Effect of Enzymatic Treatment of Chrysanthemum Indicum Linné Extracts on Lipid Accumulation and Adipogenesis in High-Fat-Diet-Induced Obese Male Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Preparation of the Ethanol Extract of Enzymatic CI (CIVT)
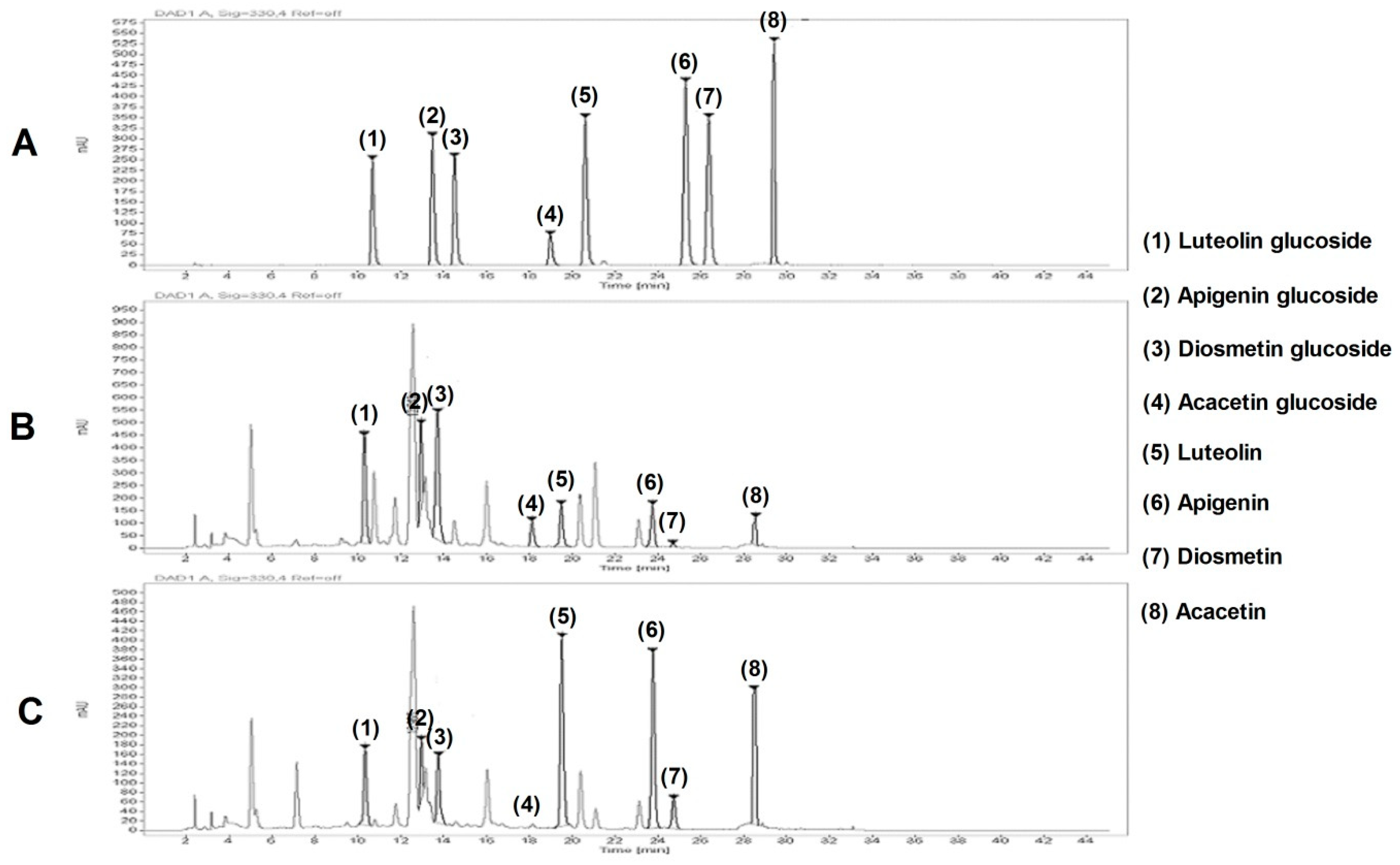
2.3. High Performance Liquid Chromatography (HPLC) Analysis
2.4. Animals and Treatment
2.5. Micro Computed Tomography (Micro-CT)
2.6. Serum and Tissue Preparation
2.7. Serum Biochemical Analysis
2.8. Enzyme-Linked Immunosorbent Assay (ELISA)
2.9. Histological Analysis
2.10. Western Blot Analysis
2.11. Statistical Analysis
3. Results
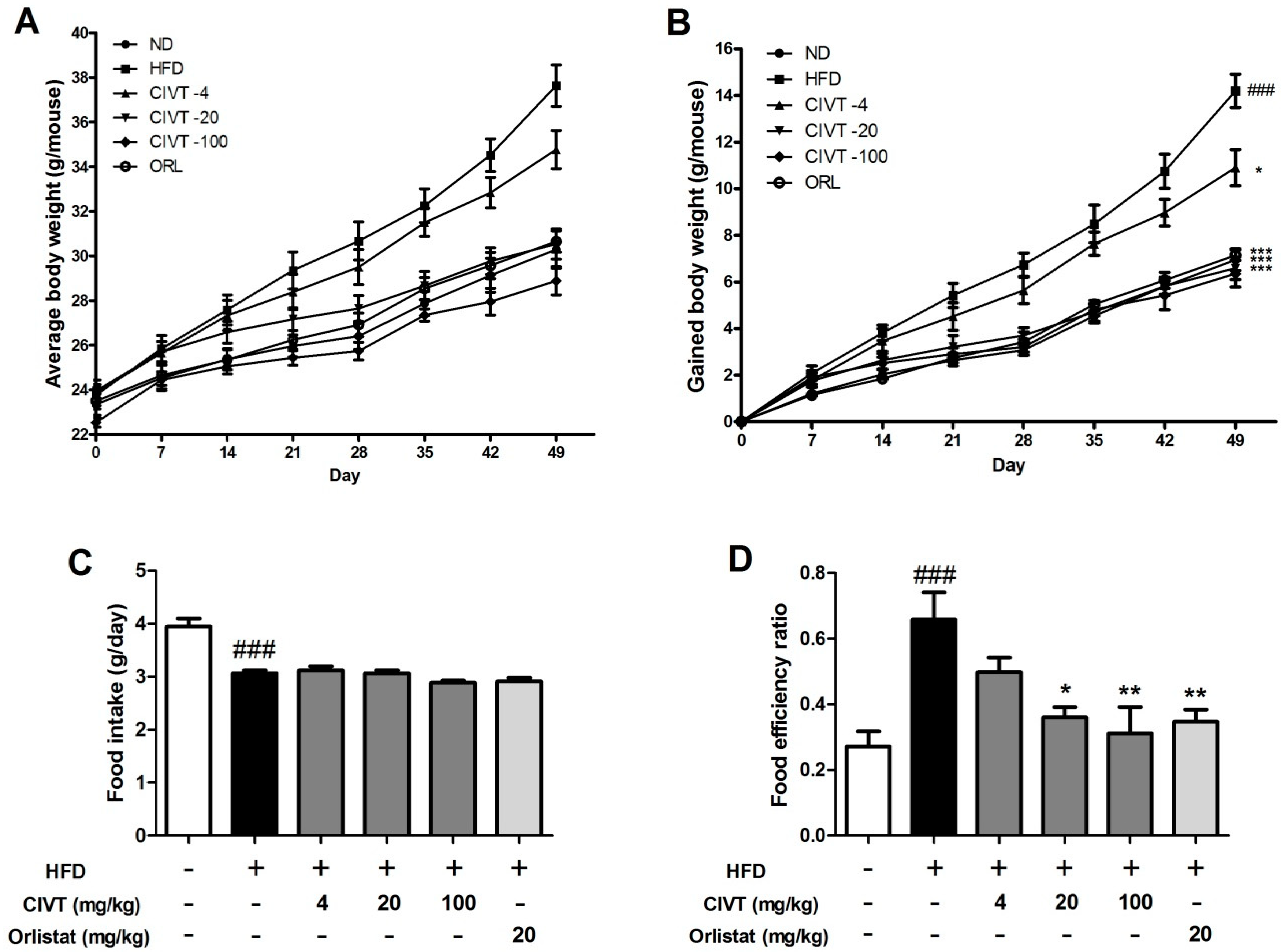
3.1. CIVT Reduces Body Weight Gain and Food Efficiency Ratio in HFD-Fed Obese Mice
3.2. Effects of CIVT on Organ Weight in HFD-Fed Obese Mice
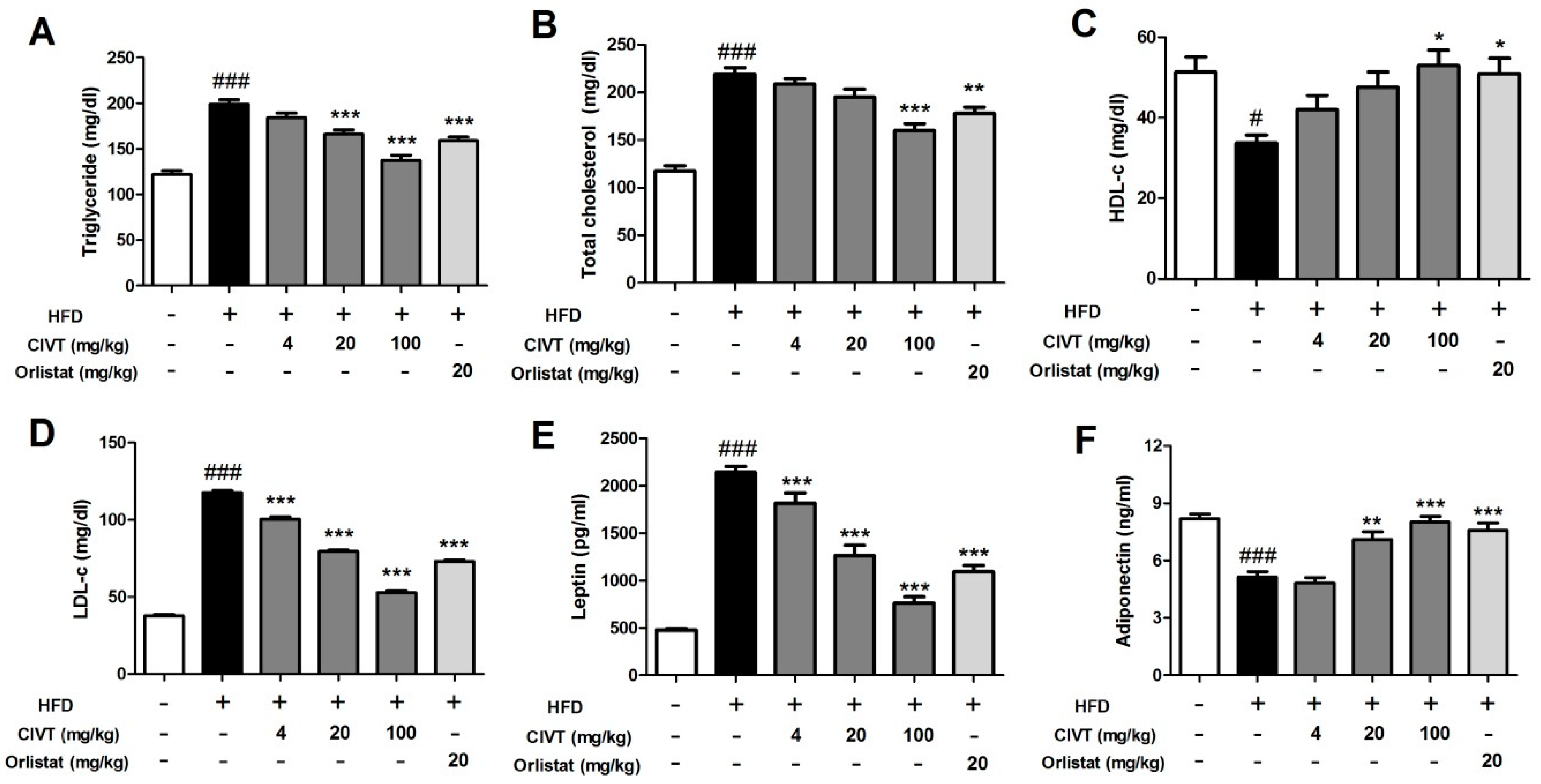
3.3. CIVT Regulates Serum Lipid Parameters in HFD-Fed Obese Mice
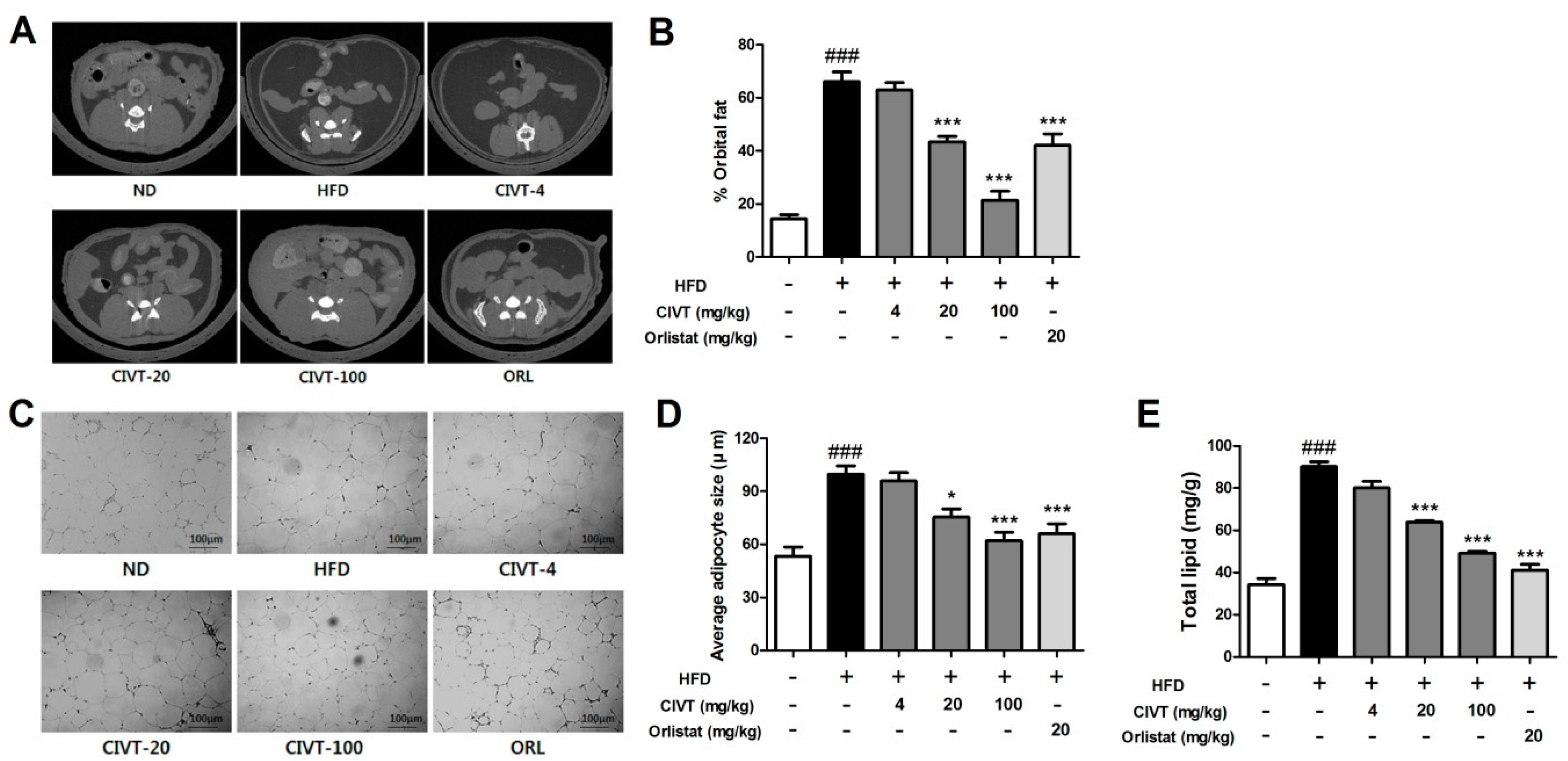
3.4. CIVT Decreases Fat Deposition, Adipocytes Size and Total Lipid Levels in the Liver Tissues in HFD-Fed Obese Mice
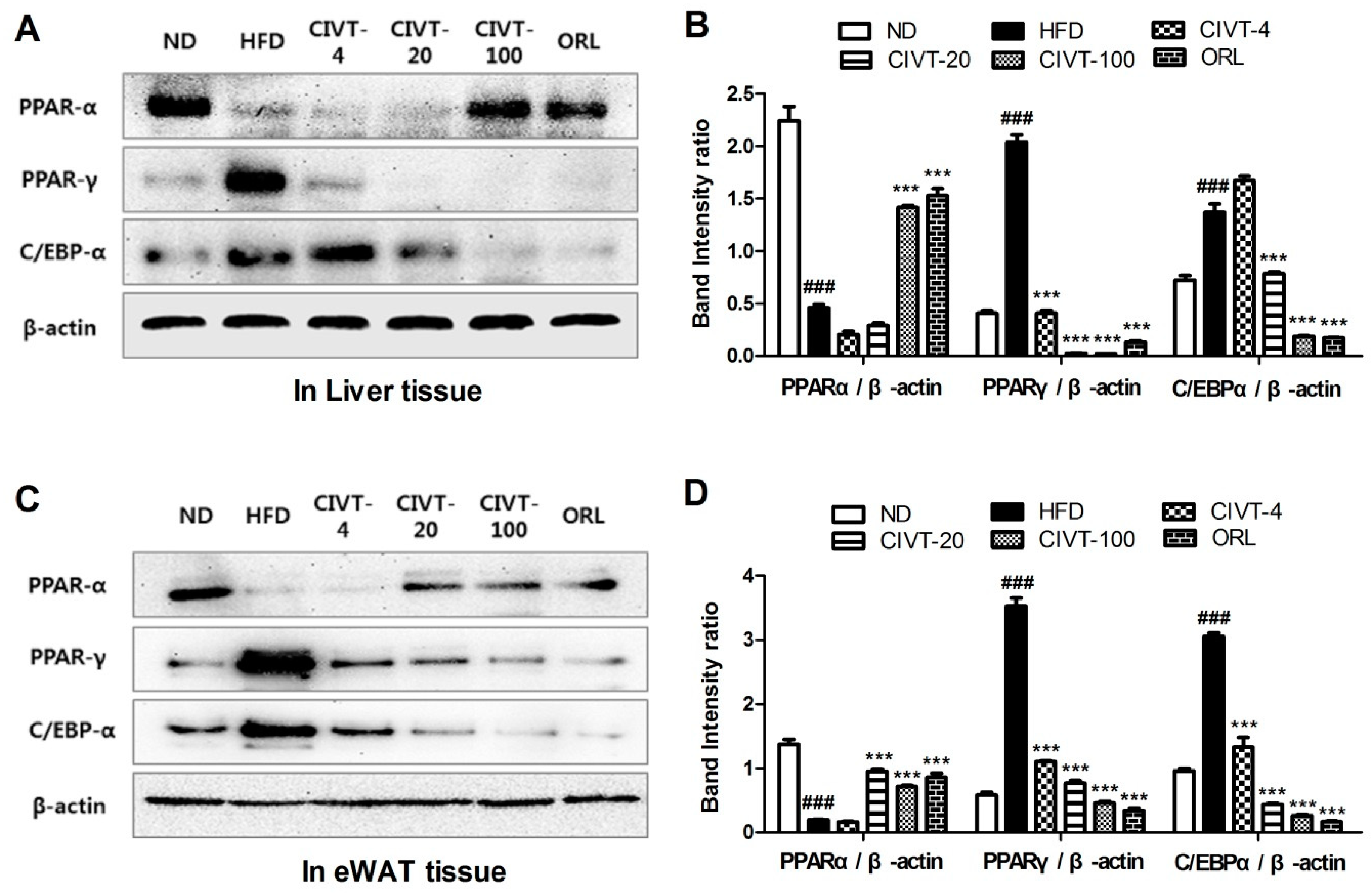
3.5. CIVT Regulates the Levels of Adipogenesis-Related Proteins in Liver Tissues and in eWAT, of HFD-Fed Obese Mice
3.6. CIVT Is More Effective Than CIEE, in the Lipid Accumulation
3.7. HPLC Analysis of CIEE and CIVT
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Hong, Y.H.; Jung, E.Y.; Park, Y.H.; Shin, K.S.; Kim, T.Y.; Yu, K.W.; Chang, W.J.; Suh, H.J. Enzymatic improvement in the polyphenol extractability and antioxidant activity of green tea extracts. Biosci. Biotechnol. Biochem. 2013, 77, 22–29. [Google Scholar] [CrossRef]
- Guan, X.; Yao, H. Optimization of Viscozyme L-assisted extraction of oat bran protein using response surface methodology. Food Chem. 2008, 106, 345–351. [Google Scholar] [CrossRef]
- Lu, M.J.; Chen, C. Enzymatic modification by tannase increases the antioxidant activity of green tea. Food. Res. Int. 2008, 41, 130–137. [Google Scholar] [CrossRef]
- De Ferranti, S.; Mozaffarian, D. The perfect storm: obesity, adipocyte dysfunction, and metabolic consequences. Clin. Chem. 2008, 54, 945–955. [Google Scholar] [CrossRef]
- Masuo, K.; Tuck, M.L.; Lambert, G.W. Hypertension and diabetes in obesity. Int. J. Hypertens. 2011, 2011, 695869. [Google Scholar] [CrossRef]
- Charlton, M. Obesity, hyperlipidemia, and metabolic syndrome. Liver Transpl. 2009, 15, S83–S89. [Google Scholar] [CrossRef]
- Rocha, V.Z.; Libby, P. Obesity, inflammation, and atherosclerosis. Nat. Rev. Cardiol. 2009, 6, 399–409. [Google Scholar] [CrossRef]
- Kawada, T.; Takahashi, N.; Fushiki, T. Biochemical and physiological characteristics of fat cell. J. Nutr. Sci. Vitaminol. 2001, 47, 1–12. [Google Scholar] [CrossRef]
- Jo, J.; Gavrilova, O.; Pack, S.; Jou, W.; Mullen, S.; Sumner, A.E.; Cushman, S.W.; Periwal, V. Hypertrophy and/or hyperplasia: dynamics of adipose tissue growth. PLoS. Comput. Biol. 2009, 5, e1000324. [Google Scholar] [CrossRef]
- Wang, Q.A.; Caroline, T.; Gupta, R.K.; Scherer, P.E. Tracking adipogenesis during white adipose tissue development, expansion and regeneration. Nat. Med. 2013, 19, 1338. [Google Scholar] [CrossRef]
- Liang, G.; Xi, L.; Qi-Qun, T. Transcriptional regulation of adipocyte differentiation: a central role for CCAAT/Enhancer-binding protein (C/EBP) β. J. Biol. Chem. 2015, 290, 755–761. [Google Scholar]
- Gumbilai, V.; Ebihara, K.; Aizawa-Abe, M.; Ebihara, C.; Zhao, M.; Yamamoto, Y.; Mashimo, T.; Hosoda, K.; Serikawa, T.; Nakao, K. Fat mass reduction with adipocyte hypertrophy and insulin resistance in heterozygous PPARγ mutant rats. Mol. Diabetes 2016, 65, 2954–2965. [Google Scholar] [CrossRef]
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and adipogenesis. Eur. J. Cell. Biol. 2013, 92, 229–236. [Google Scholar] [CrossRef]
- Roberts, C.K.; Sindhu, K.K. Oxidative stress and metabolic syndrome. Life Sci. 2009, 84, 705–712. [Google Scholar] [CrossRef]
- Moseti, D.; Regassa, A.; Kim, W.K. Molecular regulation of adipogenesis and potential anti-adipogenic bioactive molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef]
- Cheng, W.; Li, J.; You, T.; Hu, C. Anti-inflammatory and immunomodulatory activities of the extracts from the inflorescence of Chrysanthemum indicum Linne. J. Ethnopharmacol. 2005, 101, 334–337. [Google Scholar] [CrossRef]
- Shunying, Z.; Yang, Y.; Huaidong, Y.; Yue, Y.; Guolin, Z. Chemical composition and antimicrobial activity of the essential oils of Chrysanthemum indicum. J. Ethnopharmacol. 2005, 96, 151–158. [Google Scholar] [CrossRef]
- Cha, J.Y.; Nepali, S.; Lee, H.Y.; Hwang, S.W.; Choi, S.Y.; Yeon, J.M.; Song, B.J.; Kim, D.K.; Lee, Y.M. Chrysanthemum indicum L. ethanol extract reduces high-fat diet-induced obesity in mice. Exp. Ther. Med. 2018, 15, 5070–5076. [Google Scholar]
- Nepali, S.; Cha, J.Y.; Ki, H.H.; Lee, H.Y.; Kim, Y.H.; Kim, D.K.; Song, B.J.; Lee, Y.M. Chrysanthemum indicum inhibits adipogenesis and activates the AMPK pathway in high-fat-diet-induced obese mice. Am. J. Chin. Med. 2018, 46, 119–136. [Google Scholar] [CrossRef]
- Jain, S.S.; Ramanand, S.J.; Ramanand, J.B.; Akat, P.B.; Patwardhan, M.H.; Joshi, S.R. Evaluation of efficacy and safety of orlistat in obese patients. Indian J. Endocrinol. Metab. 2011, 15, 99–104. [Google Scholar]
- Padwal, R.S.; Majumdar, S.R. Drug treatments for obesity: Orlistat, sibutramine, and rimonabant. Lancet 2007, 369, 71–77. [Google Scholar] [CrossRef]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar]
- Folch, J.; Mee, L.; Stanley, G.S.H. A simple method for the isolation and purification of total lipid from animal tissues. J. Biol. Chem. 1975, 226, 497–509. [Google Scholar]
- Singla, P.; Bardoloi, A.; Parkash, A.A. Metabolic effects of obesity: A review. World J. Diabetes 2010, 1, 76–88. [Google Scholar] [CrossRef]
- Medanić, D.; Pucarin-Cvetković, J. Obesity—A public health problem and challenge. Acta Med. Croatica 2012, 66, 347–355. [Google Scholar]
- Fock, K.M.; Khoo, J. Diet and exercise in management of obesity and overweight. J. Gastroenterol. Hepatol. 2013, 4, 59–63. [Google Scholar] [CrossRef]
- Derosa, G.; Cicero, A.F.; Murdolo, G.; Piccinni, M.N.; Fogari, E.; Bertone, G.; Ciccarelli, L.; Fogari, R. Efficacy and safety comparative evualation of orlistat and sibutramine treatment in hypertensive obese patients. Diabeted. Obes. Metab. 2005, 7, 47–55. [Google Scholar] [CrossRef]
- Conforti, F.; Pan, M. Natural products in anti-obesity therapy. Molecules 2016, 21, 1750. [Google Scholar] [CrossRef]
- Khan, M.; Joseph, F. Adipose tissue and adipokines: the association with and application of adipokines in obesity. Scientifica 2014, 2014, 32859. [Google Scholar] [CrossRef]
- Klok, M.D.; Jakobsdottir, S.; Drent, M.L. The role of leptin and ghrelin in the regulation of food intake and body weight in humans: a review. Obes. Rev. 2007, 8, 21–34. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Minokoshi, Y.; Ito, Y.; Waki, H.; Uchida, S.; Yamashita, S.; Noda, M.; Kita, S.; Ueki, K.; et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat. Med. 2002, 8, 1288–1295. [Google Scholar] [CrossRef]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef]
- Evans, R.M.; Barish, G.D.; Wang, Y.X. PPARs and the complex journey to obesity. Nat. Med. 2004, 10, 355–361. [Google Scholar] [CrossRef]
- Ambele, M.A.; Dessels, C.; Durandt, C.; Pepper, M.S. Genome-wide analysis of gene expression during adipogenesis in human adipose-derived stromal cells reveals novel patterns of gene expression during adipocyte differentiation. Stem. Cell. Res. 2016, 16, 725–734. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Morikawa, T.; Murakami, T.; Toguchida, I.; Harima, S.; Matsuda, H. Medicinal flowers. I. Aldose reductase inhibitors and three new eudesmane-type sesquiterpenes, kikkanols A, B, and C, from the flowers of Chrysanthemum indicum L. Chem. Pharm. Bull. 1999, 47, 340–345. [Google Scholar] [CrossRef]
- Thangasamy, T.; Sittadjody, S.; Burd, R. Chapter 27–Quercetin: a potential complementary and alternative cancer therapy. Complement. Altern. Ther. Aging Popul. 2009, 563–584. [Google Scholar] [CrossRef]
- Athukorala, Y.; Kim, K.N.; Jeon, Y.J. Antiproliferative and antioxidant properties of an enzymatic hydrolysate from brown alga, Ecklonia cava. Food Chem. Toxicol. 2006, 44, 1065–1074. [Google Scholar] [CrossRef]
- Han, Y.N.; Noh, D.B.; Hart, D.S. Studies on the monoamine oxidase inhibitors of medicinal plants I. isolation of MAO-B inhibitors from Chrysanthemum indicum, Arch. Pharm. Res. 1987, 10, 142–147. [Google Scholar]
- Liou, C.J.; Wu, S.J.; Chen, L.C.; Yeh, K.W.; Chen, C.Y.; Huang, W.C. Acacetin from traditionally used saussurea involucrata Kar. et Kir. suppressed adipogenesis in 3T3-L1 adipocytes and attenuated lipid accumulation in obese mice. Front. Pharmacol. 2017, 8, 589. [Google Scholar] [CrossRef]
- Liu, Y.; Fu, X.; Lan, N.; Li, S.; Zhang, J.; Wang, S.; Li, C.; Shang, Y.; Huang, T.; Zhang, L. Luteolin protects against high fat diet-induced cognitive deficits in obesity mice. Behav. Brain. Res. 2014, 267, 178–188. [Google Scholar] [CrossRef]
- Jung, U.J.; Cho, Y.Y.; Choi, M.S. Apigenin ameliorates dyslipidemia, hepatic steatosis and insulin resistance by modulating metabolic and transcriptional profiles in the liver of high-fat diet-induced obese mice. Nutrients 2016, 8, 305. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ND | HFD | CIVT-4 | CIVT-20 | CIVT-100 | ORL | |
|---|---|---|---|---|---|---|
| eWAT (g) | 0.56 ± 0.14 | 2.03 ± 0.08 # | 1.83 ± 0.12 | 0.87 ± 0.17 c | 0.81 ± 0.09 c | 1.20 ± 0.14 b |
| Liver (g) | 1.11 ± 0.06 | 1.55 ± 0.08 # | 1.40 ± 0.06 | 1.23 ± 0.07 a | 1.02 ± 0.03 c | 1.31 ± 0.04 |
| Spleen (g) | 0.09 ± 0.01 | 0.12 ± 0.01 # | 0.11 ± 0.01 | 0.09 ± 0.01 a | 0.09 ± 0.01 b | 0.09 ± 0.01 b |
| Kidney (g) | 0.33 ± 0.01 | 0.42 ± 0.01 | 0.39 ± 0.02 | 0.37 ± 0.01 | 0.36 ± 0.01 | 0.38 ± 0.01 |
| ND | HFD | CIEE-100 | CIVT-100 | |
|---|---|---|---|---|
| Gained body weight (g/mouse) | 6.95 ± 0.46 | 14.20 ± 0.72 # | 8.73 ± 0.31 c | 6.35 ± 0.57 c, d |
| Food efficiency ratio | 0.27 ± 0.05 | 0.65 ± 0.08 # | 0.5 ± 0.03 a | 0.31 ± 0.08 b |
| eWAT (g) | 0.56 ± 0.14 | 2.03 ± 0.08 # | 1.27 ± 0.12 c | 0.81 ± 0.09 c, d |
| Liver (g) | 1.11 ± 0.06 | 1.55 ± 0.08 # | 1.12 ± 0.05 a | 1.02 ± 0.03 c, d |
| Orbital fat (%) | 14.32 ± 1.69 | 66.06 ± 3.65 # | 42.1 ± 4.31 c | 21.45 ± 3.38 c, e |
| Leptin (pg/mL) | 477.0 ± 11.37 | 2140.0 ± 65 # | 1025 ± 112 c | 761.8 ± 63.7 c, e |
| Adiponectin (ng/mL) | 8.18 ± 0.25 | 5.14 ± 0.29 # | 6.79 ± 0.20 c | 8.02 ± 0.29 c, d |
| CIEE | CIVT | |
|---|---|---|
| Yields (%) | 27.7 | 48.5 |
| Luteolin glucoside (mg/g) | 13.89 | 6.86 |
| Apigenin glucoside (mg/g) | 13.00 | 5.07 |
| Diosmetin glucoside (mg/g) | 17.80 | 6.32 |
| Acacetin glucoside (mg/g) | 2.23 | 0.71 |
| Luteolin (mg/g) | 1.57 | 4.41 |
| Apigenin (mg/g) | 1.51 | 4.04 |
| Diosmetin (mg/g) | 0.24 | 0.93 |
| Acacetin (mg/g) | 0.80 | 2.98 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-H.; Moon, J.-M.; Kim, Y.-H.; Lee, B.; Choi, S.-Y.; Song, B.-J.; Kim, D.-K.; Lee, Y.-M. Effect of Enzymatic Treatment of Chrysanthemum Indicum Linné Extracts on Lipid Accumulation and Adipogenesis in High-Fat-Diet-Induced Obese Male Mice. Nutrients 2019, 11, 269. https://doi.org/10.3390/nu11020269
Lee J-H, Moon J-M, Kim Y-H, Lee B, Choi S-Y, Song B-J, Kim D-K, Lee Y-M. Effect of Enzymatic Treatment of Chrysanthemum Indicum Linné Extracts on Lipid Accumulation and Adipogenesis in High-Fat-Diet-Induced Obese Male Mice. Nutrients. 2019; 11(2):269. https://doi.org/10.3390/nu11020269
Chicago/Turabian StyleLee, Ji-Hyun, Joo-Myung Moon, Yoon-Hee Kim, Bori Lee, Sang-Yong Choi, Bong-Joon Song, Dae-Ki Kim, and Young-Mi Lee. 2019. "Effect of Enzymatic Treatment of Chrysanthemum Indicum Linné Extracts on Lipid Accumulation and Adipogenesis in High-Fat-Diet-Induced Obese Male Mice" Nutrients 11, no. 2: 269. https://doi.org/10.3390/nu11020269
APA StyleLee, J. -H., Moon, J. -M., Kim, Y. -H., Lee, B., Choi, S. -Y., Song, B. -J., Kim, D. -K., & Lee, Y. -M. (2019). Effect of Enzymatic Treatment of Chrysanthemum Indicum Linné Extracts on Lipid Accumulation and Adipogenesis in High-Fat-Diet-Induced Obese Male Mice. Nutrients, 11(2), 269. https://doi.org/10.3390/nu11020269