The Influence of Dietary Fatty Acids on Immune Responses

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Dietary Fatty Acids and Innate Immune Responses
2.1. Epithelium
2.2. Macrophages
2.3. Dendritic Cells
2.4. Neutrophils
2.5. Innate Lymphoid Cells
3. Dietary Fatty Acids and Adaptive Immune Responses
3.1. T Cells
3.2. B Cells
4. Dietary Fatty Acids in Immune-Related and Metabolic Diseases
4.1. Allergic Diseases
4.1.1. Asthma
4.1.2. Allergic Rhinitis
4.1.3. Atopic Dermatitis
4.1.4. Food Allergy
4.2. Autoimmune Diseases
4.2.1. Rheumatic Diseases
4.2.2. Multiple Sclerosis
4.2.3. Type 1 Diabetes
4.3. Type 2 Diabetes
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Arachidonic Acid |
| Akt | Protein Kinase B |
| ALA | α-Linolenic Acid |
| BM-DCs | Bone Marrow-Derived Dendritic Cells |
| CD | Cluster of Differentiation |
| COX | Cyclooxygenase |
| DCs | Dendritic Cells |
| DHA | Docosahexaenoic Acid |
| EAE | Experimental Autoimmune Encephalomyelitis |
| EPA | Eicosapentaenoic Acid |
| ERK | Extracellular Signal-Regulated Kinase |
| FAs | Fatty Acids |
| GLA | γ-Linolenic Acid |
| GPRs | G-Protein-Coupled Receptors |
| HFD | High Fat Diet |
| IBD | Inflammatory Bowel Disease |
| IFN | Interferon |
| IL | Interleukin |
| ILCs | Innate Lymphoid Cells |
| JAK | Janus Kinases |
| JNK | C-Jun N-Terminal Kinase |
| LA | Linoleic Acid |
| LPS | Lipopolysaccharide |
| MA | Myristic Acid |
| MAPK | Mitogen-Activated Protein Kinase |
| mo-DCs | Monocyte-Derived Dendritic Cells |
| MUFAs | Monounsaturated Fatty Acids |
| NLRP3 | NOD-like Receptor Protein 3 |
| OA | Oleic Acid |
| PA | Palmitic Acid |
| PGE2 | Prostaglandin E2 |
| PKC | Protein Kinase C |
| PPAR | Peroxisome Proliferator-Activated Receptor |
| PUFAs | Polyunsaturated Fatty Acids |
| ROS | Reactive Oxygen Species |
| SFAs | Saturated Fatty Acids |
| STA | Stearic Acid |
| STAT | Signal Transducer and Activator of Transcription Proteins |
| TEER | Trans Epithelial Electrical Resistance |
| TJs | Tight Junctions |
| TLRs | Toll-Like Receptors |
| UFAs | Unsaturated Fatty Acids |
References
- Fahy, E.; Subramaniam, S.; Murphy, R.C.; Nishijima, M.; Raetz, C.R.; Shimizu, T.; Spener, F.; van Meer, G.; Wakelam, M.J.; Dennis, E.A. Update of the LIPID MAPS comprehensive classification system for lipids. J. Lipid Res. 2009, 50, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwingshackl, L.; Hoffmann, G. Monounsaturated fatty acids, olive oil and health status: A systematic review and meta-analysis of cohort studies. Lipids Health Dis. 2014, 13, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anez-Bustillos, L.; Dao, D.T.; Fell, G.L.; Baker, M.A.; Gura, K.M.; Bistrian, B.R.; Puder, M. Redefining essential fatty acids in the era of novel intravenous lipid emulsions. Clin. Nutr. 2017. [Google Scholar] [CrossRef] [PubMed]
- Whelan, J.; Rust, C. Innovative dietary sources of n-3 fatty acids. Annu. Rev. Nutr. 2006, 26, 75–103. [Google Scholar] [CrossRef]
- Zarate, R.; El Jaber-Vazdekis, N.; Tejera, N.; Perez, J.A.; Rodriguez, C. Significance of long chain polyunsaturated fatty acids in human health. Clin. Transl. Med. 2017, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Le, H.D.; Meisel, J.A.; de Meijer, V.E.; Gura, K.M.; Puder, M. The essentiality of arachidonic acid and docosahexaenoic acid. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Hishikawa, D.; Valentine, W.J.; Iizuka-Hishikawa, Y.; Shindou, H.; Shimizu, T. Metabolism and functions of docosahexaenoic acid-containing membrane glycerophospholipids. FEBS Lett. 2017. [Google Scholar] [CrossRef] [Green Version]
- Brown, H.A.; Marnett, L.J. Introduction to lipid biochemistry, metabolism, and signaling. Chem. Rev. 2011, 111, 5817–5820. [Google Scholar] [CrossRef]
- Cook, H.W.; McMaster, C.R. Fatty Acid Desaturation and Chain Elongation Ineukaryotes. In Biochemistry of Lipids, Lipoproteins and Membranes, 4th ed.; Vance, D.E., Vance, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2002; Volume 36. [Google Scholar]
- Coleman, R.A.; Lee, D.P. Enzymes of triacylglycerol synthesis and their regulation. Prog. Lipid Res. 2004, 43, 134–176. [Google Scholar] [CrossRef]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.A.; Brash, A.R.; Murphy, R.C. The discovery and early structural studies of arachidonic acid. J. Lipid Res. 2016, 57, 1126–1132. [Google Scholar] [CrossRef] [Green Version]
- Vannice, G.; Rasmussen, H. Position of the academy of nutrition and dietetics: Dietary fatty acids for healthy adults. J. Acad. Nutr. Diet. 2014, 114, 136–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiecolt-Glaser, J.K.; Belury, M.A.; Andridge, R.; Malarkey, W.B.; Hwang, B.S.; Glaser, R. Omega-3 supplementation lowers inflammation in healthy middle-aged and older adults: A randomized controlled trial. Brain Behav. Immun. 2012, 26, 988–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiecolt-Glaser, J.K.; Belury, M.A.; Andridge, R.; Malarkey, W.B.; Glaser, R. Omega-3 supplementation lowers inflammation and anxiety in medical students: A randomized controlled trial. Brain Behav. Immun. 2011, 25, 1725–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muldoon, M.F.; Laderian, B.; Kuan, D.C.; Sereika, S.M.; Marsland, A.L.; Manuck, S.B. Fish oil supplementation does not lower C-reactive protein or interleukin-6 levels in healthy adults. J. Intern. Med. 2016, 279, 98–109. [Google Scholar] [CrossRef] [Green Version]
- Domingo, P.; Gallego-Escuredo, J.M.; Fernandez, I.; Villarroya, J.; Torres, F.; Del Mar Gutierrez, M.; Mateo, M.G.; Villarroya, F.; Vidal, F.; Giralt, M.; et al. Effects of docosahexanoic acid supplementation on inflammatory and subcutaneous adipose tissue gene expression in HIV-infected patients on combination antiretroviral therapy (cART). A sub-study of a randomized, double-blind, placebo-controlled study. Cytokine 2018, 105, 73–79. [Google Scholar] [CrossRef]
- Allaire, J.; Couture, P.; Leclerc, M.; Charest, A.; Marin, J.; Lepine, M.C.; Talbot, D.; Tchernof, A.; Lamarche, B. A randomized, crossover, head-to-head comparison of eicosapentaenoic acid and docosahexaenoic acid supplementation to reduce inflammation markers in men and women: The Comparing EPA to DHA (ComparED) Study. Am. J. Clin. Nutr. 2016, 104, 280–287. [Google Scholar] [CrossRef]
- Rangel-Huerta, O.D.; Aguilera, C.M.; Mesa, M.D.; Gil, A. Omega-3 long-chain polyunsaturated fatty acids supplementation on inflammatory biomakers: A systematic review of randomised clinical trials. Br. J. Nutr. 2012, 107, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Cornish, S.M.; Myrie, S.B.; Bugera, E.M.; Chase, J.E.; Turczyn, D.; Pinder, M. Omega-3 supplementation with resistance training does not improve body composition or lower biomarkers of inflammation more so than resistance training alone in older men. Nutr. Res. 2018, 60, 87–95. [Google Scholar] [CrossRef]
- Zhao, M.; Chiriboga, D.; Olendzki, B.; Xie, B.; Li, Y.; McGonigal, L.J.; Maldonado-Contreras, A.; Ma, Y. Substantial Increase in Compliance with Saturated Fatty Acid Intake Recommendations after One Year Following the American Heart Association Diet. Nutrients 2018, 10, 1486. [Google Scholar] [CrossRef] [Green Version]
- Harrison, S.; Brassard, D.; Lemieux, S.; Lamarche, B. Consumption and Sources of Saturated Fatty Acids According to the 2019 Canada Food Guide: Data from the 2015 Canadian Community Health Survey. Nutrients 2019, 11, 1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Matthan, N.R.; Manson, J.E.; Howard, B.V.; Tinker, L.F.; Neuhouser, M.L.; Van Horn, L.V.; Rossouw, J.E.; Allison, M.A.; Martin, L.W.; et al. Plasma Phospholipid Fatty Acids and Coronary Heart Disease Risk: A Matched Case-Control Study within the Women’s Health Initiative Observational Study. Nutrients 2019, 11, 1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogero, M.M.; Calder, P.C. Obesity, Inflammation, Toll-Like Receptor 4 and Fatty Acids. Nutrients 2018, 10, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venter, C.; Meyer, R.W.; Nwaru, B.I.; Roduit, C.; Untersmayr, E.; Adel-Patient, K.; Agache, I.; Agostoni, C.; Akdis, C.A.; Bischoff, S.C.; et al. EAACI position paper: Influence of dietary fatty acids on asthma, food allergy, and atopic dermatitis. Allergy 2019, 74, 1429–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venter, C.; Greenhawt, M.; Meyer, R.W.; Agostoni, C.; Reese, I.; du Toit, G.; Feeney, M.; Maslin, K.; Nwaru, B.I.; Roduit, C.; et al. EAACI position paper on diet diversity in pregnancy, infancy and childhood: Novel concepts and implications for studies in allergy and asthma. Allergy 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schleimer, R.P.; Kato, A.; Kern, R.; Kuperman, D.; Avila, P.C. Epithelium: At the interface of innate and adaptive immune responses. J. Allergy Clin. Immunol. 2007, 120, 1279–1284. [Google Scholar] [CrossRef] [Green Version]
- Salim, S.A.Y.; Söderholm, J.D. Importance of disrupted intestinal barrier in inflammatory bowel diseases. Inflamm. Bowel Dis. 2010, 17, 362–381. [Google Scholar] [CrossRef]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141. [Google Scholar] [CrossRef]
- Chan, J.K.; Roth, J.; Oppenheim, J.J.; Tracey, K.J.; Vogl, T.; Feldmann, M.; Horwood, N.; Nanchahal, J. Alarmins: Awaiting a clinical response. J. Clin. Investig. 2012, 122, 2711–2719. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, Q.; Wang, M.; Zhao, S.; Xu, G.; Li, J. n-3 polyunsaturated fatty acids prevent disruption of epithelial barrier function induced by proinflammatory cytokines. Mol. Immunol. 2008, 45, 1356–1365. [Google Scholar] [CrossRef]
- Willemsen, L.E.M.; Koetsier, M.A.; Balvers, M.; Beermann, C.; Stahl, B.; van Tol, E.A.F. Polyunsaturated fatty acids support epithelial barrier integrity and reduce IL-4 mediated permeability in vitro. Eur. J. Nutr. 2008, 47, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Beguin, P.; Errachid, A.; Larondelle, Y.; Schneider, Y.J. Effect of polyunsaturated fatty acids on tight junctions in a model of the human intestinal epithelium under normal and inflammatory conditions. Food Funct. 2013, 4, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Marion-Letellier, R.; Butler, M.; Déchelotte, P.; Playford, R.J.; Ghosh, S. Comparison of cytokine modulation by natural peroxisome proliferator–activated receptor γ ligands with synthetic ligands in intestinal-like Caco-2 cells and human dendritic cells-potential for dietary modulation of peroxisome proliferator-activated receptor γ in intestinal inflammation. Am. J. Clin. Nutr. 2008, 87, 939–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirasawa, A.; Tsumaya, K.; Awaji, T.; Katsuma, S.; Adachi, T.; Yamada, M.; Sugimoto, Y.; Miyazaki, S.; Tsujimoto, G. Free fatty acids regulate gut incretin glucagon-like peptide-1 secretion through GPR120. Nat. Med. 2005, 11, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Mobraten, K.; Haug, T.M.; Kleiveland, C.R.; Lea, T. Omega-3 and omega-6 PUFAs induce the same GPR120-mediated signalling events, but with different kinetics and intensity in Caco-2 cells. Lipids Health Dis. 2013, 12, 101. [Google Scholar] [CrossRef] [Green Version]
- Haddi, A.; Guendouz, M.; Tabet, S.A.; Mehedi, N.; Kheroua, O.; Saidi, D. Polyunsaturated fatty acids affect intestinal anaphylactic response in BALB/c mice sensitized with β-lactoglobulin. Rev. Française dAllergol. 2018, 58, 437–443. [Google Scholar] [CrossRef]
- Zhao, J.; Shi, P.; Sun, Y.; Sun, J.; Dong, J.-N.; Wang, H.-G.; Zuo, L.-G.; Gong, J.-F.; Li, Y.; Gu, L.-L.; et al. DHA protects against experimental colitis in IL-10-deficient mice associated with the modulation of intestinal epithelial barrier function. Br. J. Nutr. 2015, 114, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Whiting, C.V.; Bland, P.W.; Tarlton, J.F. Dietary N-3 Polyunsaturated Fatty Acids Reduce Disease and Colonic Proinflammatory Cytokines in a Mouse Model of Colitis. Inflamm. Bowel Dis. 2005, 11, 340–349. [Google Scholar] [CrossRef]
- Hassan, A.; Ibrahim, A.; Mbodji, K.; Coëffier, M.; Ziegler, F.; Bounoure, F.; Chardigny, J.M.; Skiba, M.; Savoye, G.; Déchelotte, P.; et al. An α-Linolenic Acid-Rich Formula Reduces Oxidative Stress and Inflammation by Regulating NF-κB in Rats with TNBS-Induced Colitis. J. Nutr. 2010, 140, 1714–1721. [Google Scholar] [CrossRef] [Green Version]
- Saedisomeolia, A.; Wood, L.G.; Garg, M.L.; Gibson, P.G.; Wark, P.A. Anti-inflammatory effects of long-chain n-3 PUFA in rhinovirus-infected cultured airway epithelial cells. Br. J. Nutr. 2009, 101, 533–540. [Google Scholar] [CrossRef] [Green Version]
- Nordgren, T.M.; Heires, A.J.; Bailey, K.L.; Katafiasz, D.M.; Toews, M.L.; Wichman, C.S.; Romberger, D.J. Docosahexaenoic acid enhances amphiregulin-mediated bronchial epithelial cell repair processes following organic dust exposure. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2018, 314, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Ghadiri, M.; Mamlouk, M.; Spicer, P.; Jarolimek, W.; Grau, G.E.; Young, P.M.; Traini, D. Effect of polyunsaturated fatty acids (PUFAs) on airway epithelial cells’ tight junction. Pulm. Pharmacol. Ther. 2016, 40. [Google Scholar] [CrossRef] [PubMed]
- Haghi, M.; Traini, D.; Wood, L.G.; Oliver, B.; Young, P.M.; Chrzanowski, W. A ‘soft spot’ for drug transport: Modulation of cell stiffness using fatty acids and its impact on drug transport in lung model. J. Mater. Chem. B 2015, 3, 2583–2589. [Google Scholar] [CrossRef]
- Shively, C.A.; Appt, S.E.; Vitolins, M.Z.; Uberseder, B.; Michalson, K.T.; Silverstein-Metzler, M.G.; Register, T.C. Mediterranean versus Western Diet Effects on Caloric Intake, Obesity, Metabolism, and Hepatosteatosis in Nonhuman Primates. Obesity 2019, 27, 777–784. [Google Scholar] [CrossRef]
- Schatz, M.; Zeiger, R.S.; Zhang, F.; Chen, W.; Yang, S.-J.; Camargo, C.A. Overweight/Obesity and Risk of Seasonal Asthma Exacerbations. J. Allergy Clin. Immunol. Pract. 2013, 1, 618–622. [Google Scholar] [CrossRef]
- Barros, R.; Moreira, A.; Fonseca, J.; Delgado, L.; Castel-Branco, M.G.; Haahtela, T.; Lopes, C.; Moreira, P. Dietary intake of α-linolenic acid and low ratio of n-6: N-3 PUFA are associated with decreased exhaled NO and improved asthma control. Br. J. Nutr. 2011, 106, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Rutting, S.; Zakarya, R.; Bozier, J.; Xenaki, D.; Horvat, J.C.; Wood, L.G.; Hansbro, P.M.; Oliver, B.G. Dietary Fatty Acids Amplify Inflammatory Responses to Infection through p38 MAPK Signaling. Am. J. Respir. Cell Mol. Biol. 2019, 60, 554–568. [Google Scholar] [CrossRef]
- Lorente-Cebrian, S.; Costa, A.G.; Navas-Carretero, S.; Zabala, M.; Laiglesia, L.M.; Martinez, J.A.; Moreno-Aliaga, M.J. An update on the role of omega-3 fatty acids on inflammatory and degenerative diseases. J. Physiol. Biochem. 2015, 71, 341–349. [Google Scholar] [CrossRef]
- Sugihara, K.; Morhardt, T.L.; Kamada, N. The Role of Dietary Nutrients in Inflammatory Bowel Disease. Front. Immunol. 2018, 9, 3183. [Google Scholar] [CrossRef]
- Limketkai, B.N.; Wolf, A.; Parian, A.M. Nutritional Interventions in the Patient with Inflammatory Bowel Disease. Gastroenterol. Clin. 2018, 47, 155–177. [Google Scholar] [CrossRef]
- Scaioli, E.; Liverani, E.; Belluzzi, A. The Imbalance between n-6/n-3 Polyunsaturated Fatty Acids and Inflammatory Bowel Disease: A Comprehensive Review and Future Therapeutic Perspectives. Int. J. Mol. Sci. 2017, 18, 2619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Plakidas, A.; Lee, W.H.; Heikkinen, A.; Chanmugam, P.; Bray, G.; Hwang, D.H. Differential modulation of Toll-like receptors by fatty acids: Preferential inhibition by n-3 polyunsaturated fatty acids. J. Lipid Res. 2003, 44, 479–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, K.L.; Lamon-Fava, S.; Matthan, N.R.; Wu, D.; Lichtenstein, A.H. EPA and DHA exposure alters the inflammatory response but not the surface expression of Toll-like receptor 4 in macrophages. Lipids 2015, 50, 121–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajnavolgyi, E.; Laczik, R.; Kun, V.; Szente, L.; Fenyvesi, E. Effects of RAMEA-complexed polyunsaturated fatty acids on the response of human dendritic cells to inflammatory signals. Beilstein J. Org. Chem. 2014, 10, 3152–3160. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Yen, J.H.; Vassiliou, E.; Adhikary, S.; Toscano, M.G.; Ganea, D. Docosahexaenoic acid prevents dendritic cell maturation and in vitro and in vivo expression of the IL-12 cytokine family. Lipids Health Dis. 2010, 9, 12. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Hao, Q.; Li, Q.R.; Yan, X.W.; Ye, S.; Li, Y.S.; Li, N.; Li, J.S. Omega-3 polyunsaturated fatty acids affect lipopolysaccharide-induced maturation of dendritic cells through mitogen-activated protein kinases p38. Nutrition 2007, 23, 474–482. [Google Scholar] [CrossRef]
- Zeyda, M.; Saemann, M.D.; Stuhlmeier, K.M.; Mascher, D.G.; Nowotny, P.N.; Zlabinger, G.J.; Waldhausl, W.; Stulnig, T.M. Polyunsaturated fatty acids block dendritic cell activation and function independently of NF-kappaB activation. J. Biol. Chem. 2005, 280, 14293–14301. [Google Scholar] [CrossRef] [Green Version]
- Paschoal, V.A.; Vinolo, M.A.; Crisma, A.R.; Magdalon, J.; Curi, R. Eicosapentaenoic (EPA) and docosahexaenoic (DHA) acid differentially modulate rat neutrophil function in vitro. Lipids 2013, 48, 93–103. [Google Scholar] [CrossRef]
- Sethi, S.; Ziouzenkova, O.; Ni, H.; Wagner, D.D.; Plutzky, J.; Mayadas, T.N. Oxidized omega-3 fatty acids in fish oil inhibit leukocyte-endothelial interactions through activation of PPAR alpha. Blood 2002, 100, 1340–1346. [Google Scholar] [CrossRef] [Green Version]
- Gorjao, R.; Cury-Boaventura, M.F.; de Lima, T.M.; Curi, R. Regulation of human lymphocyte proliferation by fatty acids. Cell Biochem. Funct. 2007, 25, 305–315. [Google Scholar] [CrossRef]
- Collison, L.W.; Collison, R.E.; Murphy, E.J.; Jolly, C.A. Dietary n-3 polyunsaturated fatty acids increase T-lymphocyte phospholipid mass and acyl-CoA binding protein expression. Lipids 2005, 40, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Terada, S.; Takizawa, M.; Yamamoto, S.; Ezaki, O.; Itakura, H.; Akagawa, K.S. Suppressive mechanisms of EPA on human T cell proliferation. Microbiol. Immunol. 2001, 45, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Zeyda, M.; Szekeres, A.B.; Saemann, M.D.; Geyeregger, R.; Stockinger, H.; Zlabinger, G.J.; Waldhausl, W.; Stulnig, T.M. Suppression of T cell signaling by polyunsaturated fatty acids: Selectivity in inhibition of mitogen-activated protein kinase and nuclear factor activation. J. Immunol. 2003, 170, 6033–6039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaudszus, A.; Gruen, M.; Watzl, B.; Ness, C.; Roth, A.; Lochner, A.; Barz, D.; Gabriel, H.; Rothe, M.; Jahreis, G. Evaluation of suppressive and pro-resolving effects of EPA and DHA in human primary monocytes and T-helper cells. J. Lipid Res. 2013, 54, 923–935. [Google Scholar] [CrossRef] [Green Version]
- Yessoufou, A.; Ple, A.; Moutairou, K.; Hichami, A.; Khan, N.A. Docosahexaenoic acid reduces suppressive and migratory functions of CD4CD25 regulatory T-cells. J. Lipid Res. 2009, 50, 2377–2388. [Google Scholar] [CrossRef] [Green Version]
- Gorjao, R.; Hirabara, S.M.; de Lima, T.M.; Cury-Boaventura, M.F.; Curi, R. Regulation of interleukin-2 signaling by fatty acids in human lymphocytes. J. Lipid Res. 2007, 48, 2009–2019. [Google Scholar] [CrossRef] [Green Version]
- Denys, A.; Hichami, A.; Khan, N.A. Eicosapentaenoic acid and docosahexaenoic acid modulate MAP kinase enzyme activity in human T-cells. Mol. Cell. Biochem. 2002, 232, 143–148. [Google Scholar] [CrossRef]
- Verlengia, R.; Gorjao, R.; Kanunfre, C.C.; Bordin, S.; de Lima, T.M.; Martins, E.F.; Newsholme, P.; Curi, R. Effects of EPA and DHA on proliferation, cytokine production, and gene expression in Raji cells. Lipids 2004, 39, 857–864. [Google Scholar] [CrossRef]
- Kawano, A.; Ariyoshi, W.; Yoshioka, Y.; Hikiji, H.; Nishihara, T.; Okinaga, T. Docosahexaenoic acid enhances M2 macrophage polarization via the p38 signaling pathway and autophagy. J. Cell. Biochem. 2019, 120, 12604–12617. [Google Scholar] [CrossRef]
- Chang, H.Y.; Lee, H.N.; Kim, W.; Surh, Y.J. Docosahexaenoic acid induces M2 macrophage polarization through peroxisome proliferator-activated receptor gamma activation. Life Sci. 2015, 120, 39–47. [Google Scholar] [CrossRef]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 Is an Omega-3 Fatty Acid Receptor Mediating Potent Anti-inflammatory and Insulin-Sensitizing Effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, E.; McGillicuddy, F.C.; Harford, K.A.; Reynolds, C.M.; Phillips, C.M.; Ferguson, J.F.; Roche, H.M. Docosahexaenoic acid attenuates macrophage-induced inflammation and improves insulin sensitivity in adipocytes-specific differential effects between LC n-3 PUFA. J. Nutr. Biochem. 2012, 23, 1192–1200. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, L.Y.; Sokolowska, M.; Eberlein, M.; Alsaaty, S.; Martinez-Anton, A.; Logun, C.; Qi, H.Y.; Shelhamer, J.H. The fish oil ingredient, docosahexaenoic acid, activates cytosolic phospholipase A via GPR120 receptor to produce prostaglandin E and plays an anti-inflammatory role in macrophages. Immunology 2014. [Google Scholar] [CrossRef]
- Kong, W.; Yen, J.H.; Ganea, D. Docosahexaenoic acid prevents dendritic cell maturation, inhibits antigen-specific Th1/Th17 differentiation and suppresses experimental autoimmune encephalomyelitis. Brain Behav. Immun. 2011, 25, 872–882. [Google Scholar] [CrossRef] [Green Version]
- Zapata-Gonzalez, F.; Rueda, F.; Petriz, J.; Domingo, P.; Villarroya, F.; Diaz-Delfin, J.; de Madariaga, M.A.; Domingo, J.C. Human dendritic cell activities are modulated by the omega-3 fatty acid, docosahexaenoic acid, mainly through PPAR(gamma):RXR heterodimers: Comparison with other polyunsaturated fatty acids. J. Leukoc. Biol. 2008, 84, 1172–1182. [Google Scholar] [CrossRef] [Green Version]
- Carlsson, J.A.; Wold, A.E.; Sandberg, A.S.; Ostman, S.M. The Polyunsaturated Fatty Acids Arachidonic Acid and Docosahexaenoic Acid Induce Mouse Dendritic Cells Maturation but Reduce T-Cell Responses In Vitro. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Bonin, A.; Khan, N.A. Regulation of calcium signalling by docosahexaenoic acid in human T-cells. Implication of CRAC channels. J. Lipid Res. 2000, 41, 277–284. [Google Scholar]
- Chow, S.C.; Jondal, M. Polyunsaturated free fatty acids stimulate an increase in cytosolic Ca2+ by mobilizing the inositol 1,4,5-trisphosphate-sensitive Ca2+ pool in T cells through a mechanism independent of phosphoinositide turnover. J. Biol. Chem. 1990, 265, 902–907. [Google Scholar]
- Aires, V.; Hichami, A.; Moutairou, K.; Khan, N.A. Docosahexaenoic acid and other fatty acids induce a decrease in pHi in Jurkat T-cells. Br. J. Pharmacol. 2003, 140, 1217–1226. [Google Scholar] [CrossRef] [Green Version]
- Guermouche, B.; Yessoufou, A.; Soulimane, N.; Merzouk, H.; Moutairou, K.; Hichami, A.; Khan, N.A. n-3 fatty acids modulate T-cell calcium signaling in obese macrosomic rats. Obes. Res. 2004, 12, 1744–1753. [Google Scholar] [CrossRef]
- Triboulot, C.; Hichami, A.; Denys, A.; Khan, N.A. Dietary (n-3) polyunsaturated fatty acids exert antihypertensive effects by modulating calcium signaling in T cells of rats. J. Nutr. 2001, 131, 2364–2369. [Google Scholar] [CrossRef]
- Wong, S.W.; Kwon, M.J.; Choi, A.M.; Kim, H.P.; Nakahira, K.; Hwang, D.H. Fatty acids modulate Toll-like receptor 4 activation through regulation of receptor dimerization and recruitment into lipid rafts in a reactive oxygen species-dependent manner. J. Biol. Chem. 2009, 284, 27384–27392. [Google Scholar] [CrossRef] [Green Version]
- Loscher, C.E.; Draper, E.; Leavy, O.; Kelleher, D.; Mills, K.H.; Roche, H.M. Conjugated linoleic acid suppresses NF-kappa B activation and IL-12 production in dendritic cells through ERK-mediated IL-10 induction. J. Immunol. 2005, 175, 4990–4998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, N.; Korenbaum, E.; Mahadeva, R.; Immenschuh, S.; Grau, V.; Dinarello, C.A.; Welte, T.; Janciauskiene, S. alpha-Linoleic acid enhances the capacity of alpha-1 antitrypsin to inhibit lipopolysaccharide induced IL-1beta in human blood neutrophils. Mol. Med. 2016, 22, 680–693. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, C.; Del Mar Cavia, M.; Roelofs, H.; Wanten, G.; Alonso-Torre, S.R. Activation of human neutrophils by oleic acid involves the production of reactive oxygen species and a rise in cytosolic calcium concentration: A comparison with N-6 polyunsaturated fatty acids. Cell. Physiol. Biochem. 2011, 28, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Cury-Boaventura, M.F.; Gorjao, R.; de Lima, T.M.; Newsholme, P.; Curi, R. Comparative toxicity of oleic and linoleic acid on human lymphocytes. Life Sci. 2006, 78, 1448–1456. [Google Scholar] [CrossRef]
- Moghaddami, N.; Costabile, M.; Grover, P.K.; Jersmann, H.P.; Huang, Z.H.; Hii, C.S.; Ferrante, A. Unique effect of arachidonic acid on human neutrophil TNF receptor expression: Up-regulation involving protein kinase C, extracellular signal-regulated kinase, and phospholipase A2. J. Immunol. 2003, 171, 2616–2624. [Google Scholar] [CrossRef]
- Lux, C.A.; Koschinski, A.; Dersch, K.; Husmann, M.; Bhakdi, S. Hypersusceptibility of neutrophil granulocytes towards lethal action of free fatty acids contained in enzyme-modified atherogenic low density lipoprotein. Atherosclerosis 2009, 207, 116–122. [Google Scholar] [CrossRef]
- Cury-Boaventura, M.F.; Gorjao, R.; de Lima, T.M.; Piva, T.M.; Peres, C.M.; Soriano, F.G.; Curi, R. Toxicity of a soybean oil emulsion on human lymphocytes and neutrophils. J. Parenter. Enteral Nutr. 2006, 30, 115–123. [Google Scholar] [CrossRef]
- Hatanaka, E.; Levada-Pires, A.C.; Pithon-Curi, T.C.; Curi, R. Systematic study on ROS production induced by oleic, linoleic, and gamma-linolenic acids in human and rat neutrophils. Free Radic. Biol. Med. 2006, 41, 1124–1132. [Google Scholar] [CrossRef]
- Padovese, R.; Curi, R. Modulation of rat neutrophil function in vitro by cis- and trans-MUFA. Br. J. Nutr. 2009, 101, 1351–1359. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.M.; Hatanaka, E.; Martins, E.F.; Oliveira, F.; Liberti, E.A.; Farsky, S.H.; Curi, R.; Pithon-Curi, T.C. Effect of oleic and linoleic acids on the inflammatory phase of wound healing in rats. Cell. Biochem. Funct. 2008, 26, 197–204. [Google Scholar] [CrossRef]
- Khan, M.A.; Pace-Asciak, C.; Al-Hassan, J.M.; Afzal, M.; Liu, Y.F.; Oommen, S.; Paul, B.M.; Nair, D.; Palaniyar, N. Furanoid F-Acid F6 Uniquely Induces NETosis Compared to C16 and C18 Fatty Acids in Human Neutrophils. Biomolecules 2018, 8, 144. [Google Scholar] [CrossRef] [Green Version]
- Suganami, T.; Tanimoto-Koyama, K.; Nishida, J.; Itoh, M.; Yuan, X.; Mizuarai, S.; Kotani, H.; Yamaoka, S.; Miyake, K.; Aoe, S.; et al. Role of the Toll-like Receptor 4/NF-κB Pathway in Saturated Fatty Acid–Induced Inflammatory Changes in the Interaction Between Adipocytes and Macrophages. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samblas, M.; Carraro, J.C.; Martínez, J.A.; Milagro, F.I. The regulation of inflammation-related genes after palmitic acid and DHA treatments is not mediated by DNA methylation. J. Physiol. Biochem. 2019. [Google Scholar] [CrossRef]
- Snodgrass, R.G.; Huang, S.; Choi, I.-W.; Rutledge, J.C.; Hwang, D.H. Inflammasome-Mediated Secretion of IL-1β in Human Monocytes through TLR2 Activation; Modulation by Dietary Fatty Acids. J. Immunol. 2013, 191, 4337–4347. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, D.A.; Zhang, K.; Hung, C.; Glasgow, S.; Aruni, A.W.; Unternaehrer, J.; Payne, K.J.; Langridge, W.H.R.; De Leon, M. Palmitic acid is a toll-like receptor 4 ligand that induces human dendritic cell secretion of IL-1beta. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Mauro, C.; Smith, J.; Cucchi, D.; Coe, D.; Fu, H.; Bonacina, F.; Baragetti, A.; Cermenati, G.; Caruso, D.; Mitro, N.; et al. Obesity-Induced Metabolic Stress Leads to Biased Effector Memory CD4(+) T Cell Differentiation via PI3K p110delta-Akt-Mediated Signals. Cell. Metab. 2017, 25, 593–609. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Wang, G.; Lyu, Y.; Wang, L.; Zuo, S.; Zou, J.; Sun, L.; Zhao, W.; Shu, C.; Yang, Y.G.; et al. Upregulation of SLAMF3 on human T cells is induced by palmitic acid through the STAT5-PI3K/Akt pathway and features the chronic inflammatory profiles of type 2 diabetes. Cell Death Dis. 2019, 10, 559. [Google Scholar] [CrossRef] [Green Version]
- Haghikia, A.; Jorg, S.; Duscha, A.; Berg, J.; Manzel, A.; Waschbisch, A.; Hammer, A.; Lee, D.H.; May, C.; Wilck, N.; et al. Dietary Fatty Acids Directly Impact Central Nervous System Autoimmunity via the Small Intestine. Immunity 2015, 43, 817–829. [Google Scholar] [CrossRef] [Green Version]
- Stentz, F.B.; Kitabchi, A.E. Palmitic acid-induced activation of human T-lymphocytes and aortic endothelial cells with production of insulin receptors, reactive oxygen species, cytokines, and lipid peroxidation. Biochem. Biophys. Res. Commun. 2006, 346, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Rockett, B.D.; Salameh, M.; Carraway, K.; Morrison, K.; Shaikh, S.R. n-3 PUFA improves fatty acid composition, prevents palmitate-induced apoptosis, and differentially modifies B cell cytokine secretion in vitro and ex vivo. J. Lipid Res. 2010, 51, 1284–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weatherill, A.R.; Lee, J.Y.; Zhao, L.; Lemay, D.G.; Youn, H.S.; Hwang, D.H. Saturated and polyunsaturated fatty acids reciprocally modulate dendritic cell functions mediated through TLR4. J. Immunol. 2005, 174, 5390–5397. [Google Scholar] [CrossRef] [PubMed]
- Hammer, A.; Schliep, A.; Jorg, S.; Haghikia, A.; Gold, R.; Kleinewietfeld, M.; Muller, D.N.; Linker, R.A. Impact of combined sodium chloride and saturated long-chain fatty acid challenge on the differentiation of T helper cells in neuroinflammation. J. Neuroinflamm. 2017, 14, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adolph, S.; Fuhrmann, H.; Schumann, J. Unsaturated fatty acids promote the phagocytosis of P. aeruginosa and R. equi by RAW264.7 macrophages. Curr. Microbiol. 2012, 65, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Saini, A.; Harjai, K.; Chhibber, S. Inhibitory effect of polyunsaturated fatty acids on apoptosis induced by Streptococcus pneumoniae in alveolar macrophages. Indian J. Med. Res. 2013, 137, 1193–1198. [Google Scholar]
- Lee, K.R.; Midgette, Y.; Shah, R. Fish Oil Derived Omega 3 Fatty Acids Suppress Adipose NLRP3 Inflammasome Signaling in Human Obesity. J. Endocr. Soc. 2019, 3, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Jiang, W.; Spinetti, T.; Tardivel, A.; Castillo, R.; Bourquin, C.; Guarda, G.; Tian, Z.; Tschopp, J.; Zhou, R. Omega-3 fatty acids prevent inflammation and metabolic disorder through inhibition of NLRP3 inflammasome activation. Immunity 2013, 38, 1154–1163. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Yang, Y.; Ou, T.; Key, C.C.; Tong, S.H.; Sequeira, R.C.; Nelson, J.M.; Nie, Y.; Wang, Z.; Boudyguina, E.; et al. Dietary PUFAs attenuate NLRP3 inflammasome activation via enhancing macrophage autophagy. J. Lipid Res. 2017, 58, 1808–1821. [Google Scholar] [CrossRef] [Green Version]
- Sanderson, P.; MacPherson, G.G.; Jenkins, C.H.; Calder, P.C. Dietary fish oil diminishes the antigen presentation activity of rat dendritic cells. J. Leukoc. Biol. 1997, 62, 771–777. [Google Scholar] [CrossRef]
- Teague, H.; Rockett, B.D.; Harris, M.; Brown, D.A.; Shaikh, S.R. Dendritic cell activation, phagocytosis and CD69 expression on cognate T cells are suppressed by n-3 long-chain polyunsaturated fatty acids. Immunology 2013, 139, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Van den Elsen, L.W.; van Esch, B.C.; Hofman, G.A.; Kant, J.; van de Heijning, B.J.; Garssen, J.; Willemsen, L.E. Dietary long chain n-3 polyunsaturated fatty acids prevent allergic sensitization to cow’s milk protein in mice. Clin. Exp. Allergy 2013, 43, 798–810. [Google Scholar] [CrossRef] [PubMed]
- Uchi, S.H.; Yanai, R.; Kobayashi, M.; Hatano, M.; Kobayashi, Y.; Yamashiro, C.; Nagai, T.; Tokuda, K.; Connor, K.M.; Sonoda, K.H.; et al. Dendritic cells mediate the anti-inflammatory action of omega-3 long-chain polyunsaturated fatty acids in experimental autoimmune uveitis. PLoS ONE 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Svahn, S.L.; Grahnemo, L.; Palsdottir, V.; Nookaew, I.; Wendt, K.; Gabrielsson, B.; Schele, E.; Benrick, A.; Andersson, N.; Nilsson, S.; et al. Dietary polyunsaturated fatty acids increase survival and decrease bacterial load during septic Staphylococcus aureus infection and improve neutrophil function in mice. Infect. Immun. 2015, 83, 514–521. [Google Scholar] [CrossRef] [Green Version]
- Arnardottir, H.H.; Freysdottir, J.; Hardardottir, I. Dietary fish oil increases the proportion of a specific neutrophil subpopulation in blood and total neutrophils in peritoneum of mice following endotoxin-induced inflammation. J. Nutr. Biochem. 2013, 24, 248–255. [Google Scholar] [CrossRef]
- Barden, A.E.; Shinde, S.; Burke, V.; Puddey, I.B.; Beilin, L.J.; Irish, A.B.; Watts, G.F.; Mori, T.A. The effect of n-3 fatty acids and coenzyme Q10 supplementation on neutrophil leukotrienes, mediators of inflammation resolution and myeloperoxidase in chronic kidney disease. Prostaglandins Other Lipid Mediat. 2018, 136, 1–8. [Google Scholar] [CrossRef]
- Barden, A.; O’Callaghan, N.; Burke, V.; Mas, E.; Beilin, L.J.; Fenech, M.; Irish, A.B.; Watts, G.F.; Puddey, I.B.; Huang, R.C.; et al. n-3 Fatty Acid Supplementation and Leukocyte Telomere Length in Patients with Chronic Kidney Disease. Nutrients 2016, 8, 175. [Google Scholar] [CrossRef] [Green Version]
- Marques, C.G.; Santos, V.C.; Levada-Pires, A.C.; Jacintho, T.M.; Gorjao, R.; Pithon-Curi, T.C.; Cury-Boaventura, M.F. Effects of DHA-rich fish oil supplementation on the lipid profile, markers of muscle damage, and neutrophil function in wheelchair basketball athletes before and after acute exercise. Appl. Physiol. Nutr. Metab. 2015, 40, 596–604. [Google Scholar] [CrossRef]
- Bonatto, S.J.; Oliveira, H.H.; Nunes, E.A.; Pequito, D.; Iagher, F.; Coelho, I.; Naliwaiko, K.; Kryczyk, M.; Brito, G.A.; Repka, J.; et al. Fish oil supplementation improves neutrophil function during cancer chemotherapy. Lipids 2012, 47, 383–389. [Google Scholar] [CrossRef]
- Duriancik, D.M.; Comstock, S.S.; Langohr, I.M.; Fenton, J.I. High levels of fish oil enhance neutrophil development and activation and influence colon mucus barrier function in a genetically susceptible mouse model. J. Nutr. Biochem. 2015, 26, 1261–1272. [Google Scholar] [CrossRef]
- McMurray, D.N.; Jolly, C.A.; Chapkin, R.S. Effects of dietary n-3 fatty acids on T cell activation and T cell receptor-mediated signaling in a murine model. J. Infect. Dis. 2000, 182, 103–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, T.Y.; Barhoumi, R.; Fan, Y.Y.; Rivera, G.M.; Hannoush, R.N.; McMurray, D.N.; Chapkin, R.S. n-3 polyunsaturated fatty acids suppress CD4(+) T cell proliferation by altering phosphatidylinositol-(4,5)-bisphosphate [PI(4,5)P2] organization. Biochim. Biophys. Acta 2016, 1858, 85–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Rux, A.H.; Ma, P.; Bdeir, K.; Sachais, B.S. Endothelial expression of E-selectin is induced by the platelet-specific chemokine platelet factor 4 through LRP in an NF-kappaB-dependent manner. Blood 2005, 105, 3545–3551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cucchi, D.; Camacho-Munoz, D.; Certo, M.; Niven, J.; Smith, J.; Nicolaou, A.; Mauro, C. Omega-3 polyunsaturated fatty acids impinge on CD4+ T cell motility and adipose tissue distribution via direct and lipid mediator-dependent effects. Cardiovasc. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Lim, K.; Kim, K.H.; Kim, J.H.; Choi, J.S.; Shim, S.C. N-3 polyunsaturated fatty acids restore Th17 and Treg balance in collagen antibody-induced arthritis. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Olson, M.V.; Liu, Y.C.; Dangi, B.; Paul Zimmer, J.; Salem, N., Jr.; Nauroth, J.M. Docosahexaenoic acid reduces inflammation and joint destruction in mice with collagen-induced arthritis. Inflamm. Res. 2013, 62, 1003–1013. [Google Scholar] [CrossRef]
- Gurzell, E.A.; Teague, H.; Harris, M.; Clinthorne, J.; Shaikh, S.R.; Fenton, J.I. DHA-enriched fish oil targets B cell lipid microdomains and enhances ex vivo and in vivo B cell function. J. Leukoc. Biol. 2013, 93, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Teague, H.; Harris, M.; Fenton, J.; Lallemand, P.; Shewchuk, B.M.; Shaikh, S.R. Eicosapentaenoic and docosahexaenoic acid ethyl esters differentially enhance B-cell activity in murine obesity. J. Lipid Res. 2014, 55, 1420–1433. [Google Scholar] [CrossRef] [Green Version]
- Teague, H.; Fhaner, C.J.; Harris, M.; Duriancik, D.M.; Reid, G.E.; Shaikh, S.R. n-3 PUFAs enhance the frequency of murine B-cell subsets and restore the impairment of antibody production to a T-independent antigen in obesity. J. Lipid Res. 2013, 54, 3130–3138. [Google Scholar] [CrossRef] [Green Version]
- Tomasdottir, V.; Thorleifsdottir, S.; Vikingsson, A.; Hardardottir, I.; Freysdottir, J. Dietary omega-3 fatty acids enhance the B1 but not the B2 cell immune response in mice with antigen-induced peritonitis. J. Nutr. Biochem. 2014, 25, 111–117. [Google Scholar] [CrossRef]
- Rockett, B.D.; Teague, H.; Harris, M.; Melton, M.; Williams, J.; Wassall, S.R.; Shaikh, S.R. Fish oil increases raft size and membrane order of B cells accompanied by differential effects on function. J. Lipid Res. 2012, 53, 674–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L.; Jin, H.; Qiang, Y.; Wu, S.; Yan, C.; Han, M.; Xiao, T.; Yan, N.; An, H.; Zhou, X.; et al. High fat diet exacerbates dextran sulfate sodium induced colitis through disturbing mucosal dendritic cell homeostasis. Int. Immunopharmacol. 2016, 40, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.M.; McGillicuddy, F.C.; Harford, K.A.; Finucane, O.M.; Mills, K.H.; Roche, H.M. Dietary saturated fatty acids prime the NLRP3 inflammasome via TLR4 in dendritic cells-implications for diet-induced insulin resistance. Mol. Nutr. Food Res. 2012, 56, 1212–1222. [Google Scholar] [CrossRef] [PubMed]
- Strandberg, L.; Verdrengh, M.; Enge, M.; Andersson, N.; Amu, S.; Onnheim, K.; Benrick, A.; Brisslert, M.; Bylund, J.; Bokarewa, M.; et al. Mice chronically fed high-fat diet have increased mortality and disturbed immune response in sepsis. PLoS ONE 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.; Strøm, M.; Maslova, E.; Dahl, R.; Hoffmann, H.J.; Rytter, D.; Bech, B.H.; Henriksen, T.B.; Granström, C.; Halldorsson, T.I. Fish oil supplementation during pregnancy and allergic respiratory disease in the adult offspring. J. Allergy Clin. Immunol. 2017, 139, 104–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perreault, M.; Roke, K.; Badawi, A.; Nielsen, D.E.; Abdelmagid, S.A.; El-Sohemy, A.; Ma, D.W.; Mutch, D.M. Plasma levels of 14:0, 16:0, 16:1n-7, and 20:3n-6 are positively associated, but 18:0 and 18:2n-6 are inversely associated with markers of inflammation in young healthy adults. Lipids 2014, 49, 255–263. [Google Scholar] [CrossRef]
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef]
- Medzhitov, R. Inflammation 2010: New adventures of an old flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Murray, P.J.; Wynn, T.A. Obstacles and opportunities for understanding macrophage polarization. J. Leukoc. Biol. 2011, 89, 557–563. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, L.; Yu, C.; Yang, X.F.; Wang, H. Monocyte and macrophage differentiation: Circulation inflammatory monocyte as biomarker for inflammatory diseases. Biomark. Res. 2014, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Sokolowska, M.; Chen, L.Y.; Eberlein, M.; Martinez-Anton, A.; Liu, Y.; Alsaaty, S.; Qi, H.Y.; Logun, C.; Horton, M.; Shelhamer, J.H. Low molecular weight hyaluronan activates cytosolic phospholipase A2alpha and eicosanoid production in monocytes and macrophages. J. Biol. Chem. 2014, 289, 4470–4488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allam-Ndoul, B.; Guénard, F.; Barbier, O.; Vohl, M.C. Effect of different concentrations of omega-3 fatty acids on stimulated THP-1 macrophages. Genes Nutr. 2017, 12, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weldon, S.M.; Mullen, A.C.; Loscher, C.E.; Hurley, L.A.; Roche, H.M. Docosahexaenoic acid induces an anti-inflammatory profile in lipopolysaccharide-stimulated human THP-1 macrophages more effectively than eicosapentaenoic acid. J. Nutr. Biochem. 2007, 18, 250–258. [Google Scholar] [CrossRef]
- He, Z.; Zhu, X.; Shi, Z.; Wu, T.; Wu, L. Metabolic Regulation of Dendritic Cell Differentiation. Front. Immunol. 2019, 10, 410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lima Thomaz, L.; Peron, G.; Oliveira, J.; da Rosa, L.C.; Thome, R.; Verinaud, L. The impact of metabolic reprogramming on dendritic cell function. Int. Immunopharmacol. 2018, 63, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Chapman, N.M.; Chi, H. Emerging Roles of Cellular Metabolism in Regulating Dendritic Cell Subsets and Function. Front. Cell. Dev. Biol. 2018, 6, 152. [Google Scholar] [CrossRef] [Green Version]
- Summers, C.; Rankin, S.M.; Condliffe, A.M.; Singh, N.; Peters, A.M.; Chilvers, E.R. Neutrophil kinetics in health and disease. Trends Immunol. 2010, 31, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, H.G.; Takeo Sato, F.; Curi, R.; Vinolo, M.A.R. Fatty acids as modulators of neutrophil recruitment, function and survival. Eur. J. Pharmacol. 2016, 785, 50–58. [Google Scholar] [CrossRef]
- Mantovani, A.; Cassatella, M.A.; Costantini, C.; Jaillon, S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat. Rev. Immunol. 2011, 11, 519–531. [Google Scholar] [CrossRef]
- Bates, E.J. Eicosanoids, fatty acids and neutrophils: Their relevance to the pathophysiology of disease. Prostaglandins Leukot. Essent. Fat. Acids 1995, 53, 75–86. [Google Scholar] [CrossRef]
- Sundqvist, M.; Christenson, K.; Holdfeldt, A.; Gabl, M.; Martensson, J.; Bjorkman, L.; Dieckmann, R.; Dahlgren, C.; Forsman, H. Similarities and differences between the responses induced in human phagocytes through activation of the medium chain fatty acid receptor GPR84 and the short chain fatty acid receptor FFA2R. Biochim. Biophys. Acta Mol. Cell. Res. 2018, 1865, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, S.; Cooper, P.R.; Potter, S.L.; Mueck, B.; Jarai, G. Cloning and expression analysis of a novel G-protein-coupled receptor selectively expressed on granulocytes. J. Leukoc. Biol. 2001, 69, 1045–1052. [Google Scholar] [PubMed]
- Wang, J.; Wu, X.; Simonavicius, N.; Tian, H.; Ling, L. Medium-chain fatty acids as ligands for orphan G protein-coupled receptor GPR84. J. Biol. Chem. 2006, 281, 34457–34464. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, H.G.; Vinolo, M.A.; Magdalon, J.; Fujiwara, H.; Cavalcanti, D.M.; Farsky, S.H.; Calder, P.C.; Hatanaka, E.; Curi, R. Dietary free oleic and linoleic acid enhances neutrophil function and modulates the inflammatory response in rats. Lipids 2010, 45, 809–819. [Google Scholar] [CrossRef]
- Hekmatdoost, A.; Wu, X.; Morampudi, V.; Innis, S.M.; Jacobson, K. Dietary oils modify the host immune response and colonic tissue damage following Citrobacter rodentium infection in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, 917–928. [Google Scholar] [CrossRef]
- Svahn, S.L.; Ulleryd, M.A.; Grahnemo, L.; Stahlman, M.; Boren, J.; Nilsson, S.; Jansson, J.O.; Johansson, M.E. Dietary Omega-3 Fatty Acids Increase Survival and Decrease Bacterial Load in Mice Subjected to Staphylococcus aureus-Induced Sepsis. Infect. Immun. 2016, 84, 1205–1213. [Google Scholar] [CrossRef] [Green Version]
- Korner, A.; Schlegel, M.; Theurer, J.; Frohnmeyer, H.; Adolph, M.; Heijink, M.; Giera, M.; Rosenberger, P.; Mirakaj, V. Resolution of inflammation and sepsis survival are improved by dietary Omega-3 fatty acids. Cell Death Differ. 2018, 25, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Mayer, K.; Fegbeutel, C.; Hattar, K.; Sibelius, U.; Kramer, H.J.; Heuer, K.U.; Temmesfeld-Wollbruck, B.; Gokorsch, S.; Grimminger, F.; Seeger, W. Omega-3 vs. omega-6 lipid emulsions exert differential influence on neutrophils in septic shock patients: Impact on plasma fatty acids and lipid mediator generation. Intensive Care Med. 2003, 29, 1472–1481. [Google Scholar] [CrossRef]
- Prescott, S.L.; Barden, A.E.; Mori, T.A.; Dunstan, J.A. Maternal fish oil supplementation in pregnancy modifies neonatal leukotriene production by cord-blood-derived neutrophils. Clin. Sci. 2007, 113, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Miles, E.A.; Banerjee, T.; Dooper, M.M.; M’Rabet, L.; Graus, Y.M.; Calder, P.C. The influence of different combinations of gamma-linolenic acid, stearidonic acid and EPA on immune function in healthy young male subjects. Br. J. Nutr. 2004, 91, 893–903. [Google Scholar] [CrossRef] [Green Version]
- Thies, F.; Miles, E.A.; Nebe-von-Caron, G.; Powell, J.R.; Hurst, T.L.; Newsholme, E.A.; Calder, P.C. Influence of dietary supplementation with long-chain n-3 or n-6 polyunsaturated fatty acids on blood inflammatory cell populations and functions and on plasma soluble adhesion molecules in healthy adults. Lipids 2001, 36, 1183–1193. [Google Scholar] [CrossRef]
- Kew, S.; Mesa, M.D.; Tricon, S.; Buckley, R.; Minihane, A.M.; Yaqoob, P. Effects of oils rich in eicosapentaenoic and docosahexaenoic acids on immune cell composition and function in healthy humans. Am. J. Clin. Nutr. 2004, 79, 674–681. [Google Scholar] [CrossRef]
- Alvarez, E.; Ruiz-Gutierrez, V.; Sobrino, F.; Santa-Maria, C. Age-related changes in membrane lipid composition, fluidity and respiratory burst in rat peritoneal neutrophils. Clin. Exp. Immunol. 2001, 124, 95–102. [Google Scholar] [CrossRef]
- Rees, D.; Miles, E.A.; Banerjee, T.; Wells, S.J.; Roynette, C.E.; Wahle, K.W.; Calder, P.C. Dose-related effects of eicosapentaenoic acid on innate immune function in healthy humans: A comparison of young and older men. Am. J. Clin. Nutr. 2006, 83, 331–342. [Google Scholar] [CrossRef] [Green Version]
- Ebbo, M.; Crinier, A.; Vely, F.; Vivier, E. Innate lymphoid cells: Major players in inflammatory diseases. Nat. Rev. Immunol. 2017, 17, 665–678. [Google Scholar] [CrossRef]
- Vivier, E.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate Lymphoid Cells: 10 Years On. Cell 2018, 174, 1054–1066. [Google Scholar] [CrossRef] [Green Version]
- Melo-Gonzalez, F.; Hepworth, M.R. Functional and phenotypic heterogeneity of group 3 innate lymphoid cells. Immunology 2017, 150, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.S.; Cella, M.; McDonald, K.G.; Garlanda, C.; Kennedy, G.D.; Nukaya, M.; Mantovani, A.; Kopan, R.; Bradfield, C.A.; Newberry, R.D.; et al. AHR drives the development of gut ILC22 cells and postnatal lymphoid tissues via pathways dependent on and independent of Notch. Nat. Immunol. 2011, 13, 144–151. [Google Scholar] [CrossRef]
- Spencer, S.P.; Wilhelm, C.; Yang, Q.; Hall, J.A.; Bouladoux, N.; Boyd, A.; Nutman, T.B.; Urban, J.F., Jr.; Wang, J.; Ramalingam, T.R.; et al. Adaptation of innate lymphoid cells to a micronutrient deficiency promotes type 2 barrier immunity. Science 2014, 343, 432–437. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.H.; Taparowsky, E.J.; Kim, C.H. Retinoic Acid Differentially Regulates the Migration of Innate Lymphoid Cell Subsets to the Gut. Immunity 2015, 43, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Pelgrom, L.R.; Everts, B. Metabolic control of type 2 immunity. Eur. J. Immunol. 2017, 47, 1266–1275. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, C.; Harrison, O.J.; Schmitt, V.; Pelletier, M.; Spencer, S.P.; Urban, J.F., Jr.; Ploch, M.; Ramalingam, T.R.; Siegel, R.M.; Belkaid, Y. Critical role of fatty acid metabolism in ILC2-mediated barrier protection during malnutrition and helminth infection. J. Exp. Med. 2016, 213, 1409–1418. [Google Scholar] [CrossRef] [PubMed]
- Emgard, J.; Kammoun, H.; Garcia-Cassani, B.; Chesne, J.; Parigi, S.M.; Jacob, J.M.; Cheng, H.W.; Evren, E.; Das, S.; Czarnewski, P.; et al. Oxysterol Sensing through the Receptor GPR183 Promotes the Lymphoid-Tissue-Inducing Function of Innate Lymphoid Cells and Colonic Inflammation. Immunity 2018, 48, 120–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berard, M.; Tough, D.F. Qualitative differences between naive and memory T cells. Immunology 2002, 106, 127–138. [Google Scholar] [CrossRef]
- Bluestone, J.A.; Mackay, C.R.; O’Shea, J.J.; Stockinger, B. The functional plasticity of T cell subsets. Nat. Rev. Immunol. 2009, 9, 811–816. [Google Scholar] [CrossRef]
- Tan, N.S.; Shaw, N.S.; Vinckenbosch, N.; Liu, P.; Yasmin, R.; Desvergne, B.; Wahli, W.; Noy, N. Selective cooperation between fatty acid binding proteins and peroxisome proliferator-activated receptors in regulating transcription. Mol. Cell. Biol. 2002, 22, 5114–5127. [Google Scholar] [CrossRef] [Green Version]
- Hughes, M.L.; Liu, B.; Halls, M.L.; Wagstaff, K.M.; Patil, R.; Velkov, T.; Jans, D.A.; Bunnett, N.W.; Scanlon, M.J.; Porter, C.J. Fatty Acid-binding Proteins 1 and 2 Differentially Modulate the Activation of Peroxisome Proliferator-activated Receptor alpha in a Ligand-selective Manner. J. Biol. Chem. 2015, 290, 13895–13906. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Tian, T.; Park, C.O.; Lofftus, S.Y.; Mei, S.; Liu, X.; Luo, C.; O’Malley, J.T.; Gehad, A.; Teague, J.E.; et al. Survival of tissue-resident memory T cells requires exogenous lipid uptake and metabolism. Nature 2017, 543, 252–256. [Google Scholar] [CrossRef] [Green Version]
- Venkataraman, C.; Kuo, F. The G-protein coupled receptor, GPR84 regulates IL-4 production by T lymphocytes in response to CD3 crosslinking. Immunol. Lett. 2005, 101, 144–153. [Google Scholar] [CrossRef]
- Jolly, C.A.; McMurray, D.N.; Chapkin, R.S. Effect of dietary n-3 fatty acids on interleukin-2 and interleukin-2 receptor alpha expression in activated murine lymphocytes. Prostaglandins Leukot. Essent. Fat. Acids 1998, 58, 289–293. [Google Scholar] [CrossRef]
- Zurier, R.B.; Rossetti, R.G.; Seiler, C.M.; Laposata, M. Human peripheral blood T lymphocyte proliferation after activation of the T cell receptor: Effects of unsaturated fatty acids. Prostaglandins Leukot. Essent. Fat. Acids 1999, 60, 371–375. [Google Scholar] [CrossRef]
- Han, S.C.; Koo, D.H.; Kang, N.J.; Yoon, W.J.; Kang, G.J.; Kang, H.K.; Yoo, E.S. Docosahexaenoic Acid Alleviates Atopic Dermatitis by Generating Tregs and IL-10/TGF-beta-Modified Macrophages via a TGF-beta-Dependent Mechanism. J. Investig. Dermatol. 2015, 135, 1556–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.C.; Kang, G.J.; Ko, Y.J.; Kang, H.K.; Moon, S.W.; Ann, Y.S.; Yoo, E.S. Fermented fish oil suppresses T helper 1/2 cell response in a mouse model of atopic dermatitis via generation of CD4+CD25+Foxp3+ T cells. BMC Immunol. 2012, 13, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gogos, C.A.; Ginopoulos, P.; Salsa, B.; Apostolidou, E.; Zoumbos, N.C.; Kalfarentzos, F. Dietary omega-3 polyunsaturated fatty acids plus vitamin E restore immunodeficiency and prolong survival for severely ill patients with generalized malignancy: A randomized control trial. Cancer 1998, 82, 395–402. [Google Scholar] [CrossRef]
- Field, C.J.; Thomson, C.A.; Van Aerde, J.E.; Parrott, A.; Euler, A.; Lien, E.; Clandinin, M.T. Lower proportion of CD45R0+ cells and deficient interleukin-10 production by formula-fed infants, compared with human-fed, is corrected with supplementation of long-chain polyunsaturated fatty acids. J. Pediatr. Gastroenterol. Nutr. 2000, 31, 291–299. [Google Scholar] [CrossRef]
- Soyland, E.; Lea, T.; Sandstad, B.; Drevon, A. Dietary supplementation with very long-chain n-3 fatty acids in man decreases expression of the interleukin-2 receptor (CD25) on mitogen-stimulated lymphocytes from patients with inflammatory skin diseases. Eur. J. Clin. Investig. 1994, 24, 236–242. [Google Scholar] [CrossRef]
- Mazurak, V.C.; Lien, V.; Field, C.J.; Goruk, S.D.; Pramuk, K.; Clandinin, M.T. Long-chain polyunsaturated fat supplementation in children with low docosahexaenoic acid intakes alters immune phenotypes compared with placebo. J. Pediatr. Gastroenterol. Nutr. 2008, 46, 570–579. [Google Scholar] [CrossRef]
- Purasiri, P.; Ashby, J.; Heys, S.D.; Eremin, O. Effect of essential fatty acids on circulating T cell subsets in patients with colorectal cancer. Cancer Immunol. Immunother. 1994, 39, 217–222. [Google Scholar] [CrossRef]
- Swanson, B.; Keithley, J.; Baum, L.; Leurgans, S.; Adeyemi, O.; Barnes, L.L.; Mata, M.; Rosdil, A. Effects of Fish Oil on HIV-Related Inflammation and Markers of Immunosenescence: A Randomized Clinical Trial. J. Altern. Complement. Med. 2018, 24, 709–716. [Google Scholar] [CrossRef] [Green Version]
- Amarasekera, M.; Noakes, P.; Strickland, D.; Saffery, R.; Martino, D.J.; Prescott, S.L. Epigenome-wide analysis of neonatal CD4(+) T-cell DNA methylation sites potentially affected by maternal fish oil supplementation. Epigenetics 2014, 9, 1570–1576. [Google Scholar] [CrossRef] [Green Version]
- Miles, E.A.; Banerjee, T.; Wells, S.J.; Calder, P.C. Limited effect of eicosapentaenoic acid on T-lymphocyte and natural killer cell numbers and functions in healthy young males. Nutrition 2006, 22, 512–519. [Google Scholar] [CrossRef]
- Sokolowska, M.; Frei, R.; Lunjani, N.; Akdis, C.A.; O’Mahony, L. Microbiome and asthma. Asthma Res. Pract. 2018, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Miles, E.; Calder, P. Can early omega-3 fatty acid exposure reduce risk of childhood allergic disease? Nutrients 2017, 9, 784. [Google Scholar] [CrossRef] [Green Version]
- Van Elten, T.M.; van Rossem, L.; Wijga, A.H.; Brunekreef, B.; de Jongste, J.C.; Koppelman, G.H.; Smit, H.A. Breast milk fatty acid composition has a long-term effect on the risk of asthma, eczema, and sensitization. Allergy 2015, 70, 1468–1476. [Google Scholar] [CrossRef]
- Bateman, E.D.; Hurd, S.S.; Barnes, P.J.; Bousquet, J.; Drazen, J.M.; FitzGerald, J.M.; Gibson, P.; Ohta, K.; O’Byrne, P.; Pedersen, S.E.; et al. Global strategy for asthma management and prevention: GINA executive summary. Eur. Respir. J. 2008, 31, 143–178. [Google Scholar] [CrossRef]
- Patel, S.; Custovic, A.; Smith, J.A.; Simpson, A.; Kerry, G.; Murray, C.S. Cross-sectional association of dietary patterns with asthma and atopic sensitization in childhood-in a cohort study. Pediatr. Allergy Immunol. 2014, 25, 565–571. [Google Scholar] [CrossRef]
- Rodriguez-Rodriguez, E.; Perea, J.M.; Jimenez, A.I.; Rodriguez-Rodriguez, P.; Lopez-Sobaler, A.M.; Ortega, R.M. Fat intake and asthma in Spanish schoolchildren. Eur. J. Clin. Nutr. 2010, 64, 1065–1071. [Google Scholar] [CrossRef]
- Hwang, I.; Cha, A.; Lee, H.; Yoon, H.; Yoon, T.; Cho, B.; Lee, S.; Park, Y. N-3 polyunsaturated fatty acids and atopy in Korean preschoolers. Lipids 2007, 42, 345–349. [Google Scholar] [CrossRef]
- Lumia, M.; Luukkainen, P.; Takkinen, H.M.; Kaila, M.; Nwaru, B.I.; Nevalainen, J.; Salminen, I.; Uusitalo, L.; Niinisto, S.; Tuokkola, J.; et al. Cow’s milk allergy and the association between fatty acids and childhood asthma risk. J. Allergy Clin. Immunol. 2014, 134, 488–490. [Google Scholar] [CrossRef]
- Ng, T.P.; Niti, M.; Yap, K.B.; Tan, W.C. Dietary and supplemental antioxidant and anti-inflammatory nutrient intakes and pulmonary function. Public Health Nutr. 2014, 17, 2081–2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, J.S.; Dockery, D.W.; Neas, L.M.; Schwartz, J.; Coull, B.A.; Raizenne, M.; Speizer, F.E. Low dietary nutrient intakes and respiratory health in adolescents. Chest 2007, 132, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Hemmelgarn, B.; Ernst, P. Airway function among Inuit primary school children in far northern Quebec. Am. J. Respir. Crit. Care Med. 1997, 156, 1870–1875. [Google Scholar] [CrossRef] [PubMed]
- Backer, V.; Nepper-Christensen, S.; Porsbjerg, C.; von Linstow, M.L.; Reersted, P.; Greenlandic Population, S. Respiratory symptoms in greenlanders living in Greenland and Denmark: A population-based study. Ann. Allergy Asthma Immunol. 2004, 93, 76–82. [Google Scholar] [CrossRef]
- Bruce, S.G.; Riediger, N.D.; Lix, L.M. Chronic disease and chronic disease risk factors among First Nations, Inuit and Metis populations of northern Canada. Chronic Dis. Inj. Can. 2014, 34, 210–217. [Google Scholar]
- Backer, V.; Baines, K.J.; Powell, H.; Porsbjerg, C.; Gibson, P.G. Increased asthma and adipose tissue inflammatory gene expression with obesity and Inuit migration to a western country. Respir. Med. 2016, 111, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Candelaria, P.V.; Backer, V.; Khoo, S.K.; Bizzintino, J.A.; Hayden, C.M.; Baynam, G.; Laing, I.A.; Zhang, G.; Porsbjerg, C.; Goldblatt, J.; et al. The importance of environment on respiratory genotype/phenotype relationships in the Inuit. Allergy 2010, 65, 229–237. [Google Scholar] [CrossRef]
- Stein, M.M.; Hrusch, C.L.; Gozdz, J.; Igartua, C.; Pivniouk, V.; Murray, S.E.; Ledford, J.G.; Marques Dos Santos, M.; Anderson, R.L.; Metwali, N.; et al. Innate Immunity and Asthma Risk in Amish and Hutterite Farm Children. N. Engl. J. Med. 2016, 375, 411–421. [Google Scholar] [CrossRef] [Green Version]
- Peters, U.; Dixon, A.E.; Forno, E. Obesity and asthma. J. Allergy Clin. Immunol. 2018, 141, 1169–1179. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Perez, N.; Schiavi, E.; Frei, R.; Ferstl, R.; Wawrzyniak, P.; Smolinska, S.; Sokolowska, M.; Sievi, N.A.; Kohler, M.; Schmid-Grendelmeier, P.; et al. Altered fatty acid metabolism and reduced stearoyl-coenzyme a desaturase activity in asthma. Allergy 2017, 72, 1744–1752. [Google Scholar] [CrossRef] [Green Version]
- Lunjani, N.; Satitsuksanoa, P.; Lukasik, Z.; Sokolowska, M.; Eiwegger, T.; O’Mahony, L. Recent developments and highlights in mechanisms of allergic diseases: Microbiome. Allergy 2018, 73, 2314–2327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolowska, M.; Akdis, C.A. Highlights in immune response, microbiome and precision medicine in allergic disease and asthma. Curr. Opin. Immunol. 2017, 48, iv–ix. [Google Scholar] [CrossRef] [PubMed]
- Bisgaard, H.; Stokholm, J.; Chawes, B.L.; Vissing, N.H.; Bjarnadóttir, E.; Schoos, A.-M.M.; Wolsk, H.M.; Pedersen, T.M.; Vinding, R.K.; Thorsteinsdóttir, S. Fish oil–derived fatty acids in pregnancy and wheeze and asthma in offspring. N. Engl. J. Med. 2016, 375, 2530–2539. [Google Scholar] [CrossRef] [PubMed]
- Best, K.P.; Sullivan, T.; Palmer, D.; Gold, M.; Kennedy, D.J.; Martin, J.; Makrides, M. Prenatal fish oil supplementation and allergy: 6-year follow-up of a randomized controlled trial. Pediatrics 2016, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Xavier, R.A.; de Barros, K.V.; de Andrade, I.S.; Palomino, Z.; Casarini, D.E.; Silveira, V.L.F. Protective effect of soybean oil-or fish oil-rich diets on allergic airway inflammation. J. Inflamm. Res. 2016, 9, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maslova, E.; Rifas-Shiman, S.L.; Oken, E.; Platts-Mills, T.A.; Gold, D.R. Fatty acids in pregnancy and risk of allergic sensitization and respiratory outcomes in childhood. Ann. Allergy Asthma Immunol. 2019, 122, 120–122. [Google Scholar] [CrossRef]
- Magnusson, J.; Ekström, S.; Kull, I.; HAakansson, N.; Nilsson, S.; Wickman, M.; Melén, E.; Risérus, U.; Bergström, A. Polyunsaturated fatty acids in plasma at 8 years and subsequent allergic disease. J. Allergy Clin. Immunol. 2018, 142, 510–516. [Google Scholar] [CrossRef] [Green Version]
- Best, K.P.; Gold, M.; Kennedy, D.; Martin, J.; Makrides, M. Omega-3 long-chain PUFA intake during pregnancy and allergic disease outcomes in the offspring: A systematic review and meta-analysis of observational studies and randomized controlled trials. Am. J. Clin. Nutr. 2016, 103, 128–143. [Google Scholar] [CrossRef] [Green Version]
- Brick, T.; Schober, Y.; Bocking, C.; Pekkanen, J.; Genuneit, J.; Loss, G.; Dalphin, J.C.; Riedler, J.; Lauener, R.; Nockher, W.A.; et al. omega-3 fatty acids contribute to the asthma-protective effect of unprocessed cow’s milk. J. Allergy Clin. Immunol. 2016, 137, 1699–1706. [Google Scholar] [CrossRef] [Green Version]
- Rucci, E.; den Dekker, H.; de Jongste, J.; Steenweg-de-Graaff, J.; Gaillard, R.; Pasmans, S.; Hofman, A.; Tiemeier, H.; Jaddoe, V.; Duijts, L. Maternal fatty acid levels during pregnancy, childhood lung function and atopic diseases. The Generation R Study. Clin. Exp. Allergy 2016, 46, 461–471. [Google Scholar] [CrossRef]
- Nagakura, T.; Matsuda, S.; Shichijyo, K.; Sugimoto, H.; Hata, K. Dietary supplementation with fish oil rich in omega-3 polyunsaturated fatty acids in children with bronchial asthma. Eur. Respir. J. 2000, 16, 861–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.C.; Yang, Y.H.; Chuang, S.Y.; Huang, S.Y.; Pan, W.H. Reduced medication use and improved pulmonary function with supplements containing vegetable and fruit concentrate, fish oil and probiotics in asthmatic school children: A randomised controlled trial. Br. J. Nutr. 2013, 110, 145–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, S.; Miyake, Y.; Sasaki, S.; Tanaka, K.; Ohya, Y.; Matsunaga, I.; Yoshida, T.; Oda, H.; Ishiko, O.; Hirota, Y.; et al. Fat and fish intake and asthma in Japanese women: Baseline data from the Osaka Maternal and Child Health Study. Int. J. Tuberc. Lung Dis. 2007, 11, 103–109. [Google Scholar] [PubMed]
- De Luis, D.A.; Armentia, A.; Aller, R.; Asensio, A.; Sedano, E.; Izaola, O.; Cuellar, L. Dietary intake in patients with asthma: A case control study. Nutrition 2005, 21, 320–324. [Google Scholar] [CrossRef]
- Kompauer, I.; Demmelmair, H.; Koletzko, B.; Bolte, G.; Linseisen, J.; Heinrich, J. Association of fatty acids in serum phospholipids with lung function and bronchial hyperresponsiveness in adults. Eur. J. Epidemiol. 2008, 23, 175–190. [Google Scholar] [CrossRef]
- Li, J.; Xun, P.; Zamora, D.; Sood, A.; Liu, K.; Daviglus, M.; Iribarren, C.; Jacobs Jr, D.; Shikany, J.M.; He, K. Intakes of long-chain omega-3 (n-3) PUFAs and fish in relation to incidence of asthma among American young adults: The CARDIA study. Am. J. Clin. Nutr. 2012, 97, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Williams, N.C.; Hunter, K.A.; Shaw, D.E.; Jackson, K.G.; Sharpe, G.R.; Johnson, M.A. Comparable reductions in hyperpnoea-induced bronchoconstriction and markers of airway inflammation after supplementation with 6.2 and 3.1 g/d of long-chain n-3 PUFA in adults with asthma. Br. J. Nutr. 2017, 117, 1379–1389. [Google Scholar] [CrossRef] [Green Version]
- Schubert, R.; Kitz, R.; Beermann, C.; Rose, M.A.; Lieb, A.; Sommerer, P.C.; Moskovits, J.; Alberternst, H.; Bohles, H.J.; Schulze, J.; et al. Effect of n-3 polyunsaturated fatty acids in asthma after low-dose allergen challenge. Int. Arch. Allergy Immunol. 2009, 148, 321–329. [Google Scholar] [CrossRef]
- Okamoto, M.; Mitsunobu, F.; Ashida, K.; Mifune, T.; Hosaki, Y.; Tsugeno, H.; Harada, S.; Tanizaki, Y. Effects of dietary supplementation with n-3 fatty acids compared with n-6 fatty acids on bronchial asthma. Intern. Med. 2000, 39, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Mickleborough, T.D.; Murray, R.L.; Ionescu, A.A.; Lindley, M.R. Fish oil supplementation reduces severity of exercise-induced bronchoconstriction in elite athletes. Am. J. Respir. Crit. Care Med. 2003, 168, 1181–1189. [Google Scholar] [CrossRef]
- Mickleborough, T.D.; Vaughn, C.L.; Shei, R.J.; Davis, E.M.; Wilhite, D.P. Marine lipid fraction PCSO-524 (lyprinol/omega XL) of the New Zealand green lipped mussel attenuates hyperpnea-induced bronchoconstriction in asthma. Respir. Med. 2013, 107, 1152–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mickleborough, T.D.; Lindley, M.R.; Ionescu, A.A.; Fly, A.D. Protective effect of fish oil supplementation on exercise-induced bronchoconstriction in asthma. Chest 2006, 129, 39–49. [Google Scholar] [CrossRef] [PubMed]
- MacRedmond, R.; Singhera, G.; Attridge, S.; Bahzad, M.; Fava, C.; Lai, Y.; Hallstrand, T.S.; Dorscheid, D.R. Conjugated linoleic acid improves airway hyper-reactivity in overweight mild asthmatics. Clin. Exp. Allergy 2010, 40, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Cazzoletti, L.; Zanolin, M.E.; Spelta, F.; Bono, R.; Chamitava, L.; Cerveri, I.; Garcia-Larsen, V.; Grosso, A.; Mattioli, V.; Pirina, P. Dietary fats, olive oil and respiratory diseases in Italian adults: A population-based study. Clin. Exp. Allergy 2019, 49, 799–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, C.S.; Simpson, B.; Kerry, G.; Woodcock, A.; Custovic, A. Dietary intake in sensitized children with recurrent wheeze and healthy controls: A nested case-control study. Allergy 2006, 61, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Wada, K.; Sahashi, Y.; Tamai, Y.; Tsuji, M.; Watanabe, K.; Ohtsuchi, S.; Ando, K.; Nagata, C. Associations of intake of antioxidant vitamins and fatty acids with asthma in pre-school children. Public Health Nutr. 2013, 16, 2040–2045. [Google Scholar] [CrossRef] [Green Version]
- Brannan, J.D.; Bood, J.; Alkhabaz, A.; Balgoma, D.; Otis, J.; Delin, I.; Dahlen, B.; Wheelock, C.E.; Nair, P.; Dahlen, S.E.; et al. The effect of omega-3 fatty acids on bronchial hyperresponsiveness, sputum eosinophilia, and mast cell mediators in asthma. Chest 2015, 147, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Moreira, A.; Moreira, P.; Delgado, L.; Fonseca, J.; Teixeira, V.; Padrao, P.; Castel-Branco, G. Pilot study of the effects of n-3 polyunsaturated fatty acids on exhaled nitric oxide in patients with stable asthma. J. Investig. Allergol. Clin. Immunol. 2007, 17, 309–313. [Google Scholar]
- Price, O.J.; Hull, J.H.; Howatson, G.; Robson-Ansley, P.; Ansley, L. Vitamin D and omega-3 polyunsaturated fatty acid supplementation in athletes with exercise-induced bronchoconstriction: A pilot study. Expert Rev. Respir. Med. 2015, 9, 369–378. [Google Scholar] [CrossRef]
- Arm, J.P.; Horton, C.E.; Spur, B.W.; Mencia-Huerta, J.M.; Lee, T.H. The effects of dietary supplementation with fish oil lipids on the airways response to inhaled allergen in bronchial asthma. Am. Rev. Respir. Dis. 1989, 139, 1395–1400. [Google Scholar] [CrossRef]
- Arm, J.P.; Horton, C.E.; Mencia-Huerta, J.M.; House, F.; Eiser, N.M.; Clark, T.J.; Spur, B.W.; Lee, T.H. Effect of dietary supplementation with fish oil lipids on mild asthma. Thorax 1988, 43, 84–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thien, F.C.; Mencia-Huerta, J.M.; Lee, T.H. Dietary fish oil effects on seasonal hay fever and asthma in pollen-sensitive subjects. Am. Rev. Respir. Dis. 1993, 147, 1138–1143. [Google Scholar] [CrossRef] [PubMed]
- Ziboh, V.A.; Naguwa, S.; Vang, K.; Wineinger, J.; Morrissey, B.M.; Watnik, M.; Gershwin, M.E. Suppression of leukotriene B4 generation by ex-vivo neutrophils isolated from asthma patients on dietary supplementation with gammalinolenic acid-containing borage oil: Possible implication in asthma. Clin. Dev. Immunol. 2004, 11, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covar, R.; Gleason, M.; Macomber, B.; Stewart, L.; Szefler, P.; Engelhardt, K.; Murphy, J.; Liu, A.; Wood, S.; DeMichele, S.; et al. Impact of a novel nutritional formula on asthma control and biomarkers of allergic airway inflammation in children. Clin. Exp. Allergy 2010, 40, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Hodge, L.; Salome, C.M.; Hughes, J.M.; Liu-Brennan, D.; Rimmer, J.; Allman, M.; Pang, D.; Armour, C.; Woolcock, A.J. Effect of dietary intake of omega-3 and omega-6 fatty acids on severity of asthma in children. Eur. Respir. J. 1998, 11, 361–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaudszus, A.; Krokowski, M.; Mockel, P.; Darcan, Y.; Avagyan, A.; Matricardi, P.; Jahreis, G.; Hamelmann, E. Cis-9,trans-11-conjugated linoleic acid inhibits allergic sensitization and airway inflammation via a PPARgamma-related mechanism in mice. J. Nutr. 2008, 138, 1336–1342. [Google Scholar] [CrossRef] [Green Version]
- Miyake, Y.; Sasaki, S.; Arakawa, M.; Tanaka, K.; Murakami, K.; Ohya, Y. Fatty acid intake and asthma symptoms in Japanese children: The Ryukyus Child Health Study. Clin. Exp. Allergy 2008, 38, 1644–1650. [Google Scholar] [CrossRef]
- Waidyatillake, N.T.; Stoney, R.; Thien, F.; Lodge, C.J.; Simpson, J.; Allen, K.; Abramson, M.; Erbas, B.; Svanes, C.; Dharmage, S. Breast milk polyunsaturated fatty acids: Associations with adolescent allergic disease and lung function. Allergy 2017, 72, 1193–1201. [Google Scholar] [CrossRef]
- Sokolowska, M.; Chen, L.Y.; Liu, Y.; Martinez-Anton, A.; Logun, C.; Alsaaty, S.; Cuento, R.A.; Cai, R.; Sun, J.; Quehenberger, O.; et al. Dysregulation of lipidomic profile and antiviral immunity in response to hyaluronan in patients with severe asthma. J. Allergy Clin. Immunol. 2017, 139, 1379–1383. [Google Scholar] [CrossRef] [Green Version]
- Magnusson, J.; Kull, I.; Westman, M.; Håkansson, N.; Wolk, A.; Melén, E.; Wickman, M.; Bergström, A. Fish and polyunsaturated fat intake and development of allergic and nonallergic rhinitis. J. Allergy Clin. Immunol. 2015, 136, 1247–1253. [Google Scholar] [CrossRef] [Green Version]
- Stratakis, N.; Roumeliotaki, T.; Oken, E.; Ballester, F.; Barros, H.; Basterrechea, M.; Cordier, S.; De Groot, R.; den Dekker, H.T.; Duijts, L. Fish and seafood consumption during pregnancy and the risk of asthma and allergic rhinitis in childhood: A pooled analysis of 18 European and US birth cohorts. Int. J. Epidemiol. 2017, 46, 1465–1477. [Google Scholar] [CrossRef] [PubMed]
- Maslova, E.; Strom, M.; Oken, E.; Campos, H.; Lange, C.; Gold, D.; Olsen, S.F. Fish intake during pregnancy and the risk of child asthma and allergic rhinitis-longitudinal evidence from the Danish National Birth Cohort. Br. J. Nutr. 2013, 110, 1313–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alm, B.; Goksor, E.; Thengilsdottir, H.; Pettersson, R.; Mollborg, P.; Norvenius, G.; Erdes, L.; Aberg, N.; Wennergren, G. Early protective and risk factors for allergic rhinitis at age 4(1/2) yr. Pediatr. Allergy Immunol. 2011, 22, 398–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nafstad, P.; Nystad, W.; Magnus, P.; Jaakkola, J.J. Asthma and allergic rhinitis at 4 years of age in relation to fish consumption in infancy. J. Asthma 2003, 40, 343–348. [Google Scholar] [CrossRef]
- Tamay, Z.; Akcay, A.; Ergin, A.; Guler, N. Effects of dietary habits and risk factors on allergic rhinitis prevalence among Turkish adolescents. Int. J. Pediatr. Otorhinolaryngol. 2013, 77, 1416–1423. [Google Scholar] [CrossRef]
- Miyake, Y.; Sasaki, S.; Tanaka, K.; Ohya, Y.; Miyamoto, S.; Matsunaga, I.; Yoshida, T.; Hirota, Y.; Oda, H.; Osaka, M.; et al. Fish and fat intake and prevalence of allergic rhinitis in Japanese females: The Osaka Maternal and Child Health Study. J. Am. Coll. Nutr. 2007, 26, 279–287. [Google Scholar] [CrossRef]
- Hoff, S.; Seiler, H.; Heinrich, J.; Kompauer, I.; Nieters, A.; Becker, N.; Nagel, G.; Gedrich, K.; Karg, G.; Wolfram, G.; et al. Allergic sensitisation and allergic rhinitis are associated with n-3 polyunsaturated fatty acids in the diet and in red blood cell membranes. Eur. J. Clin. Nutr. 2005, 59, 1071–1080. [Google Scholar] [CrossRef] [Green Version]
- Dunstan, J.A.; Mori, T.A.; Barden, A.; Beilin, L.J.; Taylor, A.L.; Holt, P.G.; Prescott, S.L. Fish oil supplementation in pregnancy modifies neonatal allergen-specific immune responses and clinical outcomes in infants at high risk of atopy: A randomized, controlled trial. J. Allergy Clin. Immunol. 2003, 112, 1178–1184. [Google Scholar] [CrossRef]
- Palmer, D.J.; Sullivan, T.; Gold, M.; Prescott, S.; Heddle, R.; Gibson, R.; Makrides, M. Effect of n-3 long chain polyunsaturated fatty acid supplementation in pregnancy on infants’ allergies in first year of life: Randomised controlled trial. BMJ 2012, 344. [Google Scholar] [CrossRef] [Green Version]
- Reese, I.; Werfel, T. Do long-chain omega-3 fatty acids protect from atopic dermatitis? J. Dtsch. Dermatol. Ges. 2015, 13, 879–885. [Google Scholar] [CrossRef] [Green Version]
- Dunder, T.; Kuikka, L.; Turtinen, J.; Rasanen, L.; Uhari, M. Diet, serum fatty acids, and atopic diseases in childhood. Allergy 2001, 56, 425–428. [Google Scholar] [CrossRef] [PubMed]
- Hoppu, U.; Rinne, M.; Lampi, A.M.; Isolauri, E. Breast milk fatty acid composition is associated with development of atopic dermatitis in the infant. J. Pediatr. Gastroenterol. Nutr. 2005, 41, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Duchen, K.; Casas, R.; Fageras-Bottcher, M.; Yu, G.; Bjorksten, B. Human milk polyunsaturated long-chain fatty acids and secretory immunoglobulin A antibodies and early childhood allergy. Pediatr. Allergy Immunol. 2000, 11, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Oddy, W.H.; Pal, S.; Kusel, M.M.; Vine, D.; de Klerk, N.H.; Hartmann, P.; Holt, P.G.; Sly, P.D.; Burton, P.R.; Stanley, F.J.; et al. Atopy, eczema and breast milk fatty acids in a high-risk cohort of children followed from birth to 5 yr. Pediatr. Allergy Immunol. 2006, 17, 4–10. [Google Scholar] [CrossRef]
- Palmer, D.; Sullivan, T.; Gold, M.; Prescott, S.; Heddle, R.; Gibson, R.; Makrides, M. Randomized controlled trial of fish oil supplementation in pregnancy on childhood allergies. Allergy 2013, 68, 1370–1376. [Google Scholar] [CrossRef]
- Noakes, P.S.; Vlachava, M.; Kremmyda, L.-S.; Diaper, N.D.; Miles, E.A.; Erlewyn-Lajeunesse, M.; Williams, A.P.; Godfrey, K.M.; Calder, P.C. Increased intake of oily fish in pregnancy: Effects on neonatal immune responses and on clinical outcomes in infants at 6 mo. Am. J. Clin. Nutr. 2012, 95, 395–404. [Google Scholar] [CrossRef]
- Solvoll, K.; Soyland, E.; Sandstad, B.; Drevon, C.A. Dietary habits among patients with atopic dermatitis. Eur. J. Clin. Nutr. 2000, 54, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Bjorneboe, A.; Soyland, E.; Bjorneboe, G.E.; Rajka, G.; Drevon, C.A. Effect of n-3 fatty acid supplement to patients with atopic dermatitis. J. Intern. Med. 1989, 731, 233–236. [Google Scholar] [CrossRef]
- Gimenez-Arnau, A.; Barranco, C.; Alberola, M.; Wale, C.; Serrano, S.; Buchanan, M.R.; Camarasa, J.G. Effects of linoleic acid supplements on atopic dermatitis. Adv. Exp. Med. Biol. 1997, 433, 285–289. [Google Scholar] [CrossRef]
- Mayser, P.; Mayer, K.; Mahloudjian, M.; Benzing, S.; Kramer, H.J.; Schill, W.B.; Seeger, W.; Grimminger, F. A double-blind, randomized, placebo-controlled trial of n-3 versus n-6 fatty acid-based lipid infusion in atopic dermatitis. J. Parenter. Enteral. Nutr. 2002, 26, 151–158. [Google Scholar] [CrossRef]
- Koch, C.; Dolle, S.; Metzger, M.; Rasche, C.; Jungclas, H.; Ruhl, R.; Renz, H.; Worm, M. Docosahexaenoic acid (DHA) supplementation in atopic eczema: A randomized, double-blind, controlled trial. Br. J. Dermatol. 2008, 158, 786–792. [Google Scholar] [CrossRef]
- Eriksen, B.B.; Kare, D.L. Open trial of supplements of omega 3 and 6 fatty acids, vitamins and minerals in atopic dermatitis. J. Dermatolog. Treat. 2006, 17, 82–85. [Google Scholar] [CrossRef]
- Soyland, E.; Funk, J.; Rajka, G.; Sandberg, M.; Thune, P.; Rustad, L.; Helland, S.; Middelfart, K.; Odu, S.; Falk, E.S.; et al. Dietary supplementation with very long-chain n-3 fatty acids in patients with atopic dermatitis. A double-blind, multicentre study. Br. J. Dermatol. 1994, 130, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Berth-Jones, J.; Graham-Brown, R.A. Placebo-controlled trial of essential fatty acid supplementation in atopic dermatitis. Lancet 1993, 341, 1557–1560. [Google Scholar] [CrossRef]
- Hoppenbrouwers, T.; Cvejic-Hogervorst, J.H.; Garssen, J.; Wichers, H.J.; Willemsen, L.E. Long Chain Polyunsaturated Fatty Acids (LCPUFAs) in the Prevention of Food Allergy. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Furuhjelm, C.; Warstedt, K.; Larsson, J.; Fredriksson, M.; Böttcher, M.F.; Fälth-Magnusson, K.; Duchén, K. Fish oil supplementation in pregnancy and lactation may decrease the risk of infant allergy. Acta Paediatr. 2009, 98, 1461–1467. [Google Scholar] [CrossRef]
- Calvani, M.; Alessandri, C.; Sopo, S.M.; Panetta, V.; Pingitore, G.; Tripodi, S.; Zappala, D.; Zicari, A.M.; Lazio Association of Pediatric Allergology Study, G. Consumption of fish, butter and margarine during pregnancy and development of allergic sensitizations in the offspring: Role of maternal atopy. Pediatr. Allergy Immunol. 2006, 17, 94–102. [Google Scholar] [CrossRef]
- Soto-Ramirez, N.; Karmaus, W.; Zhang, H.; Liu, J.; Billings, D.; Gangur, V.; Amrol, D.; da Costa, K.A.; Davis, S.; Goetzl, L. Fatty acids in breast milk associated with asthma-like symptoms and atopy in infancy: A longitudinal study. J. Asthma 2012, 49, 926–934. [Google Scholar] [CrossRef]
- Nwaru, B.I.; Erkkola, M.; Lumia, M.; Kronberg-Kippila, C.; Ahonen, S.; Kaila, M.; Ilonen, J.; Simell, O.; Knip, M.; Veijola, R.; et al. Maternal intake of fatty acids during pregnancy and allergies in the offspring. Br. J. Nutr. 2012, 108, 720–732. [Google Scholar] [CrossRef] [Green Version]
- Furuhjelm, C.; Warstedt, K.; Fageras, M.; Falth-Magnusson, K.; Larsson, J.; Fredriksson, M.; Duchen, K. Allergic disease in infants up to 2 years of age in relation to plasma omega-3 fatty acids and maternal fish oil supplementation in pregnancy and lactation. Pediatr. Allergy Immunol. 2011, 22, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Kull, I.; Bergstrom, A.; Lilja, G.; Pershagen, G.; Wickman, M. Fish consumption during the first year of life and development of allergic diseases during childhood. Allergy 2006, 61, 1009–1015. [Google Scholar] [CrossRef] [PubMed]
- Notenboom, M.L.; Mommers, M.; Jansen, E.H.; Penders, J.; Thijs, C. Maternal fatty acid status in pregnancy and childhood atopic manifestations: KOALA Birth Cohort Study. Clin. Exp. Allergy 2011, 41, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Sausenthaler, S.; Koletzko, S.; Schaaf, B.; Lehmann, I.; Borte, M.; Herbarth, O.; von Berg, A.; Wichmann, H.E.; Heinrich, J.; Group, L.S. Maternal diet during pregnancy in relation to eczema and allergic sensitization in the offspring at 2 y of age. Am. J. Clin. Nutr. 2007, 85, 530–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritzen, L.; Kjaer, T.M.; Fruekilde, M.B.; Michaelsen, K.F.; Frokiaer, H. Fish oil supplementation of lactating mothers affects cytokine production in 2 1/2-year-old children. Lipids 2005, 40, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Manley, B.J.; Makrides, M.; Collins, C.T.; McPhee, A.J.; Gibson, R.A.; Ryan, P.; Sullivan, T.R.; Davis, P.G.; Committee, D.S. High-dose docosahexaenoic acid supplementation of preterm infants: Respiratory and allergy outcomes. Pediatrics 2011, 128, 71–77. [Google Scholar] [CrossRef] [PubMed]
- D’Vaz, N.; Meldrum, S.J.; Dunstan, J.A.; Martino, D.; McCarthy, S.; Metcalfe, J.; Tulic, M.K.; Mori, T.A.; Prescott, S.L. Postnatal fish oil supplementation in high-risk infants to prevent allergy: Randomized controlled trial. Pediatrics 2012, 130, 674–682. [Google Scholar] [CrossRef] [Green Version]
- Almqvist, C.; Garden, F.; Xuan, W.; Mihrshahi, S.; Leeder, S.R.; Oddy, W.; Webb, K.; Marks, G.B. Omega-3 and omega-6 fatty acid exposure from early life does not affect atopy and asthma at age 5 years. J. Allergy Clin. Immunol. 2007, 119, 1438–1444. [Google Scholar] [CrossRef]
- Gunaratne, A.W.; Makrides, M.; Collins, C.T. Maternal prenatal and/or postnatal n-3 long chain polyunsaturated fatty acids (LCPUFA) supplementation for preventing allergies in early childhood. Cochrane Database Syst. Rev. 2015. [Google Scholar] [CrossRef]
- Lee, A.L.; Park, Y. The association between n-3 polyunsaturated fatty acid levels in erythrocytes and the risk of rheumatoid arthritis in Korean women. Ann. Nutr. Metab. 2013, 63, 88–95. [Google Scholar] [CrossRef]
- Moghaddami, M.; James, M.; Proudman, S.; Cleland, L.G. Synovial fluid and plasma n3 long chain polyunsaturated fatty acids in patients with inflammatory arthritis. Prostaglandins Leukot. Essent. Fat. Acids 2015, 97, 7–12. [Google Scholar] [CrossRef]
- Gan, R.W.; Demoruelle, M.K.; Deane, K.D.; Weisman, M.H.; Buckner, J.H.; Gregersen, P.K.; Mikuls, T.R.; O’Dell, J.R.; Keating, R.M.; Fingerlin, T.E. Omega-3 fatty acids are associated with a lower prevalence of autoantibodies in shared epitope-positive subjects at risk for rheumatoid arthritis. Ann. Rheum. Dis. 2017, 76, 147–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, R.W.; Young, K.A.; Zerbe, G.O.; Demoruelle, M.K.; Weisman, M.H.; Buckner, J.H.; Gregersen, P.K.; Mikuls, T.R.; O’Dell, J.R.; Keating, R.M. Lower omega-3 fatty acids are associated with the presence of anti-cyclic citrullinated peptide autoantibodies in a population at risk for future rheumatoid arthritis: A nested case-control study. Rheumatology 2015, 55, 367–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Giuseppe, D.; Wallin, A.; Bottai, M.; Askling, J.; Wolk, A. Long-term intake of dietary long-chain n-3 polyunsaturated fatty acids and risk of rheumatoid arthritis: A prospective cohort study of women. Ann. Rheum. Dis. 2014, 73, 1949–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proudman, S.M.; James, M.J.; Spargo, L.D.; Metcalf, R.G.; Sullivan, T.R.; Rischmueller, M.; Flabouris, K.; Wechalekar, M.D.; Lee, A.T.; Cleland, L.G. Fish oil in recent onset rheumatoid arthritis: A randomised, double-blind controlled trial within algorithm-based drug use. Ann. Rheum. Dis. 2015, 74, 89–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-H.; Bae, S.-C.; Song, G.-G. Omega-3 polyunsaturated fatty acids and the treatment of rheumatoid arthritis: A meta-analysis. Arch. Med. Res. 2012, 43, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Abdulrazaq, M.; Innes, J.K.; Calder, P.C. Effect of ω-3 polyunsaturated fatty acids on arthritic pain: A systematic review. Nutrition 2017, 39, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Raouf, J.; Idborg, H.; Englund, P.; Alexanderson, H.; Dastmalchi, M.; Jakobsson, P.-J.; Lundberg, I.E.; Korotkova, M. Targeted lipidomics analysis identified altered serum lipid profiles in patients with polymyositis and dermatomyositis. Arthritis Res. Ther. 2018, 20, 83. [Google Scholar] [CrossRef] [Green Version]
- Shikama, Y.; Kudo, Y.; Ishimaru, N.; Funaki, M. Potential Role of Free Fatty Acids in the Pathogenesis of Periodontitis and Primary Sjögren’s Syndrome. Int. J. Mol. Sci. 2017, 18, 836. [Google Scholar] [CrossRef]
- Nelson, J.W.; Leigh, N.J.; Mellas, R.E.; McCall, A.D.; Aguirre, A.; Baker, O.J. ALX/FPR2 receptor for RvD1 is expressed and functional in salivary glands. Am. J. Physiol.-Cell Physiol. 2013, 306, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Molfino, A.; Amabile, M.I.; Monti, M.; Muscaritoli, M. Omega-3 Polyunsaturated Fatty Acids in Critical Illness: Anti-Inflammatory, Proresolving, or Both? Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Felau, S.M.; Sales, L.P.; Solis, M.Y.; Hayashi, A.P.; Roschel, H.; Sa-Pinto, A.L.; Andrade, D.C.O.; Katayama, K.Y.; Irigoyen, M.C.; Consolim-Colombo, F.; et al. Omega-3 Fatty Acid Supplementation Improves Endothelial Function in Primary Antiphospholipid Syndrome: A Small-Scale Randomized Double-Blind Placebo-Controlled Trial. Front. Immunol. 2018, 9, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bäärnhielm, M.; Olsson, T.; Alfredsson, L. Fatty fish intake is associated with decreased occurrence of multiple sclerosis. Mult. Scler. J. 2014, 20, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Hoare, S.; Lithander, F.; Van Der Mei, I.; Ponsonby, A.-L.; Lucas, R.; Group, A.I. Higher intake of omega-3 polyunsaturated fatty acids is associated with a decreased risk of a first clinical diagnosis of central nervous system demyelination: Results from the Ausimmune Study. Mult. Scler. J. 2016, 22, 884–892. [Google Scholar] [CrossRef] [PubMed]
- Bjørnevik, K.; Chitnis, T.; Ascherio, A.; Munger, K.L. Polyunsaturated fatty acids and the risk of multiple sclerosis. Mult. Scler. J. 2017, 23, 1830–1838. [Google Scholar] [CrossRef]
- Sand, I.K. The role of diet in multiple sclerosis: Mechanistic connections and current evidence. Curr. Nutr. Rep. 2018, 7, 150–160. [Google Scholar] [CrossRef] [Green Version]
- Niinisto, S.; Takkinen, H.M.; Erlund, I.; Ahonen, S.; Toppari, J.; Ilonen, J.; Veijola, R.; Knip, M.; Vaarala, O.; Virtanen, S.M. Fatty acid status in infancy is associated with the risk of type 1 diabetes-associated autoimmunity. Diabetologia 2017, 60, 1223–1233. [Google Scholar] [CrossRef]
- Norris, J.M.; Kroehl, M.; Fingerlin, T.E.; Frederiksen, B.N.; Seifert, J.; Wong, R.; Clare-Salzler, M.; Rewers, M. Erythrocyte membrane docosapentaenoic acid levels are associated with islet autoimmunity: The Diabetes Autoimmunity Study in the Young. Diabetologia 2014, 57, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Jove, M.; Planavila, A.; Sanchez, R.M.; Merlos, M.; Laguna, J.C.; Vazquez-Carrera, M. Palmitate induces tumor necrosis factor-alpha expression in C2C12 skeletal muscle cells by a mechanism involving protein kinase C and nuclear factor-kappaB activation. Endocrinology 2006, 147, 552–561. [Google Scholar] [CrossRef]
- Jove, M.; Planavila, A.; Laguna, J.C.; Vazquez-Carrera, M. Palmitate-induced interleukin 6 production is mediated by protein kinase C and nuclear-factor kappaB activation and leads to glucose transporter 4 down-regulation in skeletal muscle cells. Endocrinology 2005, 146, 3087–3095. [Google Scholar] [CrossRef] [Green Version]
- Kadotani, A.; Tsuchiya, Y.; Hatakeyama, H.; Katagiri, H.; Kanzaki, M. Different impacts of saturated and unsaturated free fatty acids on COX-2 expression in C(2)C(12) myotubes. Am. J. Physiol. Endocrinol. Metab. 2009, 297, 129–1303. [Google Scholar] [CrossRef] [Green Version]
- Peng, G.; Li, L.; Liu, Y.; Pu, J.; Zhang, S.; Yu, J.; Zhao, J.; Liu, P. Oleate blocks palmitate-induced abnormal lipid distribution, endoplasmic reticulum expansion and stress, and insulin resistance in skeletal muscle. Endocrinology 2011, 152, 2206–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvado, L.; Coll, T.; Gomez-Foix, A.M.; Salmeron, E.; Barroso, E.; Palomer, X.; Vazquez-Carrera, M. Oleate prevents saturated-fatty-acid-induced ER stress, inflammation and insulin resistance in skeletal muscle cells through an AMPK-dependent mechanism. Diabetologia 2013, 56, 1372–1382. [Google Scholar] [CrossRef] [PubMed]
- Bloch-Damti, A.; Bashan, N. Proposed mechanisms for the induction of insulin resistance by oxidative stress. Antioxid. Redox Signal. 2005, 7, 1553–1567. [Google Scholar] [CrossRef] [PubMed]
- Rachek, L.I.; Musiyenko, S.I.; LeDoux, S.P.; Wilson, G.L. Palmitate induced mitochondrial deoxyribonucleic acid damage and apoptosis in l6 rat skeletal muscle cells. Endocrinology 2007, 148, 293–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambertucci, R.H.; Hirabara, S.M.; Silveira Ldos, R.; Levada-Pires, A.C.; Curi, R.; Pithon-Curi, T.C. Palmitate increases superoxide production through mitochondrial electron transport chain and NADPH oxidase activity in skeletal muscle cells. J. Cell. Physiol. 2008, 216, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Yuzefovych, L.; Wilson, G.; Rachek, L. Different effects of oleate vs. palmitate on mitochondrial function, apoptosis, and insulin signaling in L6 skeletal muscle cells: Role of oxidative stress. Am. J. Physiol. Endocrinol. Metab. 2010, 299, 1096–1105. [Google Scholar] [CrossRef] [Green Version]
- Wen, H.; Gris, D.; Lei, Y.; Jha, S.; Zhang, L.; Huang, M.T.; Brickey, W.J.; Ting, J.P. Fatty acid-induced NLRP3-ASC inflammasome activation interferes with insulin signaling. Nat. Immunol. 2011, 12, 408–415. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Furuhashi, M.; Li, P.; Cao, H.; Tuncman, G.; Sonenberg, N.; Gorgun, C.Z.; Hotamisligil, G.S. Double-stranded RNA-dependent protein kinase links pathogen sensing with stress and metabolic homeostasis. Cell 2010, 140, 338–348. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.J.; Brainard, J.; Song, F.; Wang, X.; Abdelhamid, A.; Hooper, L.; Group, P. Omega-3, omega-6, and total dietary polyunsaturated fat for prevention and treatment of type 2 diabetes mellitus: Systematic review and meta-analysis of randomised controlled trials. BMJ 2019, 366. [Google Scholar] [CrossRef] [Green Version]
- Lourdudoss, C.; Di Giuseppe, D.; Wolk, A.; Westerlind, H.; Klareskog, L.; Alfredsson, L.; van Vollenhoven, R.F.; Lampa, J. Dietary Intake of Polyunsaturated Fatty Acids and Pain in Spite of Inflammatory Control Among Methotrexate-Treated Early Rheumatoid Arthritis Patients. Arthritis Care Res. 2018, 70, 205–212. [Google Scholar] [CrossRef]
- Miles, E.A.; Calder, P.C. Influence of marine n-3 polyunsaturated fatty acids on immune function and a systematic review of their effects on clinical outcomes in rheumatoid arthritis. Br. J. Nutr. 2012, 107, 171–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berbert, A.A.; Kondo, C.R.; Almendra, C.L.; Matsuo, T.; Dichi, I. Supplementation of fish oil and olive oil in patients with rheumatoid arthritis. Nutrition 2005, 21, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Galarraga, B.; Ho, M.; Youssef, H.M.; Hill, A.; McMahon, H.; Hall, C.; Ogston, S.; Nuki, G.; Belch, J.J. Cod liver oil (n-3 fatty acids) as an non-steroidal anti-inflammatory drug sparing agent in rheumatoid arthritis. Rheumatology 2008, 47, 665–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das Gupta, A.B.; Hossain, A.K.; Islam, M.H.; Dey, S.R.; Khan, A.L. Role of omega-3 fatty acid supplementation with indomethacin in suppression of disease activity in rheumatoid arthritis. Bangladesh Med. Res. Counc. Bull. 2009, 35, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, S.; Schmidt, E.B.; Schlemmer, A.; Rasmussen, C.; Johansen, M.B.; Christensen, J.H. Beneficial effect of n-3 polyunsaturated fatty acids on inflammation and analgesic use in psoriatic arthritis: A randomized, double blind, placebo-controlled trial. Scand. J. Rheumatol. 2018, 47, 27–36. [Google Scholar] [CrossRef]
- Sundrarjun, T.; Komindr, S.; Archararit, N.; Dahlan, W.; Puchaiwatananon, O.; Angthararak, S.; Udomsuppayakul, U.; Chuncharunee, S. Effects of n-3 fatty acids on serum interleukin-6, tumour necrosis factor-alpha and soluble tumour necrosis factor receptor p55 in active rheumatoid arthritis. J. Int. Med. Res. 2004, 32, 443–454. [Google Scholar] [CrossRef]
- Remans, P.H.; Sont, J.K.; Wagenaar, L.W.; Wouters-Wesseling, W.; Zuijderduin, W.M.; Jongma, A.; Breedveld, F.C.; Van Laar, J.M. Nutrient supplementation with polyunsaturated fatty acids and micronutrients in rheumatoid arthritis: Clinical and biochemical effects. Eur. J. Clin. Nutr. 2004, 58, 839–845. [Google Scholar] [CrossRef]
- Dawczynski, C.; Schubert, R.; Hein, G.; Muller, A.; Eidner, T.; Vogelsang, H.; Basu, S.; Jahreis, G. Long-term moderate intervention with n-3 long-chain PUFA-supplemented dairy products: Effects on pathophysiological biomarkers in patients with rheumatoid arthritis. Br. J. Nutr. 2009, 101, 1517–1526. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.; Lee, A.; Shim, S.C.; Lee, J.H.; Choe, J.Y.; Ahn, H.; Choi, C.B.; Sung, Y.K.; Bae, S.C. Effect of n-3 polyunsaturated fatty acid supplementation in patients with rheumatoid arthritis: A 16-week randomized, double-blind, placebo-controlled, parallel-design multicenter study in Korea. J. Nutr. Biochem. 2013, 24, 1367–1372. [Google Scholar] [CrossRef]
- Tedeschi, S.K.; Costenbader, K.H. Is There a Role for Diet in the Therapy of Rheumatoid Arthritis? Curr. Rheumatol. Rep. 2016, 18, 23. [Google Scholar] [CrossRef]
- Bryner, R.W.; Woodworth-Hobbs, M.E.; Williamson, D.L.; Alway, S.E. Docosahexaenoic acid protects muscle cells from palmitate-induced atrophy. ISRN Obes. 2012, 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikama, Y.; Ishimaru, N.; Kudo, Y.; Bando, Y.; Aki, N.; Hayashi, Y.; Funaki, M. Effects of free fatty acids on human salivary gland epithelial cells. J. Dent. Res. 2013, 92, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Trotter, A.; Anstadt, E.; Clark, R.B.; Nichols, F.; Dwivedi, A.; Aung, K.; Cervantes, J.L. The role of phospholipase A2 in multiple Sclerosis: A systematic review and meta-analysis. Mult. Scler. Relat. Disord. 2019, 27, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Haase, S.; Haghikia, A.; Gold, R.; Linker, R.A. Dietary fatty acids and susceptibility to multiple sclerosis. Mult. Scler. J. 2018, 24, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Fawcett, P.R.; Shaw, D.A.; Weightman, D. Polyunsaturated fatty acids in treatment of acute remitting multiple sclerosis. Br. Med. J. 1978, 2, 1390–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, J.H.; Zilkha, K.J.; Langman, M.J.; Wright, H.P.; Smith, A.D.; Belin, J.; Thompson, R.H. Double-blind trial of linoleate supplementation of the diet in multiple sclerosis. Br. Med. J. 1973, 1, 765–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paty, D.W.; Cousin, H.K.; Read, S.; Adlakha, K. Linoleic acid in multiple sclerosis: Failure to show any therapeutic benefit. Acta Neurol. Scand. 1978, 58, 53–58. [Google Scholar] [CrossRef]
- Bates, D.; Cartlidge, N.E.; French, J.M.; Jackson, M.J.; Nightingale, S.; Shaw, D.A.; Smith, S.; Woo, E.; Hawkins, S.A.; Millar, J.H.; et al. A double-blind controlled trial of long chain n-3 polyunsaturated fatty acids in the treatment of multiple sclerosis. J. Neurol. Neurosurg. Psychiatry 1989, 52, 18–22. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.M.; Willett, W.C.; Hernan, M.A.; Olek, M.J.; Ascherio, A. Dietary fat in relation to risk of multiple sclerosis among two large cohorts of women. Am. J. Epidemiol. 2000, 152, 1056–1064. [Google Scholar] [CrossRef] [Green Version]
- Nauta, A.J.; Ben Amor, K.; Knol, J.; Garssen, J.; van der Beek, E.M. Relevance of pre- and postnatal nutrition to development and interplay between the microbiota and metabolic and immune systems. Am. J. Clin. Nutr. 2013, 98, 586–593. [Google Scholar] [CrossRef]
- Dullaart, R.P.; Beusekamp, B.J.; Meijer, S.; Hoogenberg, K.; van Doormaal, J.J.; Sluiter, W.J. Long-term effects of linoleic-acid-enriched diet on albuminuria and lipid levels in type 1 (insulin-dependent) diabetic patients with elevated urinary albumin excretion. Diabetologia 1992, 35, 165–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Diabetes, A. Standards of Medical Care in Diabetes-2017 Abridged for Primary Care Providers. Clin. Diabetes 2017, 35, 5–26. [Google Scholar] [CrossRef] [Green Version]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Pickup, J.C. Inflammation and activated innate immunity in the pathogenesis of type 2 diabetes. Diabetes Care 2004, 27, 813–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oguntibeju, O.O. Type 2 diabetes mellitus, oxidative stress and inflammation: Examining the links. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 45–63. [Google Scholar]
- Sriwijitkamol, A.; Christ-Roberts, C.; Berria, R.; Eagan, P.; Pratipanawatr, T.; DeFronzo, R.A.; Mandarino, L.J.; Musi, N. Reduced skeletal muscle inhibitor of kappaB beta content is associated with insulin resistance in subjects with type 2 diabetes: Reversal by exercise training. Diabetes 2006, 55, 760–767. [Google Scholar] [CrossRef] [Green Version]
- Senn, J.J. Toll-like receptor-2 is essential for the development of palmitate-induced insulin resistance in myotubes. J. Biol. Chem. 2006, 281, 26865–26875. [Google Scholar] [CrossRef] [Green Version]
- Clore, J.N.; Allred, J.; White, D.; Li, J.; Stillman, J. The role of plasma fatty acid composition in endogenous glucose production in patients with type 2 diabetes mellitus. Metabolism 2002, 51, 1471–1477. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, D.; Gentile, C.L.; Pagliassotti, M.J. Reduced endoplasmic reticulum luminal calcium links saturated fatty acid-mediated endoplasmic reticulum stress and cell death in liver cells. Mol. Cell. Biochem. 2009, 331, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Diakogiannaki, E.; Morgan, N.G. Differential regulation of the ER stress response by long-chain fatty acids in the pancreatic beta-cell. Biochem. Soc. Trans. 2008, 36, 959–962. [Google Scholar] [CrossRef] [Green Version]
- Vessby, B.; Uusitupa, M.; Hermansen, K.; Riccardi, G.; Rivellese, A.A.; Tapsell, L.C.; Nalsen, C.; Berglund, L.; Louheranta, A.; Rasmussen, B.M.; et al. Substituting dietary saturated for monounsaturated fat impairs insulin sensitivity in healthy men and women: The KANWU Study. Diabetologia 2001, 44, 312–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez-Gomez, Y.; Cruz-Teno, C.; Rangel-Zuniga, O.A.; Peinado, J.R.; Perez-Martinez, P.; Delgado-Lista, J.; Garcia-Rios, A.; Camargo, A.; Vazquez-Martinez, R.; Ortega-Bellido, M.; et al. Effect of dietary fat modification on subcutaneous white adipose tissue insulin sensitivity in patients with metabolic syndrome. Mol. Nutr. Food Res. 2014, 58, 2177–2188. [Google Scholar] [CrossRef] [PubMed]
- Bonnard, C.; Durand, A.; Peyrol, S.; Chanseaume, E.; Chauvin, M.A.; Morio, B.; Vidal, H.; Rieusset, J. Mitochondrial dysfunction results from oxidative stress in the skeletal muscle of diet-induced insulin-resistant mice. J. Clin. Investig. 2008, 118, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.J.; Lustig, M.E.; Boyle, K.E.; Woodlief, T.L.; Kane, D.A.; Lin, C.T.; Price, J.W., 3rd; Kang, L.; Rabinovitch, P.S.; Szeto, H.H.; et al. Mitochondrial H2O2 emission and cellular redox state link excess fat intake to insulin resistance in both rodents and humans. J. Clin. Investig. 2009, 119, 573–581. [Google Scholar] [CrossRef]
- Kelley, D.E.; He, J.; Menshikova, E.V.; Ritov, V.B. Dysfunction of mitochondria in human skeletal muscle in type 2 diabetes. Diabetes 2002, 51, 2944–2950. [Google Scholar] [CrossRef] [Green Version]
- Petersen, K.F.; Dufour, S.; Befroy, D.; Garcia, R.; Shulman, G.I. Impaired mitochondrial activity in the insulin-resistant offspring of patients with type 2 diabetes. N. Engl. J. Med. 2004, 350, 664–671. [Google Scholar] [CrossRef] [Green Version]
- Di Meo, S.; Iossa, S.; Venditti, P. Skeletal muscle insulin resistance: Role of mitochondria and other ROS sources. J. Endocrinol. 2017, 233, 15–42. [Google Scholar] [CrossRef] [Green Version]
- Finucane, O.M.; Lyons, C.L.; Murphy, A.M.; Reynolds, C.M.; Klinger, R.; Healy, N.P.; Cooke, A.A.; Coll, R.C.; McAllan, L.; Nilaweera, K.N.; et al. Monounsaturated fatty acid-enriched high-fat diets impede adipose NLRP3 inflammasome-mediated IL-1beta secretion and insulin resistance despite obesity. Diabetes 2015, 64, 2116–2128. [Google Scholar] [CrossRef] [Green Version]
- Poreba, M.; Mostowik, M.; Siniarski, A.; Golebiowska-Wiatrak, R.; Malinowski, K.P.; Haberka, M.; Konduracka, E.; Nessler, J.; Undas, A.; Gajos, G. Treatment with high-dose n-3 PUFAs has no effect on platelet function, coagulation, metabolic status or inflammation in patients with atherosclerosis and type 2 diabetes. Cardiovasc. Diabetol. 2017, 16, 50. [Google Scholar] [CrossRef] [Green Version]
- O’Mahoney, L.L.; Matu, J.; Price, O.J.; Birch, K.M.; Ajjan, R.A.; Farrar, D.; Tapp, R.; West, D.J.; Deighton, K.; Campbell, M.D. Omega-3 polyunsaturated fatty acids favourably modulate cardiometabolic biomarkers in type 2 diabetes: A meta-analysis and meta-regression of randomized controlled trials. Cardiovasc. Diabetol. 2018, 17, 98. [Google Scholar] [CrossRef]







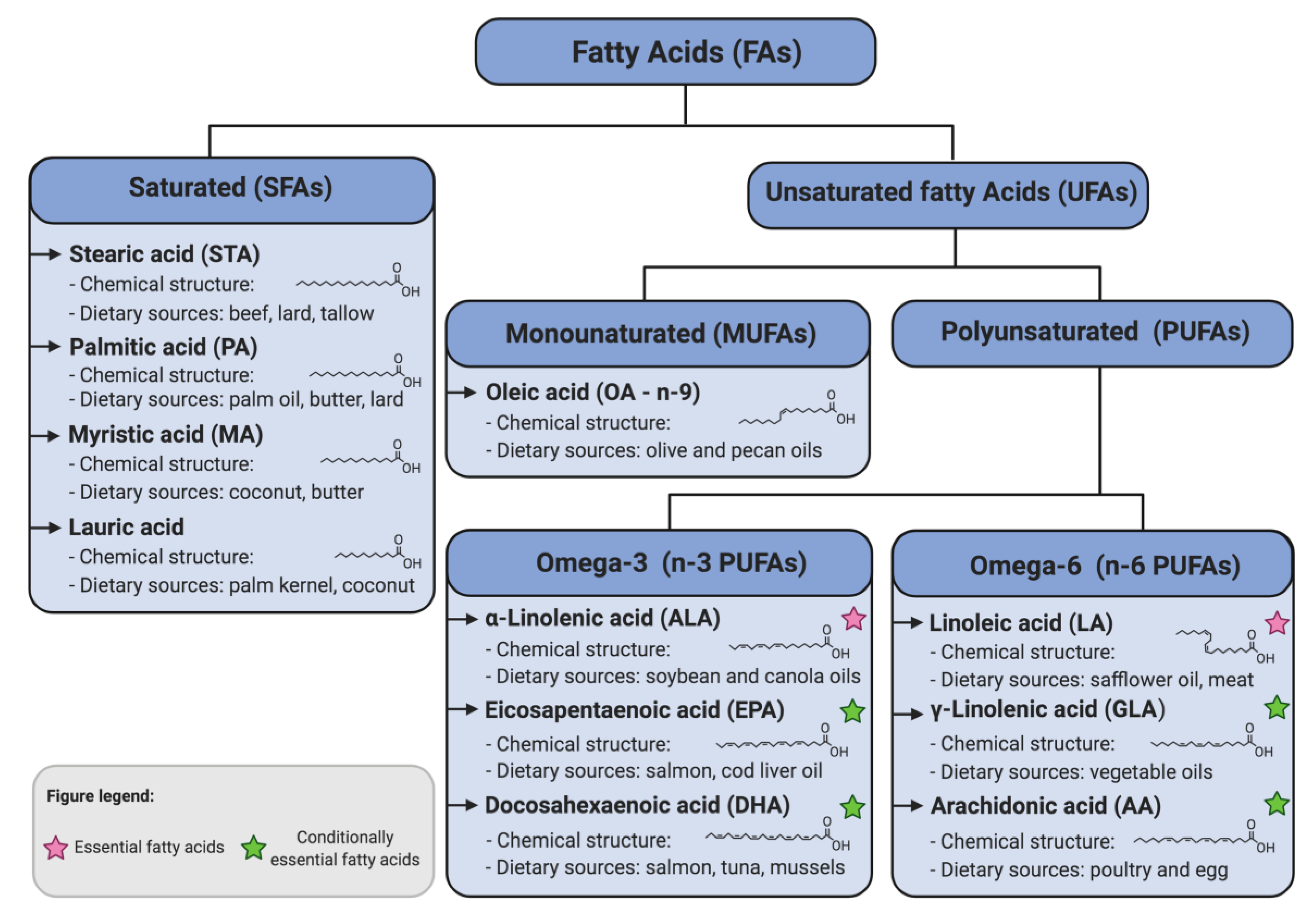{kind=link}
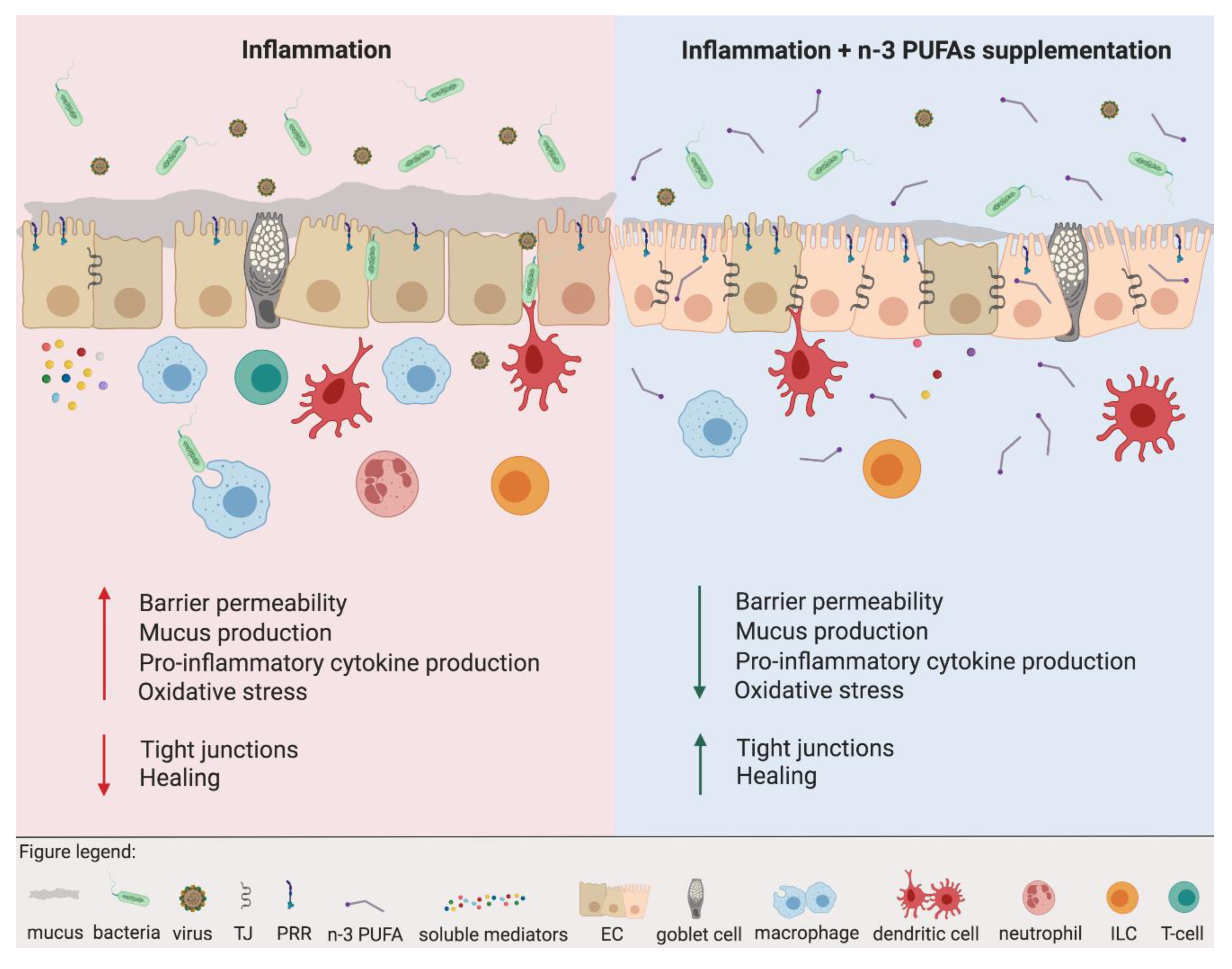{kind=link}
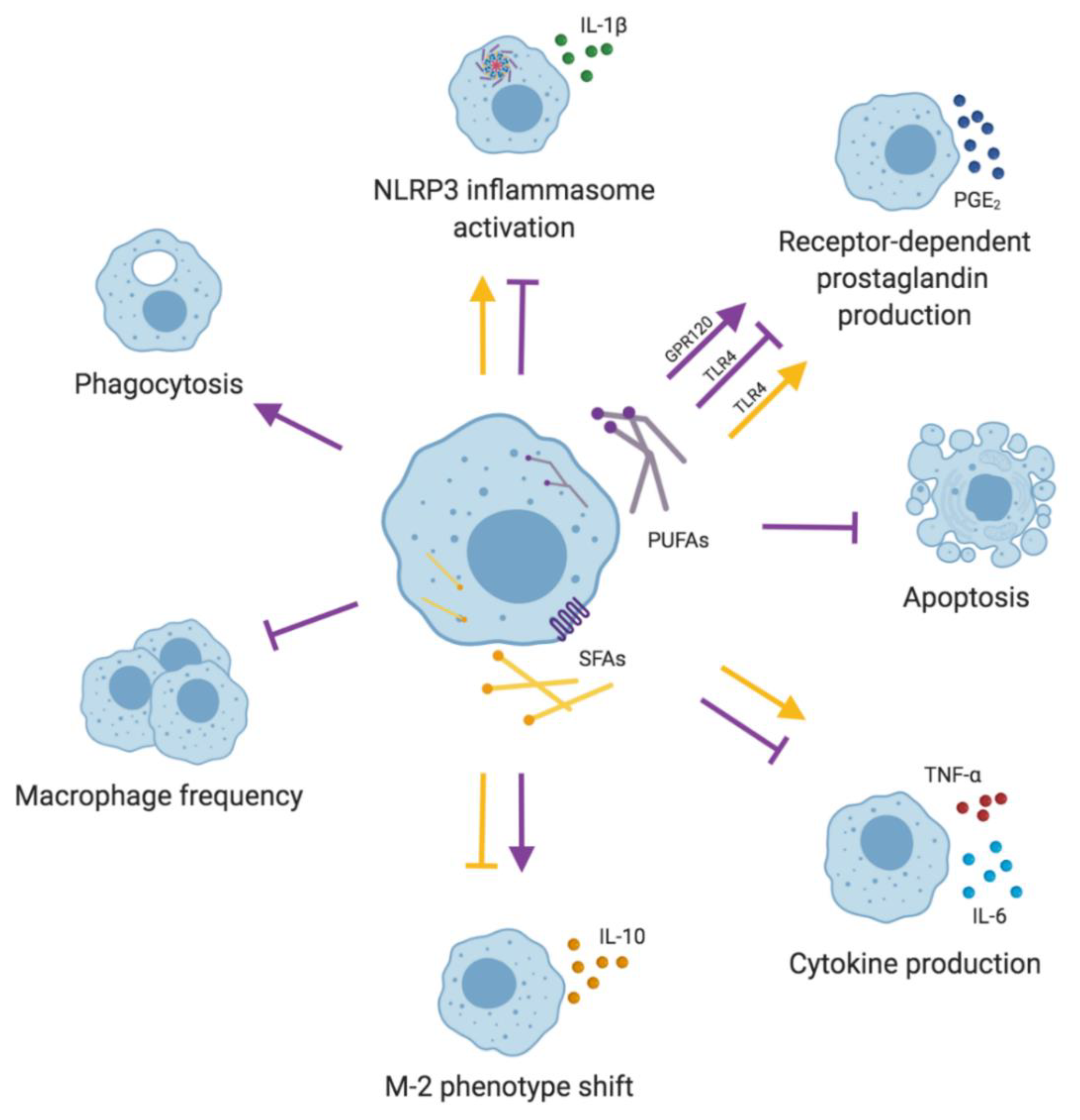{kind=link}
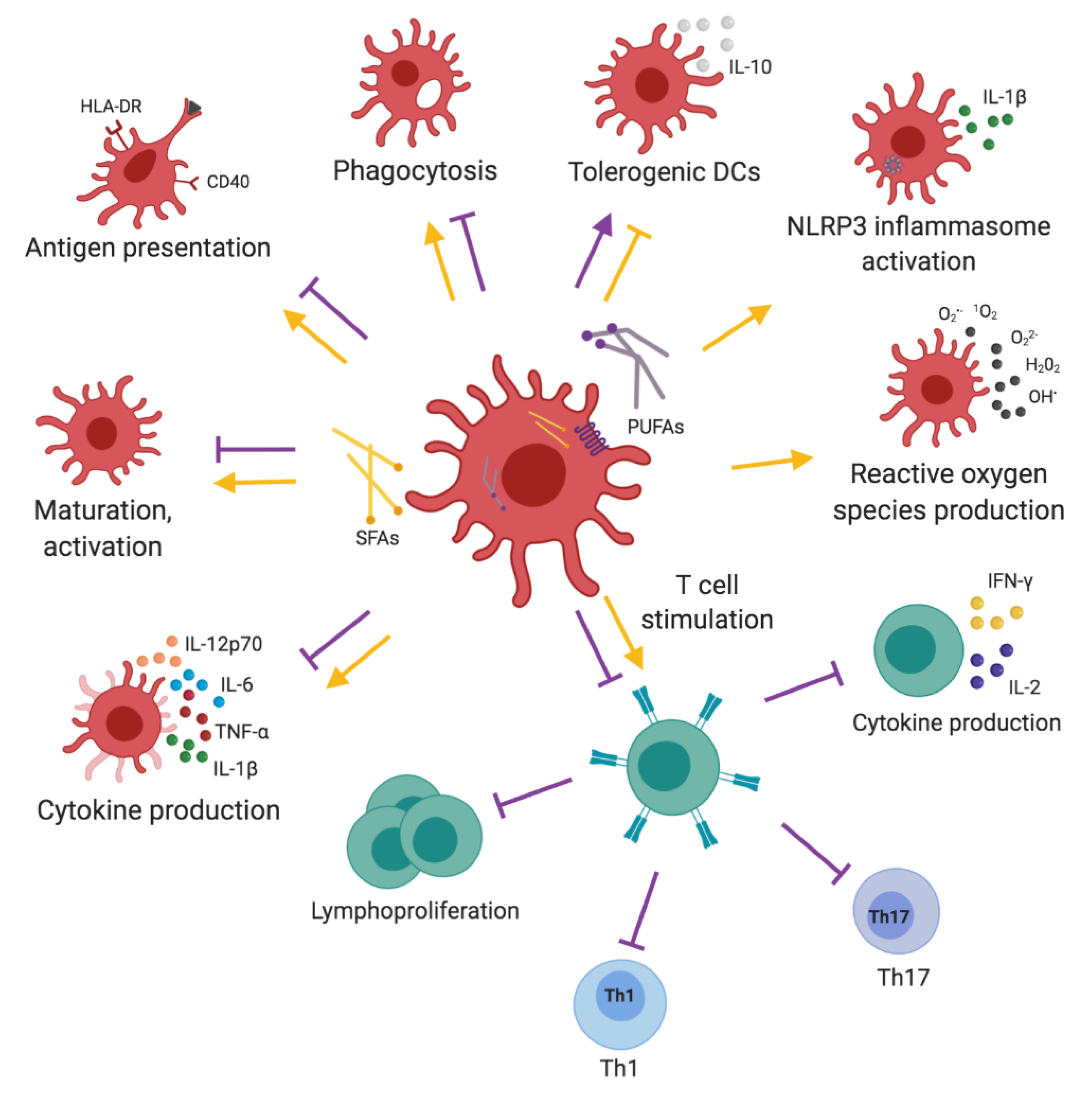{kind=link}
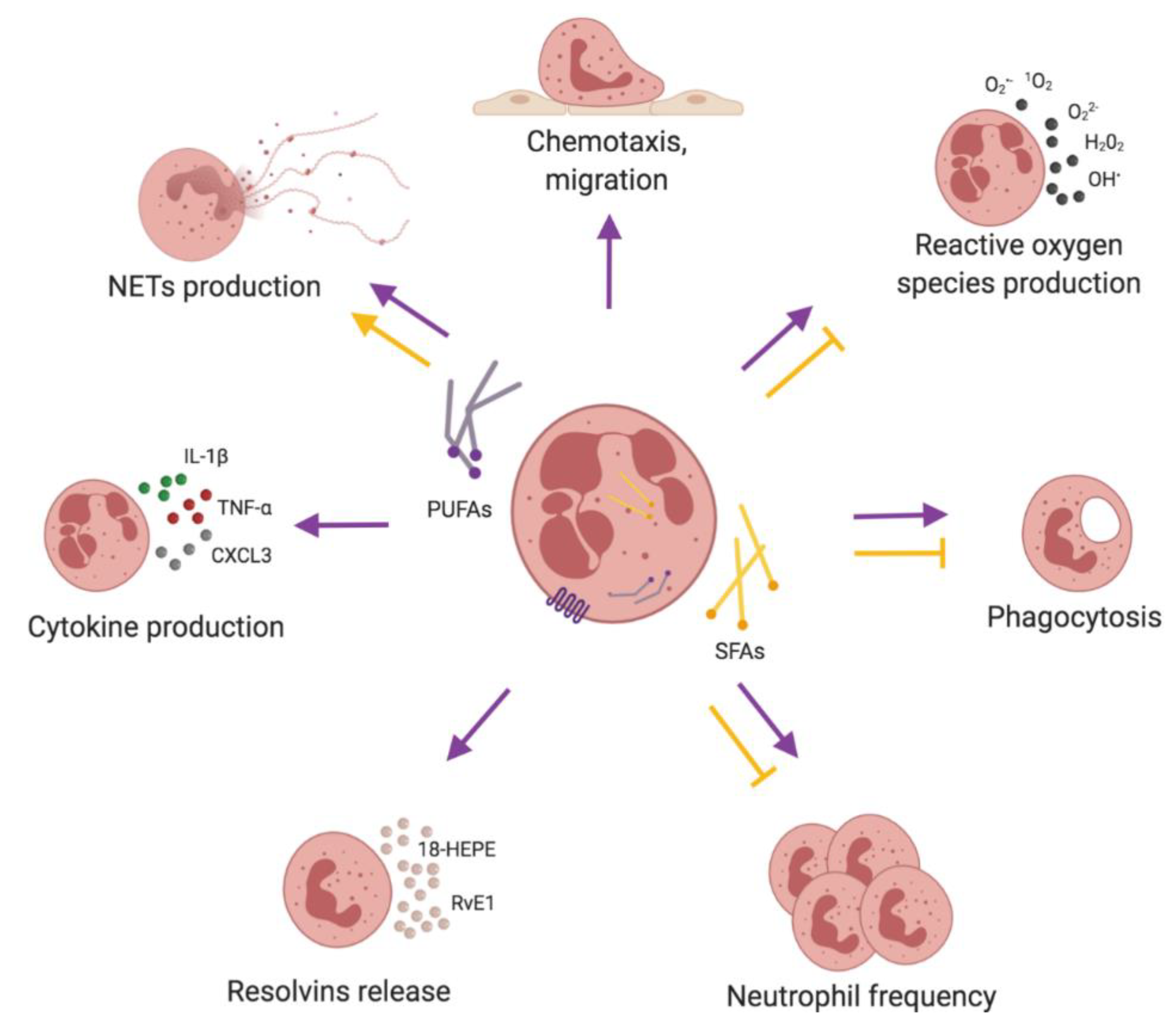{kind=link}
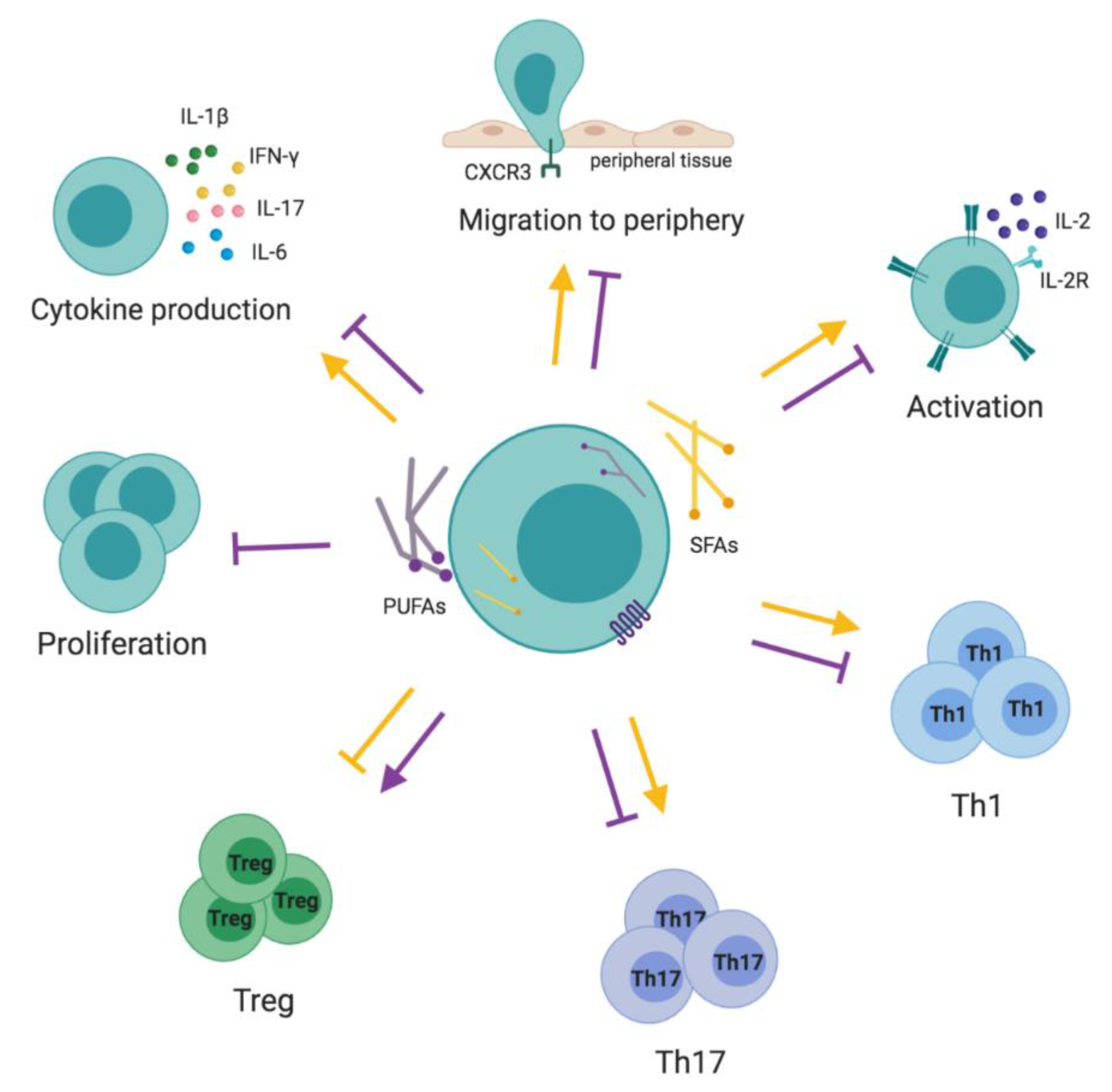{kind=link}
| FAs | Cell Type | In Vitro Effect | Ref. |
|---|---|---|---|
| EPA | Epithelium | ↓ Membrane permeability, ↑ TEER and restored TJ distribution in IECs | [31] |
| ↓ IL-6 and/or IL-8 production mediated by PPARγ in IECs | [34] | ||
| ↑ GPR120-dependent calcium accumulation, ↑ MAPK-ERK1/2 pathways and ↓ IL-1β-induced NF-κB activation in IECs | [36] | ||
| Macrophages | ↓ TLR4-dependent and LPS-mediated COX-2 activation and subsequent prostaglandin release | [53,54] | |
| Dendritic cells | ↓ Activated CD1a+ DC frequency and ↓ GPR120 receptor expression | [55] | |
| ↓ MHC class II (HLA-DR) and ↓ costimulatory molecule CD40, CD80, CD83, CD86 expression | [56,57,58] | ||
| ↓ Expression of IL-12p40 and TNF-α | [58] | ||
| ↓ T cell activation and ↓ IL-2 and IFN-γ production by DC-EPA-stimulated T cells | [58] | ||
| ↓ Responses to LPS, independent from NF-κB and eicosanoid system | [58] | ||
| ↓ Capacity to stimulate T cell proliferation | [34] | ||
| ↓TNF-α and IL-10 production in DCs, ↓ IFN-γ and ↑ IL-13 secretion in mixed DC-lymphocyte reaction | [34] | ||
| Neutrophils | ↑ ROS production and TNF-α production | [59] | |
| ↓ Adhesion due to ↑ oxidized-EPA-dependent activation of PPARα | [60] | ||
| T cells | ↓ IL-2, IL-2R, TNF-α, IL-4, and IL-10 expression | [61,62,63,64,65,66] | |
| ↓ JAK1 and JAK3 phosphorylation with the subsequent inhibition of STAT5 phosphorylation | [67] | ||
| ↓ Akt and ERK1/2 phosphorylation induced by IL-2 | [67,68] | ||
| ↑ ROS-dependent cell death | [63] | ||
| B cells | ↓ IL-10, TNF-α and INF-γ production | [69] | |
| DHA | Epithelium | ↓ Membrane permeability, ↑ TEER and restored TJ distribution in IECs | [31] |
| ↑ Barrier functions in the presence of inflammatory conditions (IL-1β, TNF-α, IFN-γ and LPS) in IECs | [33] | ||
| ↓ IL-6 and/or IL-8 production mediated by PPARγ in IECs | [34] | ||
| ↑ GPR120-dependent calcium accumulation, ↑ MAPK-ERK1/2 pathways and ↓ IL-1β-induced NF-κB activation in IECs | [36] | ||
| ↓ IL-6 and IP-10 secretion after rhinovirus infection (AECs) | [41] | ||
| ↑ Permeability and mucus production and ↓ TEER in AECs | [43] | ||
| ↑ Healing capacity after dust extract stimulation in AECs | [42] | ||
| Macrophages | ↑ M-2 polarization regulated by p38 MAPK and mediated via PPARγ | [70,71] | |
| ↑ GPR120-mediated M2-like polarization, ↑ phagocytic activity and ↑ IL-10 secretion | [72,73] | ||
| ↑ GRP120-dependent activation of cPLA2, ↑ release of AA, COX-2 activation and PGE2 release and ↑ anti-inflammatory PGE2-mediated signaling on EP4 | [74] | ||
| ↓ TLR4-dependent and LPS-mediated COX-2 activation and subsequent prostaglandin release | [53,54] | ||
| Dendritic cells | ↓ Activated CD1a+ DC frequency and ↓ GPR120 receptor expression | [55] | |
| ↓ MHC class II (HLA-DR) and ↓ costimulatory molecule CD40, CD80, CD83, CD86 expression | [56,57,58] | ||
| ↓ Maturation, expression of IL-12p70, IL-6, IL-23 production | [75] | ||
| TLR2, 3, 4 and 9-dependent ↓ of IL-12p70, IL-23 and IL-27 expression mediated by PPARγ activation and ↓NF-κBp65 | [56] | ||
| ↑ CD36, HLA-DR, CD83, CD86 and ↓ CD1a expression | [76] | ||
| ↓ IL-12p70 and IL-6; ↓ IL-10 expression dependent of PPARγ:RXR heterodimer | [76] | ||
| ↓TNF-α and IL-10 production in DCs, ↓ IFN-γ and ↑ IL-13 secretion in mixed DC-lymphocyte reaction | [34] | ||
| ↓ Capacity to stimulate proliferation of antigen-specific T cells and their Th1/Th17 differentiation | [34,75] | ||
| ↑ Expression of p27Kip1, and ↓ T-bet, GATA3, and RORγT in DC-DHA activated T cells | [75] | ||
| ↑ TGF-β and FoxP3 expression in DC-DHA activated T cells | [75] | ||
| Neutrophils | ↑ ROS production, ↑ IL-1β, and TNF-α production and ↑ phagocytosis and antifungal responses | [59] | |
| T cells | ↓ IL-2, IL-2R, TNF-α, IL-4, and IL-10 expression | [61,62,63,64,65,66] | |
| ↓ JAK1 and JAK3 phosphorylation with the subsequent ↓ of STAT5 phosphorylation | [67] | ||
| ↓ Akt and ERK1/2 phosphorylation induced by IL-2 | [67,68] | ||
| ↓ T-cell proliferation in DCs and T cell co-culture by ↑ FoxP3+ T cell population | [77] | ||
| Modulation of calcium concentration, recruiting calcium from the intracellular pool and opening calcium release-activated calcium channels | [78,79,80,81,82] | ||
| B cells | ↓ TLR4 expression, probably due to altering the membrane FA composition and the formation of the lipid rafts | [83] | |
| ↓ IL-10, TNF-α, INF-γ and IL-6 expression | [69] | ||
| ALA | Epithelium | ↓ IL-6 and/or IL-8 secretion (IECs) | [34] |
| Dendritic cells | ↓ Activated CD1a+ DC frequency and ↓ GPR120 receptor expression | [55] | |
| LA | Epithelium | ↑ Permeability and mucus production and ↓ TEER in AECs | [43] |
| Dendritic cells | ↑ IL-10, IL-10R and ↓ IL-12 expression, ↑ activation of ERK and NF-κB | [84] | |
| Neutrophils | ↓ LPS-induced IL-1β secretion, mediated by ↓ TLR2, TLR4, NFKBIA, P2RX7, NLRP3, CASP-1 expression and ↑ activation of PPAR-γ after A1AT-LA stimulation | [85] | |
| ↑ ROS production dependent from calcium signaling | [86] | ||
| T cells | ↑ ROS-induced apoptosis | [87] | |
| Other n-6 PUFAs | Epithelium | ↑ Intestinal barrier integrity after IL-4-induced inflammation (DGLA, AA) in IECs | [32] |
| ↓ IL-6 and/or IL-8 production mediated by PPARγ in IECs | [34] | ||
| ↑ GPR120-dependent calcium accumulation, ↑ MAPK-ERK1/2 pathways and ↓ IL-1β-induced NF-κB activation (AA) in IECs | [36] | ||
| ↑ Permeability and mucus production, ↓ TEER (AA, GLA) in AECs | [43] | ||
| ↑ Cytotoxicity and ↑ IL-6 secretion (AA) in AECs | [43,44] | ||
| ↑ IL-6 and IL-8 expression dependent on JNK and p38 MAPK signaling after AA stimulation combined with rhinovirus infection in AECs | [48] | ||
| Dendritic cells | ↓ Expression of IL-12p40 and TNF-α (AA) | [58] | |
| ↓ T cell activation and ↓ IL-2 and IFN-γ production by EPA-DC stimulated T cells (AA) | [58] | ||
| ↓ Responses to LPS, independent from NF-κB and eicosanoid system | [58] | ||
| Neutrophils | ↑ TNFR1 and TNFR2 receptor expression mediated via PKC, ERK1/2 and cPLA2 (AA) | [88] | |
| ↓ ATP production and ↑ LDH release | [89] | ||
| ↑ Mitochondria depolarization and lipid accumulation | [90] | ||
| ↑ ROS production dependent from calcium signaling (AA) | [86] | ||
| T cells | ↓ T-cell proliferation in DCs and T cell co-culture by ↑ FoxP3+ T cell population (AA) | [77] | |
| OA | Neutrophils | ↑ ROS production, phagocytosis and killing capacities, ↑ VEGF, CXCL3 and IL-1β production | [91,92,93] |
| ↑ NETosis | [94] | ||
| T cells | ↑ Apoptosis via caspase-3 pathway (high doses) | [87] | |
| PA | Macrophages | ↑ Activation of TLR4 and NF-κB pathway | [95] |
| ↑ IL-18 and TNF-α secretion | [96] | ||
| ↑ Activation of TLR2 and NLRP3 inflammasome and ↑ of IL-1β-secretion | [97] | ||
| Dendritic cells | ↑ TLR4 endocytosis, ↑ CD86, and CD83 expression and ↑ TLR4-dependent secretion of IL-1β via NF-κB canonical pathway | [98] | |
| Dendritic cells | ↑ ROS production | [98] | |
| ↑ Maturation and ↑ activation | [98] | ||
| Neutrophils | ↑ NETosis | [94] | |
| T cells | ↑ Activation of PI3K/Akt signaling | [99,100,101] | |
| ↑ SLAM3 expression dependent from JAK/STAT5, ↑ of TNF-α, IL-1β, IL-6, IFN-γ, IL17A and IL-2 expression | [100,102] | ||
| ↑ CD69 and ↓ CCR7 and L-selectin expression | [99] | ||
| B cells | ↓ B-cell activation (through lipoapoptosis) | [103] | |
| Lauric acid | Dendritic cells | ↑ CD40, CD80, CD86 and MHC class II expression | [104] |
| ↑ IL-12p70 and IL-6 secretion | [104] | ||
| ↑ TLR4 activity and T cell activation capacity | [104] | ||
| T cells | ↑ IL-2, IL-17 and GM-CS production | [105] | |
| ↑ Th17 differentiation | [105] | ||
| ↓ Th2 differentiation | [105] | ||
| B cells | ↑ TLR4 dimerization and ↑ recruitment to lipid rafts on the plasma membrane | [83] |
| Diet | Cell Type | In Vivo Effect | Ref. |
|---|---|---|---|
| n-3 PUFA-rich diet | Epithelium | ↑ Barrier function in the mouse model of intestinal anaphylactic response | [37] |
| ↓ Inflammatory score, ↑ TJ proteins expression (ZO-1 and Occludin), ↓ IL-17, TNF-α and IFN-γ production in the IL-10-deficient model of chronic colitis | [38] | ||
| ↓ Intestinal pathology scores and IL-12, TNF-α, IL-1β secretion and ↑ of ZO-1 expression in SCID mice in the model of colitis | [39] | ||
| ↓ Oxidative stress (8-IP, glutathione and iNOS) in a rat model of colitis | [40] | ||
| ↑ Healing capacity during dust-stimulated lung inflammation | [42] | ||
| Macrophages | ↓ Frequency of ATMs and ↑ M2 anti-inflammatory phenotype and ↑ expression of IL-10, arginase, YM-1, Clec7a and MMR | [72] | |
| ↑ Phagocytosis, microbicidal activity and ↓ apoptosis | [106,107] | ||
| ↓ NLRP3 inflammasome activation via GPR120 and GPR40 receptors and their downstream protein β-arrestin-2 | [108,109] | ||
| ↑ Autophagy | [110] | ||
| Dendritic cells | ↓ Capacity to stimulate proliferation of antigen-specific T cells and their Th1/Th17 differentiation | [75] | |
| ↓ Antigen-presenting properties and ↓ CD2+ DCs, and ↓ CD18, CD11a, HLA-DR, CD54 expression | [111] | ||
| ↓ CD80 and CD11c+ expression, ↓ TNF-α production and ↓ phagocytosis | [112] | ||
| ↓ Th1-inducing, pro-inflammatory (CD11c+ CD11b− CD8α+) lymphoid DCs and ↑ myeloid, tolerogenic (CD11c+ CD11b+ CD8α−) DCs subpopulations | [112,113] | ||
| ↓ T cell proliferation and ↓ IFN-γ and IL-17 production in the DC-lymphocyte reaction | [114] | ||
| Neutrophils | Protection during S. aureus sepsis, with ↓ mortality and bacteria load | [115] | |
| ↑ CD11bhigh, Ly6Ghigh and MHC class IIhigh neutrophil subpopulation in the blood | [116] | ||
| ↑ Specialized pro-resolving mediators, such as 18-HEPE, RvE1, RvE2, RvE3, 17-HDHA and RvE5 | [117] | ||
| ↑ Neutrophil telomere length, probably due to ↓ oxidative stress | [118] | ||
| ↑ Membrane integrity, ↓ ROS production, and mitochondrial membrane potential after exercise | [119] | ||
| ↑ Functions and frequency in oncologic patients undergoing chemotherapy | [120] | ||
| ↑ Neutrophil-dependent inflammation in mice genetically susceptible to colitis | [121] | ||
| T cells | ↓ Proliferation via ↓ IL-2 secretion and IL-2RA expression | [122] | |
| ↓ Production of diacylglycerol, ceramide, and level of phospholipase Cγ | [122] | ||
| Disrupted spatial organization of the second messenger, PI(4,5)P2, perturbing downstream signals required for T cell proliferation | [123] | ||
| ↓ Frequency of pro-inflammatory T cells in the fat tissues, mediated by CCR-4, CXCR4 and ↓ expression of P-selectin and ICAM-1 on the endothelium | [66,124] | ||
| ↓ Formation of pseudopods and ↓ ratio between F-actin and G-actin and ↓ Rhoα and Rac1 involved in cell migration | [125] | ||
| ↓ IL-6, IL-23, IL-17 expression and ↑ FoxP3, CTLA-4, TGF-β and IL-10 expression | [66,126,127] | ||
| ↓ Treg-dependent proliferation of T effector cells mediated by ↓ of ERK1/2 and Akt phosphorylation and ↑ histone deacetylase and p27Kip1 expression | [66] | ||
| B cells | ↑ CD69 and CD40 expression | [103,128] | |
| ↑ IL-6, IFN-γ, TNF-α and IL-10, IL-5, IL-13, and IL-9 expression | [103,128,129] | ||
| ↑ Percentage of splenic transitional, marginal zone B cells and peritoneal B1 cells | [130,131] | ||
| ↑ Surface expression of IgM in spleen and serum | [130,131] | ||
| ↑ Caecal IgA | [128] | ||
| Changed lipid composition of B cells and ↑ size, order and the distribution of rafts | [128,129,132] | ||
| SFA-rich diet | Dendritic cells | ↓ Tolerogenic (CD11c+ CD103+ CD11b+) and ↑ pro-inflammatory (CD11c+ CD103− CD11b+) subpopulations in the gut | [133] |
| ↑ TLR4-dependent NLRP3 inflammasome activation and IL-1β secretion | [134] | ||
| Neutrophils | ↓ Survival rate and ↑ bacterial proliferation in septic mice | [135] | |
| ↓ Cell frequency, ↓ phagocytosis and ↓ ROS production in septic mice | [135] | ||
| T cells | ↑ CXCR3 and ↓ CCR7 and L-selectin expression | [99] | |
| ↑ Severity of the disease, ↑ Th1, and Th17 cell differentiation and ↑ T cell infiltration into the central nervous system | [101,136] | ||
| Positive association with several markers of inflammation, such as C-reactive protein, IL-1RA, IFN-γ and CCL5 | [137] |
| Allergic Diseases | Effects of n-3 PUFAs | Ref. | |||
| Asthma | Pregnancy and lactation | Prevention of asthma and allergic disease development in the offspring | [136,196,214,216,217] | ||
| ↑ Risk of allergic diseases and ↓ lung function in the offspring associated with high n-3 PUFAs in the colostrum | [249] | ||||
| Infancy and childhood | ↓ Risk of asthma, rhinitis and aeroallergen sensitization | [200,204,220,221] | |||
| ↓ Asthma symptoms scores and ↓ responsiveness to acetylcholine | [222] | ||||
| Improvement in pulmonary functions and ↓ use of short-acting inhaled bronchodilators and inhaled corticosteroids | [290] | ||||
| ↑ Prevalence of wheeze | [248] | ||||
| Adulthood | ↓ Prevalence of asthma symptoms, airway hyperresponsiveness and ↑ asthma control and lung function | [47,202,206,207,224,226,227,228,230,233] | |||
| ↓ Exhaled nitric oxide, ↓ serum eosinophils and ↑ FEV1 | [225,229] | ||||
| ↓ Bronchodilator use, ↓ exhale nitric oxide and ↓ severity of exercise-induced bronchoconstriction | [228,231,232,233] | ||||
| Allergic Rhinitis | Infancy and childhood | ↓ Risk of rhinitis development | [251,254,255,256] | ||
| Adulthood | ↓ Prevalence | [257,258] | |||
| Atopic Dermatitis | Pregnancy and lactation | ↓ Incidence and intensity in the offspring | [259,260] | ||
| Infancy and childhood | ↓ Frequency of eczema | [261] | |||
| Adulthood | ↓ Severity and symptoms | [271,272,275] | |||
| ↑ SCORing Atopic Dermatitis (SCORAD) after 4, 8 and 16 weeks of treatment | [273] | ||||
| Food Allergy | Pregnancy and lactation | ↓ Allergic sensitization to food proteins (antigen-dependent effect) | [196,260,277,278,279,280,289] | ||
| Infancy and childhood | ↓ Odds ratio for food allergy and ↓ risk for sensitization (antigen-dependent effect) | [259,277,281,282,287] | |||
| Autoimmune Diseases | Effects of n-3 PUFAs | Ref. | |||
| Rheumatoid Arthritis | Positive correlation with synovial fluid composition and inversed correlation with pain score | [291] | |||
| ↓ Incidence and improved prognosis | [292,293,294,295,296] | ||||
| ↓ Duration of morning stiffness and ↓ number of tender and swollen joints | [297] | ||||
| Polymyositis and Dermatomyositis | ↑ Skeletal muscle growth and functions | [298] | |||
| Sjögren Syndrome | ↓ Inflammation and ↑ barrier function in the salivary glands | [299,300] | |||
| Systemic Lupus Erythematosus | ↓ Inflammatory mediators, leukocyte chemotaxis and adhesion molecules expression | [301] | |||
| Antiphospholipid Syndrome | ↑ Endothelial function | [302] | |||
| Multiple Sclerosis | ↑ M2-like macrophages shift, prevented de-myelinization, promoted neuroprotection and re-myelinization ↓ incidence | [303,304,305,306] | |||
| Type 1 Diabetes | Protection against T1DM in infants, no protective effect in older children reported | [307,308] | |||
| Metabolic Diseases | Effects of FAs | Ref. | |||
| Type 2 Diabetes | SFAs | ↑ Intramyocellular accumulation of DAG and/or ceramide, ↑ PKCθ activation and ↑ NF-κB-dependent expression of IL-6 and TNF-α | [309,310] | ||
| ↑ TLR-independent PGE2 production via COX-2 and p38 MAPK pathways | [311] | ||||
| ↓ AMPK activation, with subsequent ↑ ER stress associated with inflammation and insulin resistance | [312,313] | ||||
| ↑ ROS production in muscle cells and muscle insulin resistance development | [314,315,316,317] | ||||
| ↑ ROS production and ROS-dependent ↑ NLRP3-inflammasome activation and insulin resistance | [318] | ||||
| ↑ PKR activation and induction of JNK inflammatory pathways and inhibition of insulin signaling | [319] | ||||
| UFAs | ↓ PA-dependent COX-2 expression and related PGE2 production (OA, LA) | [311] | |||
| ↓ PA-dependent ER stress associated with reduced inflammation and insulin resistance (OA) | [313] | ||||
| ↓ PA-dependent muscle destruction and insulin resistance development (OA) | [317] | ||||
| ↓ NLRP3-inflammasome-dependent IL-1β secretion in adipose tissue and insulin resistance (EPA, DHA) | [108] | ||||
| Limited or no effect of PUFA supplementation on T2DM prevention and treatment | [320] | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radzikowska, U.; Rinaldi, A.O.; Çelebi Sözener, Z.; Karaguzel, D.; Wojcik, M.; Cypryk, K.; Akdis, M.; Akdis, C.A.; Sokolowska, M. The Influence of Dietary Fatty Acids on Immune Responses. Nutrients 2019, 11, 2990. https://doi.org/10.3390/nu11122990
Radzikowska U, Rinaldi AO, Çelebi Sözener Z, Karaguzel D, Wojcik M, Cypryk K, Akdis M, Akdis CA, Sokolowska M. The Influence of Dietary Fatty Acids on Immune Responses. Nutrients. 2019; 11(12):2990. https://doi.org/10.3390/nu11122990
Chicago/Turabian StyleRadzikowska, Urszula, Arturo O. Rinaldi, Zeynep Çelebi Sözener, Dilara Karaguzel, Marzena Wojcik, Katarzyna Cypryk, Mübeccel Akdis, Cezmi A. Akdis, and Milena Sokolowska. 2019. "The Influence of Dietary Fatty Acids on Immune Responses" Nutrients 11, no. 12: 2990. https://doi.org/10.3390/nu11122990
APA StyleRadzikowska, U., Rinaldi, A. O., Çelebi Sözener, Z., Karaguzel, D., Wojcik, M., Cypryk, K., Akdis, M., Akdis, C. A., & Sokolowska, M. (2019). The Influence of Dietary Fatty Acids on Immune Responses. Nutrients, 11(12), 2990. https://doi.org/10.3390/nu11122990