Biotechnological Approaches for Generating Zinc-Enriched Crops to Combat Malnutrition


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods of Biofortification
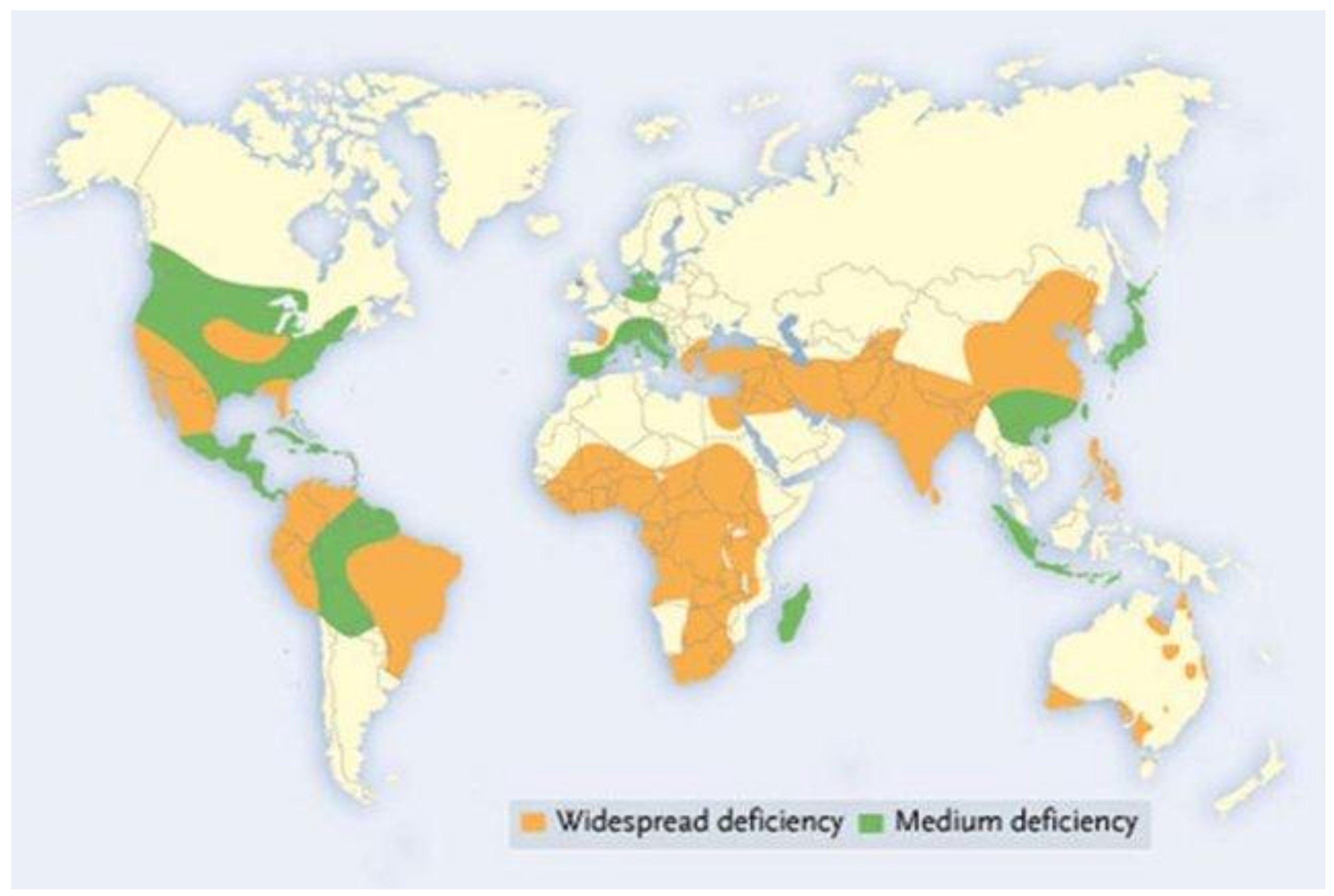
2.1. Zinc and Human Biology
2.1.1. Importance of Zinc in Human Health
2.1.2. Zinc Homeostasis in the Human Digestive System
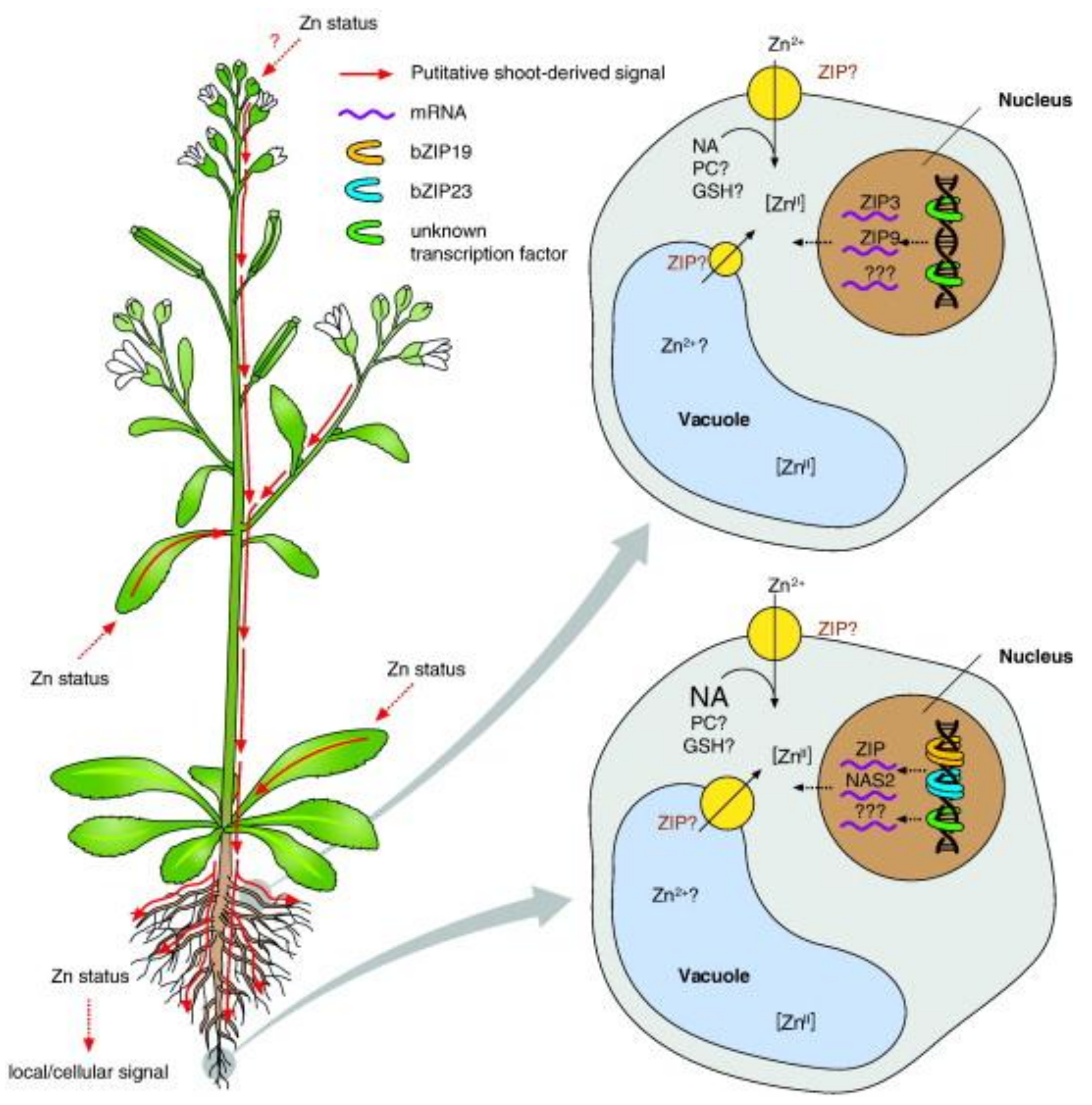
2.2. Zinc and Plant Biology
2.2.1. Mechanism of Zinc Uptake in Plants and Possibilities for Biotechnological Applications.
2.2.2. Lowering the Presence of Anti-Nutrients to Improve Zinc Bioavailability in Crops
2.2.3. Outcome of Zinc Variability in Soil on Plants
2.2.4. How Plants Subsist on Zinc Deficient Soils
2.2.5. How Plants Subsist on Zinc Toxic Soils
2.3. India as a Case Study
2.4. Biofortification Using Transgenic Plants
3. Conclusions
Conflicts of Interest
References
- Noulas, C.; Tziouvalekas, M.; Karyotis, T. Zinc in soils, water and food crops. J. Trace Elem. Med. Biol. 2018, 49, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Implication of Micronutrients in Agriculture and Health with Special Reference to Iron and Zinc. Available online: https://www.researchgate.net/publication/256117507_Implication_of_Micronutrients_in_Agriculture_and_Health_with_Special_Reference_to_Iron_and_Zinc (accessed on 18 November 2018).
- Milius, S. Worries Grow that Climate Change will Quietly Steal Nutrients from Major Food Crops. Science News Year in Review Nutrition, Climate, Sustainability. 2017. Available online: https://www.sciencenews.org/article/nutrition-climate-change-top-science-stories-2017-yir (accessed on 18 November 2018).
- Milius, S. Changing climate could worse foods’ nutrition. Sci. News 2017, 191, 14. [Google Scholar]
- Myers, S.S.; Wessells, K.R.; Kloog, I.; Zanobetti, A.; Schwartz, J. Effect of increased concentrations of atmospheric carbon dioxide on the global threat of zinc deficiency: A modelling study. Lancet Glob. Health. 2015, 3, 639. [Google Scholar] [CrossRef]
- Myers, S.S.; Zanobetti, A.; Kloog, I.; Leakey, A.D.B.; Bloom, A.J.; Carlisle, E.; Dietterich, L.H.; Fitzgerald, G.; Hasegawa, T.; Holbrook, N.M.; et al. Increasing CO2 threatens human nutrition. Nature 2014, 510, 139. [Google Scholar] [CrossRef] [PubMed]
- Hotz, C.; McClafferty, B. From harvest to health: Challenges for developing biofortified staple foods and determining their impact on micronutrient status. Food Nutr. Bull. 2007, 28, S271–S279. [Google Scholar] [CrossRef] [PubMed]
- Mayer, J.E.; Pfeiffer, W.H.; Beyer, P. Biofortified crops to alleviate micronutrient malnutrition. Curr. Opin. Plant Biol. 2008, 11, 166–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, G. Nutritional enhancement of plants. Curr. Opin. Biotechnol. 2003, 14, 221–225. [Google Scholar] [CrossRef]
- Wessels, I.; Maywald, M.; Rink, L. Zinc as a gatekeeper of immune function. Nutrients 2017, 9, 1286. [Google Scholar] [CrossRef]
- Bouis, H.E.; Hotz, C.; McClafferty, B.; Meenakshi, J.V.; Pfeiffer, W.H. Biofortification: A new tool to reduce micronutrient malnutrition. Food Nutr. Bull. 2011, 32, S31–S40. [Google Scholar] [CrossRef] [PubMed]
- Gilani, G.S.; Nasim, A. Impact of foods nutritionally enhanced through biotechnology in alleviating malnutrition in developing countries. J. AOAC Int. 2007, 90, 1440–1444. [Google Scholar] [PubMed]
- King, J.C.; Brown, K.H.; Gibson, R.S.; Krebs, N.F.; Lowe, N.M.; Siekmann, J.H.; Raiten, D.J. Biomarkers of Nutrition for Development (BOND)-Zinc Review. J. Nutr. 2016, 146, 858S–885S. [Google Scholar] [CrossRef] [PubMed]
- Krebs, N.F. Overview of zinc absorption and excretion in the human gastrointestinal tract. J. Nutr. 2000, 130, 1374S–1377S. [Google Scholar] [CrossRef] [PubMed]
- Olsen, L.I.; Palmgren, M. Many rivers to cross: The journey of zinc from soil to seed. Front. Plant Sci. 2014, 5, 30. [Google Scholar] [CrossRef] [PubMed]
- Improving the zinc-use efficiency in plants: A review. Available online: https://www.researchgate.net/publication/319503619 (accessed on 18 November 2018).
- The Zinc Homeostasis Network of Land Plants. Available online: https://www.sciencedirect.com/science/article/pii/S0167488912001322. (accessed on 18 November 2018).
- Johnson-Beebout, S.E.; Goloran, J.B.; Rubianes, F.H.; Jacob, J.D.; Castillo, O.B. Zn uptake behavior of rice genotypes and its implication on grain Zn biofortification. Sci. Rep. 2016, 6, 38301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subbaiah, L.V.; Prasad, T.N.; Krishna, T.G.; Sudhakar, P.; Reddy, B.R.; Pradeep, T. Novel effects of nanoparticulate delivery of zinc on growth, productivity, and zincbiofortification in maize (Zea mays L.). J. Agric. Food Chem. 2016, 64, 3778–3788. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, T.; Ishikawa, S.; Fujimaki, S. Route and regulation of zinc, cadmium, and iron transport in rice plants (oryza sativa l.) during vegetative growth and grain filling: Metal transporters, metal speciation, grain cd reduction and Zn and Fe biofortification. Int. J. Mol. Sci. 2015, 16, 19111–19129. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B. Genetically modified plants for improved trace element nutrition. J. Nutr. 2003, 133, 1490S–1493S. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—Iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]
- Ali, N.; Paul, S.; Gayen, D.; Sarkar, S.N.; Datta, S.K.; Datta, K. RNAi mediated down regulation of myo-inositol-3-phosphate synthase to generate low phytaterice. Rice 2013, 6, 12. [Google Scholar] [CrossRef]
- Li, W.X.; Zhao, H.J.; Pang, W.Q.; Cui, H.R.; Poirier, Y.; Shu, Q.Y. Seed-specific silencing of OsMRP5 reduces seed phytic acid and weight in rice. Transgenic Res. 2014, 23, 585–599. [Google Scholar] [CrossRef]
- Sakai, H.; Iwai, T.; Matsubara, C.; Usui, Y.; Okamura, M.; Yatou, O.; Terada, Y.; Aoki, N.; Nishida, S.; Yoshida, K.T. A decrease in phytic acid content substantially affects the distribution of mineral elements within rice seeds. Plant Sci. 2015, 238, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Shohag, M.J.; Yang, X. Biofortification and bioavailability of rice grain zinc as affected by different forms of foliar zinc fertilization. PLoS ONE 2012, 7, e45428. [Google Scholar] [CrossRef] [PubMed]
- Poblaciones, M.J.; Rengel, Z. Soil and foliar zinc biofortification in field pea (Pisum sativum L.): Grain accumulation and bioavailability in raw and cooked grains. Food Chem. 2016, 212, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Kappara, S.; Neelamraju, S.; Ramanan, R. Down regulation of a heavy metal transporter gene influences several domestication traits and grain Fe-Zn content in rice. Plant. Sci. 2018, 276, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Tiong, J.; McDonald, G.; Genc, Y.; Shirley, N.; Langridge, P.; Huang, C.Y. Increased expression of six ZIP family genes by zinc (Zn) deficiency is associated with enhanced uptake and root-to-shoot translocation of Zn in barley (Hordeum vulgare). New Phytol. 2015, 207, 1097–1109. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, K.; Pang, Y.; Naveed, S.A.; Zhao, X.; Wang, X.; Wang, Y.; Dingkuhn, M.; Pasuquin, J.; Li, Z.; et al. QTL mapping and candidate gene analysis of ferrous iron and zinc toxicity tolerance at seedling stage in rice by genome-wide association study. BMC Genom. 2017, 18, 828. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.J.; Nestel, P.; Meenakshi, J.V.; Qaim, M.; Sachdev, H.P.; Bhutta, Z.A. Plant breeding to control zinc deficiency in India: How cost-effective is biofortification? Public Health Nutr. 2007, 10, 492–501. [Google Scholar] [CrossRef]
- Morrissey, J.; Guerinot, M.L. Iron uptake and transport in plants: The good, the bad, and the ionome. Chem. Rev. 2009, 109, 4553–4567. [Google Scholar] [CrossRef]
- Masuda, H.; Ishimaru, Y.; Aung, M.S.; Kobayashi, T.; Kakei, Y.; Takahashi, M.; Higuchi, K.; Nakanishi, H.; Nishizawa, N.K. Iron biofortification in rice by the introduction of multiple genes involved in iron nutrition. Sci. Rep. 2012, 2, 543. [Google Scholar] [CrossRef]
- Masuda, H.; Kobayashi, T.; Ishimaru, Y.; Takahashi, M.; Aung, M.S.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Iron-biofortification in rice by the introduction of three barley genes participated in mugineic acid biosynthesis with soybean ferritin gene. Front. Plant. Sci. 2013, 4, 132. [Google Scholar] [CrossRef]
- Aung, M.S.; Masuda, H.; Kobayashi, T.; Nakanishi, H.; Yamakawa, T.; Nishizawa, N.K. Iron biofortification of myanmar rice. Front. Plant. Sci. 2013, 4, 158. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Ali, N.; Datta, S.K.; Datta, K. Development of an iron-enriched high-yieldings indica rice cultivar by introgression of a high-iron trait from transgenic iron-biofortifiedrice. Plant. Foods Hum. Nutr. 2014, 203–208. [Google Scholar] [CrossRef]
- Tan, S.; Han, R.; Li, P.; Yang, G.; Li, S.; Zhang, P.; Wang, W.B.; Zhao, W.Z.; Yin, L.P. Over-expression of the MxIRT1 gene increases iron and zinc content in rice seeds. Transgenic Res. 2015, 24, 109–122. [Google Scholar] [CrossRef]
- Trijatmiko, K.R.; Dueñas, C.; Tsakirpaloglou, N.; Torrizo, L.; Arines, F.M.; Adeva, C.; Balindong, J.; Oliva, N.; Sapasap, M.V.; Borrero, J.; et al. Biofortified indica rice attains iron and zinc nutrition dietary targets in the field. Sci. Rep. 2016, 6, 19792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banakar, R.; Alvarez Fernandez, A.; Díaz-Benito, P.; Abadia, J.; Capell, T.; Christou, P. Phytosiderophores determine thresholds for iron and zinc accumulation in biofortified rice endosperm while inhibiting the accumulation of cadmium. J. Exp. Bot. 2017, 68, 4983–4995. [Google Scholar] [CrossRef] [Green Version]
- Bouis, H. Reducing Mineral and Vitamin Deficiencies through Biofortification: Progress Under HarvestPlus. World Rev. Nutr. Diet. 2018, 118, 112–122. [Google Scholar] [CrossRef]
- Sayre, R.; Beeching, J.R.; Cahoon, E.B.; Egesi, C.; Fauquet, C.; Fellman, J.; Fregene, M.; Gruissem, W.; Mallowa, S.; Manary, M.; et al. The BioCassava plus program: Biofortification of cassava for sub-Saharan Africa. Annu. Rev. Plant. Biol. 2011, 62, 251–272. [Google Scholar] [CrossRef]



© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hefferon, K. Biotechnological Approaches for Generating Zinc-Enriched Crops to Combat Malnutrition. Nutrients 2019, 11, 253. https://doi.org/10.3390/nu11020253
Hefferon K. Biotechnological Approaches for Generating Zinc-Enriched Crops to Combat Malnutrition. Nutrients. 2019; 11(2):253. https://doi.org/10.3390/nu11020253
Chicago/Turabian StyleHefferon, Kathleen. 2019. "Biotechnological Approaches for Generating Zinc-Enriched Crops to Combat Malnutrition" Nutrients 11, no. 2: 253. https://doi.org/10.3390/nu11020253
APA StyleHefferon, K. (2019). Biotechnological Approaches for Generating Zinc-Enriched Crops to Combat Malnutrition. Nutrients, 11(2), 253. https://doi.org/10.3390/nu11020253