Plasma Free Amino Acid Responses to Whey Protein and Their Relationships with Gastric Emptying, Blood Glucose- and Appetite-Regulatory Hormones and Energy Intake in Lean Healthy Men

 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Outline
2.3. Protein Drinks
2.4. Protocol
2.5. Measurements
2.5.1. Gastric Emptying
2.5.2. Plasma Ghrelin, CCK, GLP-1, Insulin, Glucagon, Free AA and Blood Glucose Concentrations
2.5.3. Energy Intake
2.6. Data and Statistical Analysis
3. Results
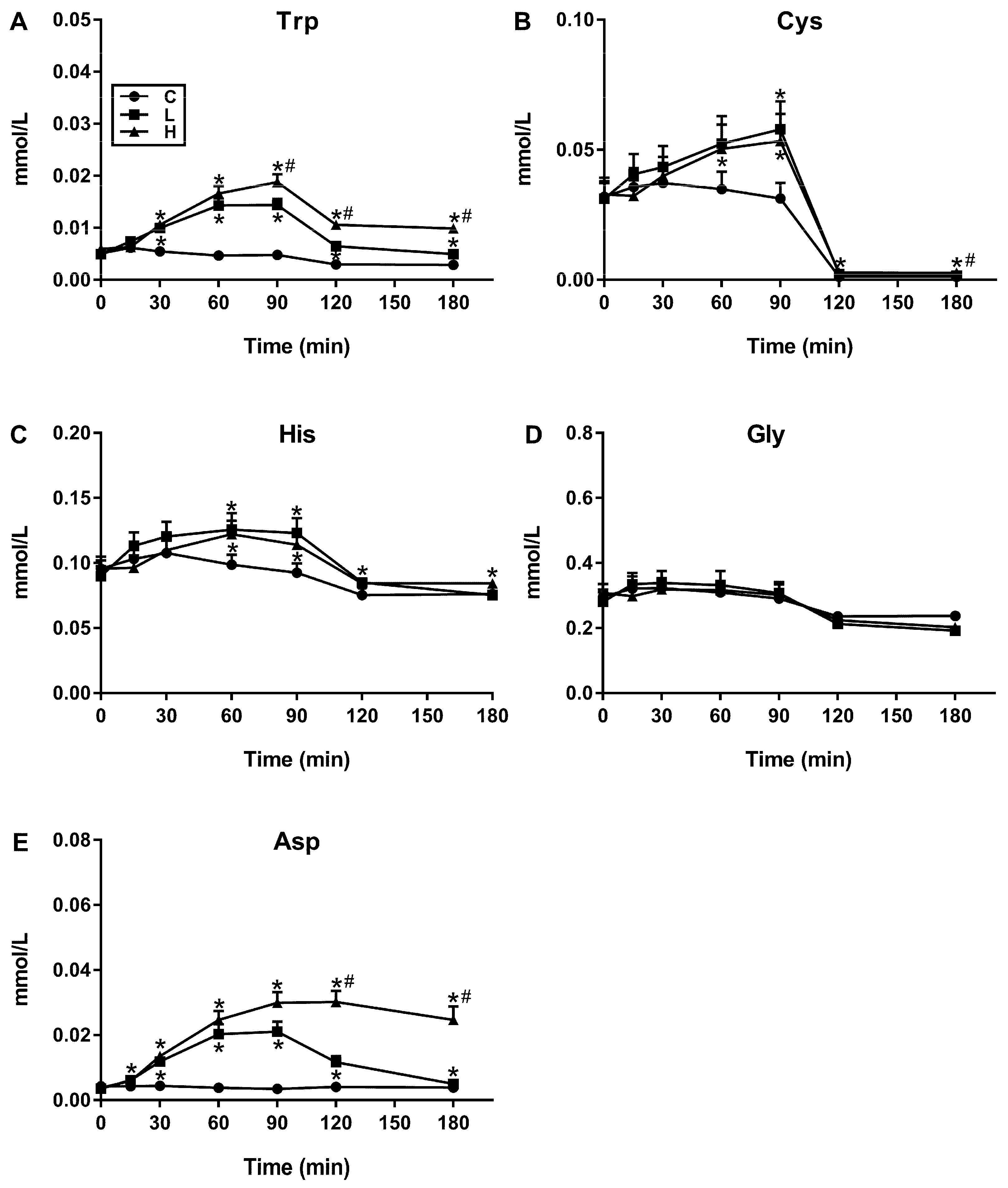
3.1. Plasma AA Concentrations in Response to Increasing Protein Loads
3.2. Relationships between Gastric Emptying, Ghrelin, CCK, GLP-1, Insulin, Glucagon, Blood Glucose and Energy Intake with Plasma AA concentrations
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Treatment | |||
|---|---|---|---|
| AA | L | H | Per 100 g |
| g [mmol/L] | g [mmol/L] | g | |
| Glu | 5.7 [88] | 13.2 [211] | 18.9 |
| Leu | 4.3 [77] | 10.1 [181] | 14.4 |
| Asp | 4.0 [69] | 9.2 [163] | 13.2 |
| Lys | 3.8 [59] | 8.8 [142] | 12.5 |
| Ile | 2.0 [35] | 4.6 [83] | 6.6 |
| Val | 1.8 [35] | 4.1 [82] | 5.9 |
| Ala | 1.8 [46] | 4.1 [108] | 5.9 |
| Thr | 1.7 [33] | 3.9 [77] | 5.5 |
| Pro | 1.4 [28] | 3.4 [70] | 4.8 |
| Tyr | 1.3 [16] | 3.0 [39] | 4.3 |
| Ser | 1.2 [26] | 2.9 [65] | 4.1 |
| Phe | 1.2 [17] | 2.8 [40] | 4.0 |
| Cys | 1.2 [23] | 2.8 [54] | 4.0 |
| Arg | 0.9 [12] | 2.2 [30] | 3.1 |
| Met | 0.8 [12] | 1.9 [30] | 2.7 |
| Trp | 0.8 [9] | 1.9 [22] | 2.7 |
| His | 0.7 [10] | 1.5 [23] | 2.2 |
| Gly | 0.6 [18] | 1.4 [44] | 2.0 |
| Gln | Not reported | ||
| Asn | Not reported | ||
| Total | 35.04 | 81.76 | 116.8 |
| AA | R2 Value | P Value |
|---|---|---|
| mmol/L | ||
| Glu (NE) | 0.66 | <0.001 |
| Leu (E) | 0.84 | <0.001 |
| Asp (NE) | 0.83 | <0.001 |
| Lys (E) | 0.76 | <0.001 |
| Ile (E) | 0.84 | <0.001 |
| Val (E) | 0.66 | <0.001 |
| Ala (NE) | 0.26 | 0.003 |
| Thr (E) | 0.61 | <0.001 |
| Pro (C) | 0.31 | 0.001 |
| Tyr (C) | 0.72 | <0.001 |
| Ser (C) | 0.35 | <0.001 |
| Phe (E) | 0.54 | <0.001 |
| Cys (C) | 0.14 | 0.034 |
| Arg (C) | 0.51 | <0.001 |
| Met (E) | 0.74 | <0.001 |
| Trp (E) | 0.74 | <0.001 |
| His (E) | 0.11 | 0.062 |
| Gly (C) | 0 | 0.874 |
| Gln (C) | Not reported | |
| Asn (NE) | Not reported | |
| C | L | H | P Valueb | |
|---|---|---|---|---|
| Ghrelin (pmol/L 180 min·L−1) | 80,372 ± 10,256 | 66,690 ± 8536 | 59,053 ± 7854 | <0.01 |
| CCK (pmol/L 180 min·L−1) | 729 ± 102 | 1016 ± 131 | 1215 ± 119 | <0.001 |
| GLP-1 (pmol/L 180 min·L−1) | 4564 ± 439 | 6018 ± 408 | 7247 ± 470 | <0.001 |
| Insulin (mU/L 180 min·L−1) | 503 ± 63 | 1463 ± 175 | 2275 ± 300 | <0.01 |
| Glucagon (pmol/L 180 min·L−1) | 3187 ± 158 | 5973 ± 308 | 8219 ± 376 | <0.001 |
| T50 (min) | 12 ± 0.5 | 26 ± 3 c | 65 ± 9 c,d | <0.001 |
| Gastric emptying rate (kcal/min) | NA | 2.6 ± 0.3 | 2.9 ± 0.3 | >0.05 |
| Blood glucose (mmol/L 180 min·L−1) | 932 ± 16 | 921 ± 19 | 900 ± 13 | >0.05 |
| Energy intake (kcal) | 1174 ± 91 | 1027 ± 81 c | 998 ± 71 c | <0.01 |
| Amount eaten (g) | 1097 ± 78 | 1114 ± 112 | 1068 ± 69 | >0.05 |
References
- Farnsworth, E.; Luscombe, N.D.; Noakes, M.; Wittert, G.; Argyiou, E.; Clifton, P.M. Effect of a high-protein, energy-restricted diet on body composition, glycemic control, and lipid concentrations in overweight and obese hyperinsulinemic men and women. Am. J. Clin. Nutr. 2003, 78, 31–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, B.; Noakes, M.; Luscombe, N.; Clifton, P. Effect of a high-protein, high-monounsaturated fat weight loss diet on glycemic control and lipid levels in type 2 diabetes. Diabetes Care 2002, 25, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Brennan, I.M.; Luscombe-Marsh, N.D.; Seimon, R.V.; Otto, B.; Horowitz, M.; Wishart, J.M.; Feinle-Bisset, C. Effects of fat, protein, and carbohydrate and protein load on appetite, plasma cholecystokinin, peptide YY, and ghrelin, and energy intake in lean and obese men. Am. J. Physiol. 2012, 303, G129–G140. [Google Scholar] [CrossRef] [PubMed]
- Stonehouse, W.; Wycherley, T.; Luscombe-Marsh, N.; Taylor, P.; Brinkworth, G.; Riley, M. Dairy intake enhances body weight and composition changes during energy restriction in 18–50-year-old adults—A meta-analysis of randomized controlled trials. Nutrients 2016, 8, 394. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Stevens, J.E.; Cukier, K.; Maddox, A.F.; Wishart, J.M.; Jones, K.L.; Clifton, P.M.; Horowitz, M.; Rayner, C.K. Effects of a protein preload on gastric emptying, glycemia, and gut hormones after a carbohydrate meal in diet-controlled type 2 diabetes. Diabetes Care 2009, 32, 1600–1602. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.H.; Tecimer, S.N.; Shah, D.; Zafar, T.A. Protein source, quantity, and time of consumption determine the effect of proteins on short-term food intake in young men. J. Nutr. 2004, 134, 3011–3015. [Google Scholar] [CrossRef]
- Pal, S.; Ellis, V. The acute effects of four protein meals on insulin, glucose, appetite and energy intake in lean men. Br. J. Nutr. 2010, 104, 1241–1248. [Google Scholar] [CrossRef]
- Gunnerud, U.J.; Ostman, E.M.; Bjorck, I.M. Effects of whey proteins on glycaemia and insulinaemia to an oral glucose load in healthy adults; a dose-response study. Eur. J. Clin. Nutr. 2013, 67, 749–753. [Google Scholar] [CrossRef]
- Akhavan, T.; Luhovyy, B.L.; Brown, P.H.; Cho, C.E.; Anderson, G.H. Effect of premeal consumption of whey protein and its hydrolysate on food intake and postmeal glycemia and insulin responses in young adults. Am. J. Clin. Nutr. 2010, 91, 966–975. [Google Scholar] [CrossRef]
- Trico, D.; Frascerra, S.; Baldi, S.; Mengozzi, A.; Nesti, L.; Mari, A.; Natali, A. The insulinotropic effect of a high-protein nutrient preload is mediated by the increase of plasma amino acids in type 2 diabetes. Eur. J. Nutr. 2018. [Google Scholar] [CrossRef]
- King, D.G.; Walker, M.; Campbell, M.D.; Breen, L.; Stevenson, E.J.; West, D.J. A small dose of whey protein co-ingested with mixed-macronutrient breakfast and lunch meals improves postprandial glycemia and suppresses appetite in men with type 2 diabetes: A randomized controlled trial. Am. J. Clin. Nutr. 2018, 107, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Frid, A.H.; Nilsson, M.; Holst, J.J.; Bjorck, I.M. Effect of whey on blood glucose and insulin responses to composite breakfast and lunch meals in type 2 diabetic subjects. Am. J. Clin. Nutr. 2005, 82, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Calbet, J.A.; MacLean, D.A. Plasma glucagon and insulin responses depend on the rate of appearance of amino acids after ingestion of different protein solutions in humans. J. Nutr. 2002, 132, 2174–2182. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.; Holst, J.J.; Bjorck, I.M. Metabolic effects of amino acid mixtures and whey protein in healthy subjects: Studies using glucose-equivalent drinks. Am. J. Clin. Nutr. 2007, 85, 996–1004. [Google Scholar] [CrossRef]
- Lindgren, O.; Pacini, G.; Tura, A.; Holst, J.J.; Deacon, C.F.; Ahren, B. Incretin effect after oral amino acid ingestion in humans. J. Clin. Endocrinol. Metab. 2015, 100, 1172–1176. [Google Scholar] [CrossRef]
- Luscombe-Marsh, N.D.; Hutchison, A.T.; Soenen, S.; Steinert, R.E.; Clifton, P.M.; Horowitz, M.; Feinle-Bisset, C. Plasma free amino acid responses to intraduodenal whey protein, and relationships with insulin, glucagon-like peptide-1 and energy intake in lean healthy men. Nutrients 2016, 8, 4. [Google Scholar] [CrossRef]
- Hutchison, A.T.; Piscitelli, D.; Horowitz, M.; Jones, K.L.; Clifton, P.M.; Standfield, S.; Hausken, T.; Feinle-Bisset, C.; Luscombe-Marsh, N.D. Acute load-dependent effects of oral whey protein on gastric emptying, gut hormone release, glycemia, appetite, and energy intake in healthy men. Am. J. Clin. Nutr. 2015, 102, 1574–1584. [Google Scholar] [CrossRef]
- Ryan, A.T.; Feinle-Bisset, C.; Kallas, A.; Wishart, J.M.; Clifton, P.M.; Horowitz, M.; Luscombe-Marsh, N.D. Intraduodenal protein modulates antropyloroduodenal motility, hormone release, glycemia, appetite, and energy intake in lean men. Am. J. Clin. Nutr. 2012, 96, 474–482. [Google Scholar] [CrossRef] [Green Version]
- Nair, N.S.; Brennan, I.M.; Little, T.J.; Gentilcore, D.; Hausken, T.; Jones, K.L.; Wishart, J.M.; Horowitz, M.; Feinle-Bisset, C. Reproducibility of energy intake, gastric emptying, blood glucose, plasma insulin and cholecystokinin responses in healthy young males. Br. J. Nutr. 2009, 101, 1094–1102. [Google Scholar] [CrossRef]
- Brennan, I.M.; Feltrin, K.L.; Nair, N.S.; Hausken, T.; Little, T.J.; Gentilcore, D.; Wishart, J.M.; Jones, K.L.; Horowitz, M.; Feinle-Bisset, C. Effects of the phases of the menstrual cycle on gastric emptying, glycemia, plasma GLP-1 and insulin, and energy intake in healthy lean women. Am. J. Physiol. 2009, 297, G602–G610. [Google Scholar] [CrossRef] [Green Version]
- Stunkard, A.J.; Messick, S. The three-factor eating questionnaire to measure dietary restraint, disinhibition and hunger. J. Psychosom. Res. 1985, 29, 71–83. [Google Scholar] [CrossRef]
- Maddern, G.J.; Chatterton, B.E.; Collins, P.J.; Horowitz, M.; Shearman, D.J.; Jamieson, G.G. Solid and liquid gastric emptying in patients with gastro-oesophageal reflux. Br. J. Surg. 1985, 72, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Vist, G.E.; Maughan, R.J. The effect of osmolality and carbohydrate content on the rate of gastric emptying of liquids in man. J. Physiol. 1995, 486 Pt 2, 523–531. [Google Scholar] [CrossRef]
- Steinert, R.E.; Ullrich, S.S.; Geary, N.; Asarian, L.; Bueter, M.; Horowitz, M.; Feinle-Bisset, C. Comparative effects of intraduodenal amino acid infusions on food intake and gut hormone release in healthy males. Physiol. Rep. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Parker, B.A.; Doran, S.; Wishart, J.; Horowitz, M.; Chapman, I.M. Effects of small intestinal and gastric glucose administration on the suppression of plasma ghrelin concentrations in healthy older men and women. Clin. Endocrinol. 2005, 62, 539–546. [Google Scholar] [CrossRef]
- MacIntosh, C.G.; Morley, J.E.; Wishart, J.; Morris, H.; Jansen, J.B.; Horowitz, M.; Chapman, I.M. Effect of exogenous cholecystokinin (CCK)-8 on food intake and plasma CCK, leptin, and insulin concentrations in older and young adults: Evidence for increased CCK activity as a cause of the anorexia of aging. J. Clin. Endocrinol. Metab. 2001, 86, 5830–5837. [Google Scholar] [CrossRef]
- Pilichiewicz, A.N.; Chaikomin, R.; Brennan, I.M.; Wishart, J.M.; Rayner, C.K.; Jones, K.L.; Smout, A.J.; Horowitz, M.; Feinle-Bisset, C. Load-dependent effects of duodenal glucose on glycemia, gastrointestinal hormones, antropyloroduodenal motility, and energy intake in healthy men. Am. J. Physiol. 2007, 293, E743–E753. [Google Scholar] [CrossRef] [Green Version]
- Bland, J.M.; Altman, D.G. Calculating correlation coefficients with repeated observations: Part 2—Correlation between subjects. BMJ 1995, 310, 633. [Google Scholar] [CrossRef]
- Hegarty, J.E.; Fairclough, P.D.; Moriarty, K.J.; Clark, M.L.; Kelly, M.J.; Dawson, A.M. Comparison of plasma and intraluminal amino acid profiles in man after meals containing a protein hydrolysate and equivalent amino acid mixture. Gut 1982, 23, 670–674. [Google Scholar] [CrossRef]
- Adibi, S.A.; Gray, S.J. Intestinal absorption of essential amino acids in man. Gastroenterology 1967, 52, 837–845. [Google Scholar] [CrossRef]
- Matthews, D.M. Intestinal absorption of amino acids and peptides. Z. Ernahrungswiss. Suppl. 1977, 20, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Jordi, J.; Herzog, B.; Camargo, S.M.; Boyle, C.N.; Lutz, T.A.; Verrey, F. Specific amino acids inhibit food intake via the area postrema or vagal afferents. J. Physiol. 2013, 591, 5611–5621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Guo, K.; LeBlanc, R.E.; Loh, D.; Schwartz, G.J.; Yu, Y.H. Increasing dietary leucine intake reduces diet-induced obesity and improves glucose and cholesterol metabolism in mice via multimechanisms. Diabetes 2007, 56, 1647–1654. [Google Scholar] [CrossRef] [PubMed]
- Steinert, R.E.; Landrock, M.F.; Ullrich, S.S.; Standfield, S.; Otto, B.; Horowitz, M.; Feinle-Bisset, C. Effects of intraduodenal infusion of the branched-chain amino acid leucine on ad libitum eating, gut motor and hormone functions, and glycemia in healthy men. Am. J. Clin. Nutr. 2015, 102, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Elovaris, R.A.; Fitzgerald, P.C.E.; Bitarafan, V.; Ullrich, S.S.; Horowitz, M.; Feinle-Bisset, C. Intraduodenal administration of L-valine has no effect on antropyloroduodenal pressures, plasma cholecystokinin concentrations or energy intake in healthy, lean men. Nutrients 2019, 11, 99. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, S.S.; Fitzgerald, P.C.; Nkamba, I.; Steinert, R.E.; Horowitz, M.; Feinle-Bisset, C. Intragastric lysine lowers the circulating glucose and insulin responses to a mixed-nutrient drink without slowing gastric emptying in healthy adults. J. Nutr. 2017, 147, 1275–1281. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, S.S.; Fitzgerald, P.C.E.; Giesbertz, P.; Steinert, R.E.; Horowitz, M.; Feinle-Bisset, C. Effects of intragastric administration of tryptophan on the blood glucose response to a nutrient drink and energy intake, in lean and obese men. Nutrients 2018, 10, 463. [Google Scholar] [CrossRef]
- Steinert, R.E.; Luscombe-Marsh, N.D.; Little, T.J.; Standfield, S.; Otto, B.; Horowitz, M.; Feinle-Bisset, C. Effects of intraduodenal infusion of L-tryptophan on ad libitum eating, antropyloroduodenal motility, glycemia, insulinemia, and gut peptide secretion in healthy men. J. Clin. Endocrinol. Metab. 2014, 99, 3275–3284. [Google Scholar] [CrossRef]
- Salehi, A.; Gunnerud, U.; Muhammed, S.J.; Ostman, E.; Holst, J.J.; Bjorck, I.; Rorsman, P. The insulinogenic effect of whey protein is partially mediated by a direct effect of amino acids and GIP on beta-cells. Nutr. Metab. 2012, 9, 48. [Google Scholar] [CrossRef]
- Eisenstein, A.B.; Strack, I. Amino acid stimulation of glucagon secretion by perifused islets of high-protein-fed rats. Diabetes 1978, 27, 370–376. [Google Scholar] [CrossRef]
- Rolls, B.J.; Kim-Harris, S.; Fischman, M.W.; Foltin, R.W.; Moran, T.H.; Stoner, S.A. Satiety after preloads with different amounts of fat and carbohydrate: Implications for obesity. Am. J. Clin. Nutr. 1994, 60, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Little, T.J.; Russo, A.; Meyer, J.H.; Horowitz, M.; Smyth, D.R.; Bellon, M.; Wishart, J.M.; Jones, K.L.; Feinle-Bisset, C. Free fatty acids have more potent effects on gastric emptying, gut hormones, and appetite than triacylglycerides. Gastroenterology 2007, 133, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Martens, M.J.; Lemmens, S.G.; Born, J.M.; Westerterp-Plantenga, M.S. A solid high-protein meal evokes stronger hunger suppression than a liquefied high-protein meal. Obesity 2011, 19, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Veldhorst, M.A.; Nieuwenhuizen, A.G.; Hochstenbach-Waelen, A.; van Vught, A.J.; Westerterp, K.R.; Engelen, M.P.; Brummer, R.J.; Deutz, N.E.; Westerterp-Plantenga, M.S. Dose-dependent satiating effect of whey relative to casein or soy. Physiol. Behav. 2009, 96, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, L.Q.; Lorenzen, J.K.; Bendsen, N.T.; Rasmussen, C.; Astrup, A. Effect of dairy proteins on appetite, energy expenditure, body weight, and composition: A review of the evidence from controlled clinical trials. Adv. Nutr. 2013, 4, 418–438. [Google Scholar] [CrossRef] [PubMed]



| Treatment | |||||
|---|---|---|---|---|---|
| AA | C | L | H | F2,30 | P Valueb |
| mmol/L | |||||
| Gln (C) | 0.80 ± 0.6 | 0.74 ± 0.07 | 0.81 ± 0.08 | 0.78 | 0.468 |
| Ala (NE) | 0.41 ± 0.03 | 0.38 ± 0.04 | 0.43 ± 0.04 | 1.41 | 0.261 |
| Gly (C) | 0.30 ± 0.02 | 0.28 ± 0.03 | 0.31 ± 0.03 | 1.15 | 0.329 |
| Val (E) | 0.28 ± 0.02 | 0.26 ± 0.02 | 0.29 ± 0.03 | 0.71 | 0.499 |
| Pro (C) | 0.26 ± 0.02 | 0.27 ± 0.04 | 0.27 ± 0.03 | 0.07 | 0.930 |
| Lys (E) | 0.20 ± 0.02 | 0.19 ± 0.02 | 0.21 ± 0.02 | 0.89 | 0.420 |
| Leu (E) | 0.14 ± 0.01 | 0.14 ± 0.01 | 0.15 ± 0.02 | 0.74 | 0.488 |
| Thr (E) | 0.14 ± 0.01 | 0.13 ± 0.01 | 0.14 ± 0.01 | 1.54 | 0.232 |
| Ser (C) | 0.12 ± 0.01 | 0.11 ± 0.01 | 0.12 ± 0.01 | 1.06 | 0.359 |
| Arg (C) | 0.10 ± 0.01 | 0.09 ± 0.01 | 0.10 ± 0.01 | 2.03 | 0.148 |
| His (E) | 0.10 ± 0.01 | 0.09 ± 0.01 | 0.10 ± 0.01 | 0.45 | 0.639 |
| Ile (E) | 0.08 ± 0.01 | 0.07 ± 0.01 | 0.08 ± 0.01 | 2.16 | 0.132 |
| Tyr (C) | 0.07 ± 0.01 | 0.07 ± 0.01 | 0.08 ± 0.01 | 1.53 | 0.234 |
| Asn (NE) | 0.07 ± 0.01 | 0.06 ± 0.01 | 0.07 ± 0.01 | 0.56 | 0.580 |
| Phe (E) | 0.06 ± 0.01 | 0.06 ± 0.01 | 0.06 ± 0.01 | 0.65 | 0.527 |
| Glu (NE) | 0.06 ± 0.01 | 0.05 ± 0.01 | 0.05 ± 0.01 | 1.12 | 0.341 |
| Cys (C) | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.16 | 0.850 |
| Met (E) | 0.03 ± 0.003 | 0.03 ± 0.003 | 0.03 ± 0.003 | 0.62 | 0.543 |
| Trp (E) | 0.005 ± 0.00 | 0.005 ± 0.00 | 0.006 ± 0.00 | 3.88 | 0.032 |
| Asp (NE) | 0.004 ± 0.00 | 0.004 ± 0.00 | 0.004 ± 0.00 | 1.94 | 0.162 |
| Total | 3.27 ± 0.21 | 3.06 ± 0.26 | 3.35 ± 0.28 | 0.94 | 0.404 |
| Treatment | |||||
|---|---|---|---|---|---|
| AA | C | L | H | F2,30 | P Valueb |
| mmol·180 min·L−1 | |||||
| Glu (NE) | 1.9 ± 0.4 | 8.1 ± 1.2 c | 11.2 ± 1.0 c,d | 36.8 | <0.001 |
| Leu (E) | 0.6 ± 0.2 | 48.4 ± 3.5 c | 74.4 ± 5.2 c,d | 133 | <0.001 |
| Asp (NE) | 0.1 ± 0.02 | 1.7 ± 0.2 c | 3.5 ± 0.3 c,d | 74.0 | <0.001 |
| Lys (E) | 1.7 ± 0.4 | 33.7 ± 3.1 c | 49.6 ± 4.4 c,d | 67.6 | <0.001 |
| Ile (E) | 0.3 ± 0.1 | 22.4 ± 1.7 c | 35.0 ± 2.5 c,d | 123 | <0.001 |
| Val (E) | 1.5 ± 0.5 | 30.1 ± 3.3 c | 40.3 ± 4.1 c | 45.1 | <0.001 |
| Ala (NE) | 2.3 ± 0.7 | 22.1 ± 3.0 c | 18.7 ± 3.0 c | 15.0 | 0.001 |
| Thr (E) | 1.1 ± 0.3 | 14.1 ± 1.5 c | 18.2 ± 2.0 c | 35.7 | <0.001 |
| Pro (C) | 2.1 ± 0.6 | 15.3 ± 1.8 c | 14.1 ± 2.0 c | 16.1 | <0.001 |
| Tyr (C) | 0.3 ± 0.1 | 8.4 ± 0.9 c | 12.7 ± 1.3 c,d | 51.6 | <0.001 |
| Ser (C) | 0.9 ± 0.2 | 6.9 ± 1.0 c | 7.1 ± 1.0 c | 14.6 | <0.001 |
| Phe (E) | 0.3 ± 0.1 | 3.9 ± 0.6 c | 5.3 ± 0.8 c | 22.6 | <0.001 |
| Cys (C) | 0.3 ± 0.1 | 1.7 ± 0.4 c | 1.3 ± 0.3 c | 7.43 | 0.002 |
| Arg (C) | 0.5 ± 0.1 | 6.9 ± 0.9 c | 8.2 ± 1.0 c | 24.4 | 0.001 |
| Met (E) | 0.2 ± 0.05 | 4.2 ± 0.4 c | 6.6 ± 0.7 c,d | 52.7 | <0.001 |
| Trp (E) | 0.04 ± 0.01 | 0.82 ± 0.07 c | 1.15 ± 0.1 c | 66.2 | <0.001 |
| His (E) | 0.8 ± 0.2 | 3.9 ± 0.7 c | 2.8 ± 0.4 c | 8.13 | 0.006 |
| Gly (C) | 2.6 ± 0.6 | 6.3 ± 1.4 | 3.2 ± 0.8 | 3.50 | 0.066 |
| Gln (C) | 8.5 ± 1.9 | 25.4 ± 4.8 c | 14.9 ± 3.1 | 5.30 | 0.011 |
| Asn (NE) | 0.6 ± 0.1 | 5.9 ± 0.7 c | 6.9 ± 0.9 c | 23.9 | <0.001 |
| Total | 23.2 ± 6.0 | 255.25 ± 29.0 c | 302.9 ± 34.0 c | 27.9 | <0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elovaris, R.A.; Hutchison, A.T.; Lange, K.; Horowitz, M.; Feinle-Bisset, C.; Luscombe-Marsh, N.D. Plasma Free Amino Acid Responses to Whey Protein and Their Relationships with Gastric Emptying, Blood Glucose- and Appetite-Regulatory Hormones and Energy Intake in Lean Healthy Men. Nutrients 2019, 11, 2465. https://doi.org/10.3390/nu11102465
Elovaris RA, Hutchison AT, Lange K, Horowitz M, Feinle-Bisset C, Luscombe-Marsh ND. Plasma Free Amino Acid Responses to Whey Protein and Their Relationships with Gastric Emptying, Blood Glucose- and Appetite-Regulatory Hormones and Energy Intake in Lean Healthy Men. Nutrients. 2019; 11(10):2465. https://doi.org/10.3390/nu11102465
Chicago/Turabian StyleElovaris, Rachel A., Amy T. Hutchison, Kylie Lange, Michael Horowitz, Christine Feinle-Bisset, and Natalie D. Luscombe-Marsh. 2019. "Plasma Free Amino Acid Responses to Whey Protein and Their Relationships with Gastric Emptying, Blood Glucose- and Appetite-Regulatory Hormones and Energy Intake in Lean Healthy Men" Nutrients 11, no. 10: 2465. https://doi.org/10.3390/nu11102465
APA StyleElovaris, R. A., Hutchison, A. T., Lange, K., Horowitz, M., Feinle-Bisset, C., & Luscombe-Marsh, N. D. (2019). Plasma Free Amino Acid Responses to Whey Protein and Their Relationships with Gastric Emptying, Blood Glucose- and Appetite-Regulatory Hormones and Energy Intake in Lean Healthy Men. Nutrients, 11(10), 2465. https://doi.org/10.3390/nu11102465