCCGTCGTGGATCT | 208 | XM_017321385.1 |
| GAPDH-R | GTCCTCAGTGTAGCCCAAGATG | ||
| Gene Name | Basal Forebrain | Hypothalamus | ||||||
|---|---|---|---|---|---|---|---|---|
| Veh | PS150 | Veh + Caffeine | PS150 + Caffeine | Veh | PS150 | Veh + Caffeine | PS150 + Caffeine | |
| A1R | 1.00 ± 0.13 | 0.91 ± 0.06 | 0.86 ± 0.17 | 1.00 ± 0.15 | 1.00 ± 0.11 | 1.38 ± 0.19 * | 1.08 ± 0.18 | 1.03 ± 0.11 |
| A2AR | 1.00 ± 0.21 | 0.90 ± 0.46 | 1.50 ± 0.3 | 1.39 ± 0.28 | 1.00 ± 0.14 | 0.95 ± 0.23 | 0.88 ± 0.15 | 0.89 ± 0.22 |
| NT5e | 1.00 ± 0.15 | 0.83 ± 0.19 | 0.90 ± 0.15 | 0.97 ± 0.22 | 1.00 ± 0.06 | 1.02 ± 0.23 | 1.09 ± 0.17 | 1.12 ± 0.23 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, A.; Shih, C.-T.; Huang, C.-L.; Wu, C.-C.; Lin, C.-T.; Tsai, Y.-C. Hypnotic Effects of Lactobacillus fermentum PS150TM on Pentobarbital-Induced Sleep in Mice. Nutrients 2019, 11, 2409. https://doi.org/10.3390/nu11102409
Lin A, Shih C-T, Huang C-L, Wu C-C, Lin C-T, Tsai Y-C. Hypnotic Effects of Lactobacillus fermentum PS150TM on Pentobarbital-Induced Sleep in Mice. Nutrients. 2019; 11(10):2409. https://doi.org/10.3390/nu11102409
Chicago/Turabian StyleLin, Alexander, Ching-Ting Shih, Chin-Lin Huang, Chien-Chen Wu, Ching-Ting Lin, and Ying-Chieh Tsai. 2019. "Hypnotic Effects of Lactobacillus fermentum PS150TM on Pentobarbital-Induced Sleep in Mice" Nutrients 11, no. 10: 2409. https://doi.org/10.3390/nu11102409
APA StyleLin, A., Shih, C. -T., Huang, C. -L., Wu, C. -C., Lin, C. -T., & Tsai, Y. -C. (2019). Hypnotic Effects of Lactobacillus fermentum PS150TM on Pentobarbital-Induced Sleep in Mice. Nutrients, 11(10), 2409. https://doi.org/10.3390/nu11102409