Daidzein Intake Is Associated with Equol Producing Status through an Increase in the Intestinal Bacteria Responsible for Equol Production

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Material and Methods
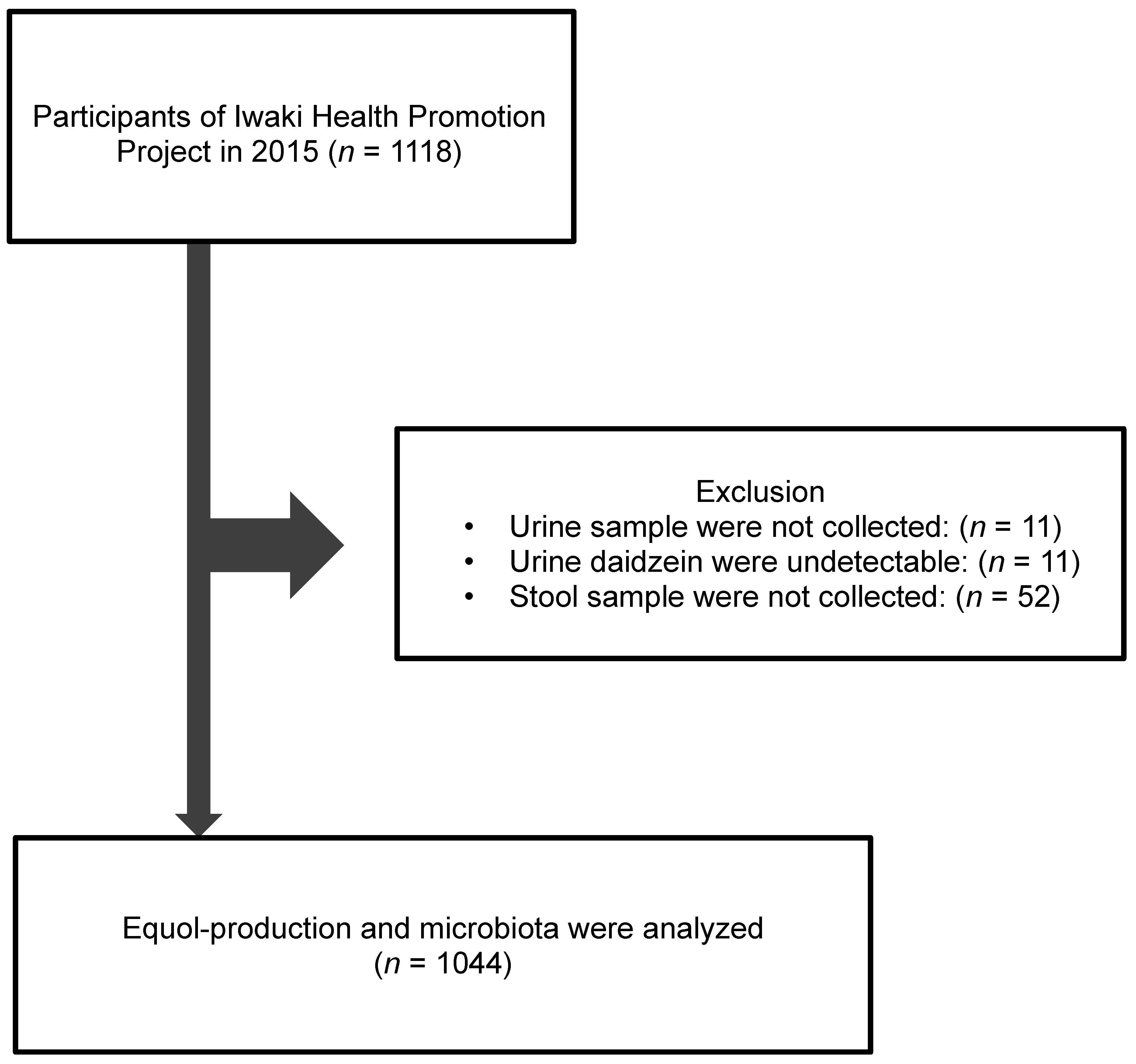
2.1. Study Subjects
2.2. Urine Concentration of Equol and Daidzein
2.3. Intake of Daidzein
2.4. Next Generation Sequence Analysis of Gut Microbiota
2.5. Statistical Analysis
2.6. Ethics Statement
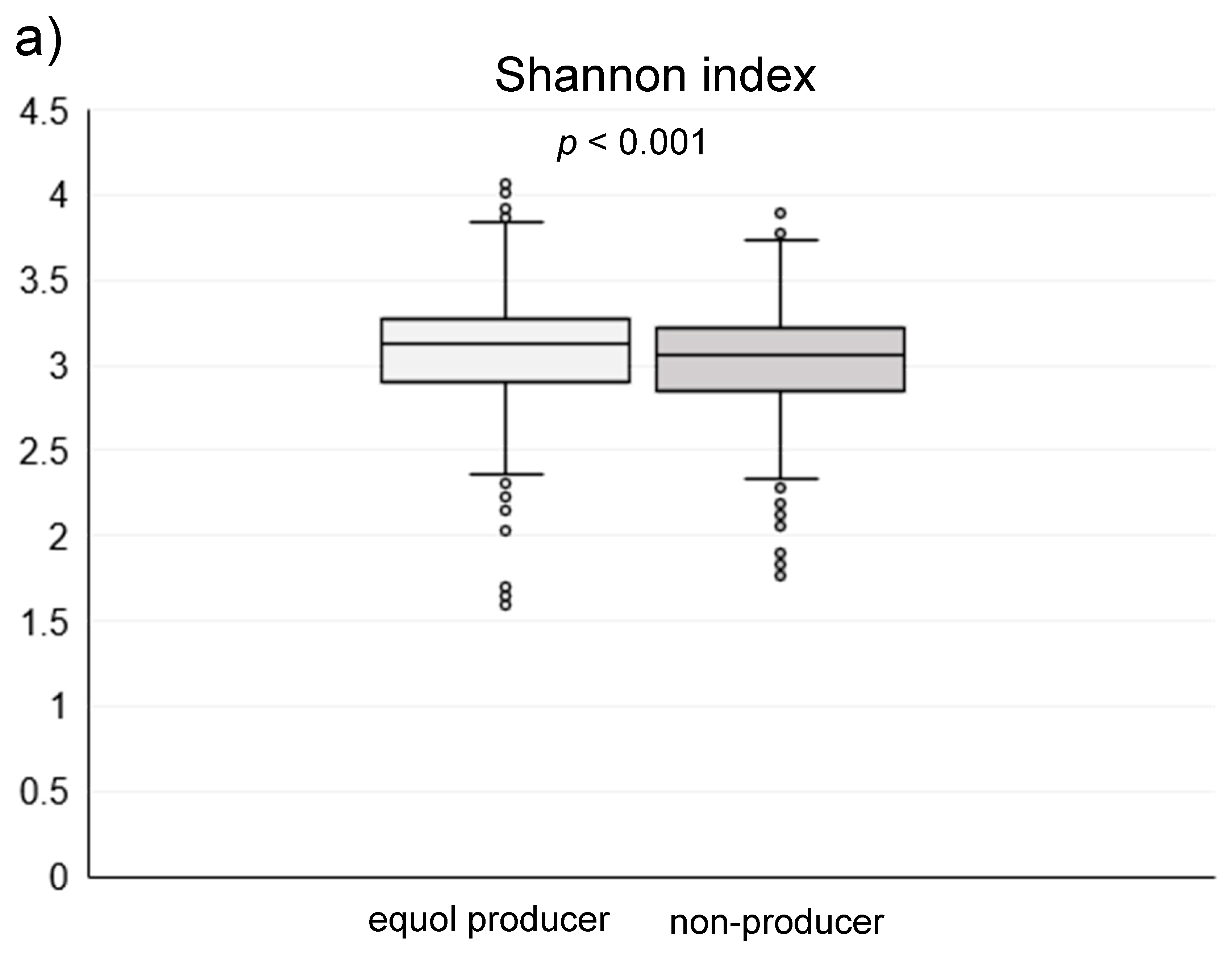
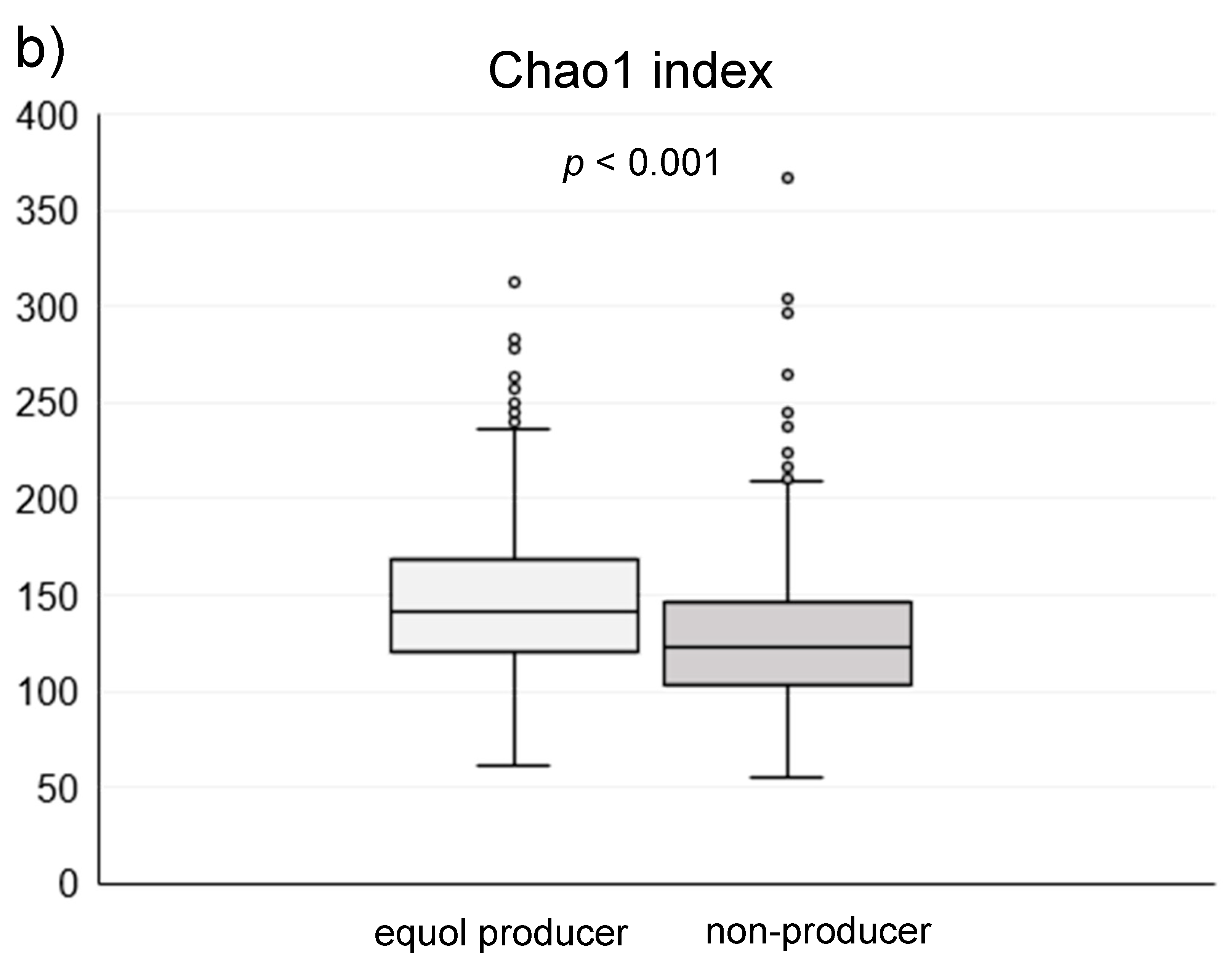
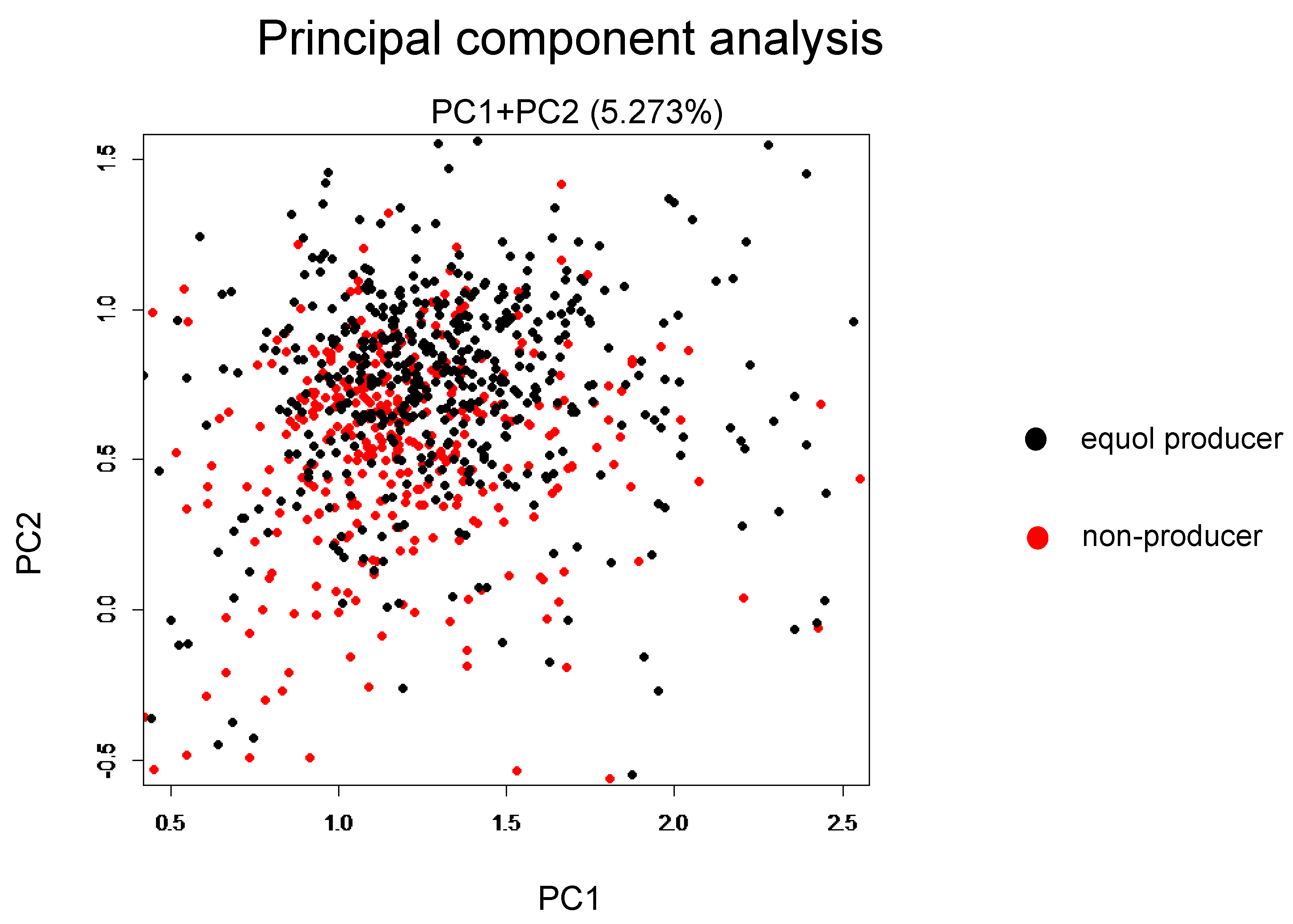
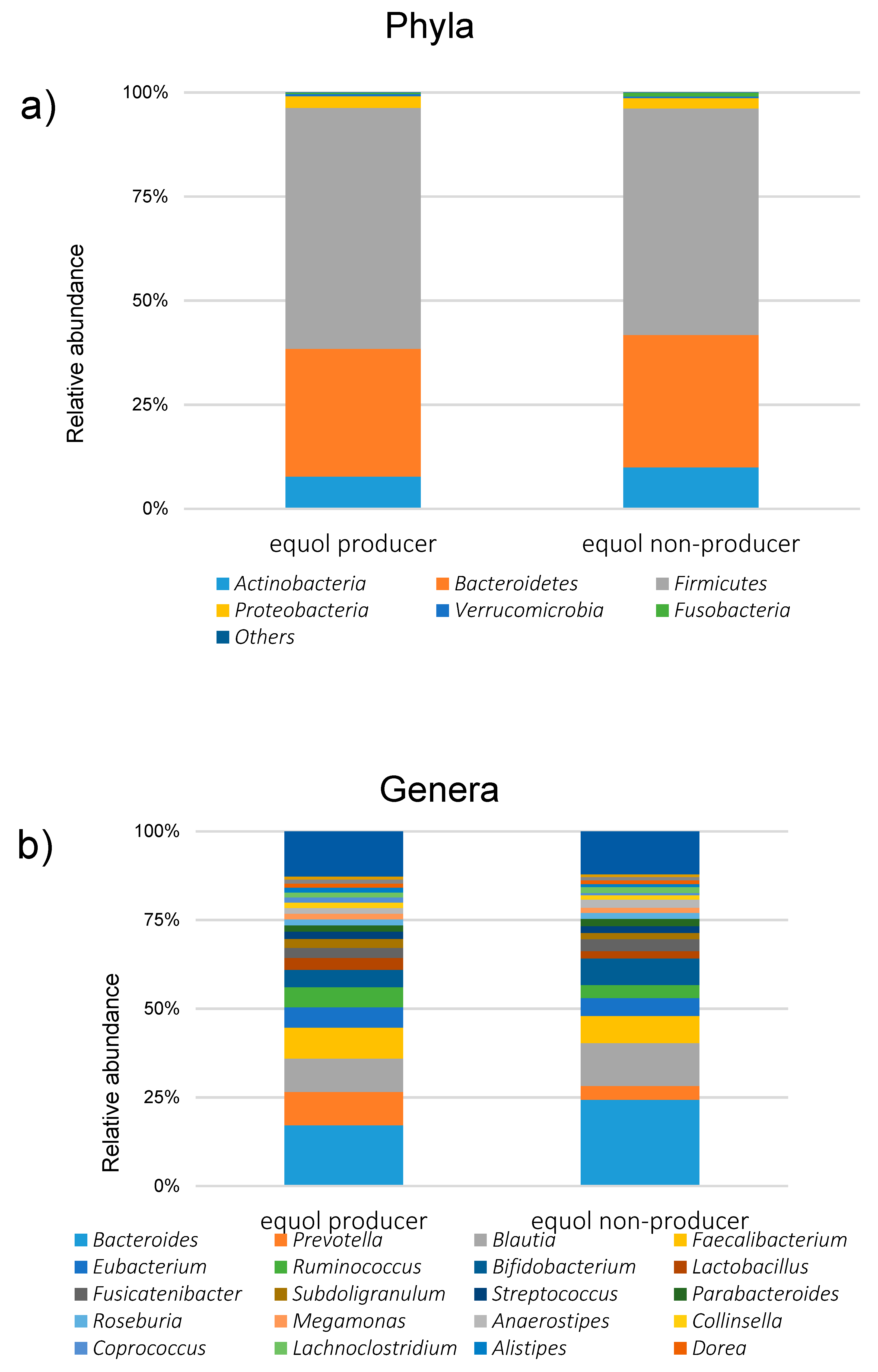
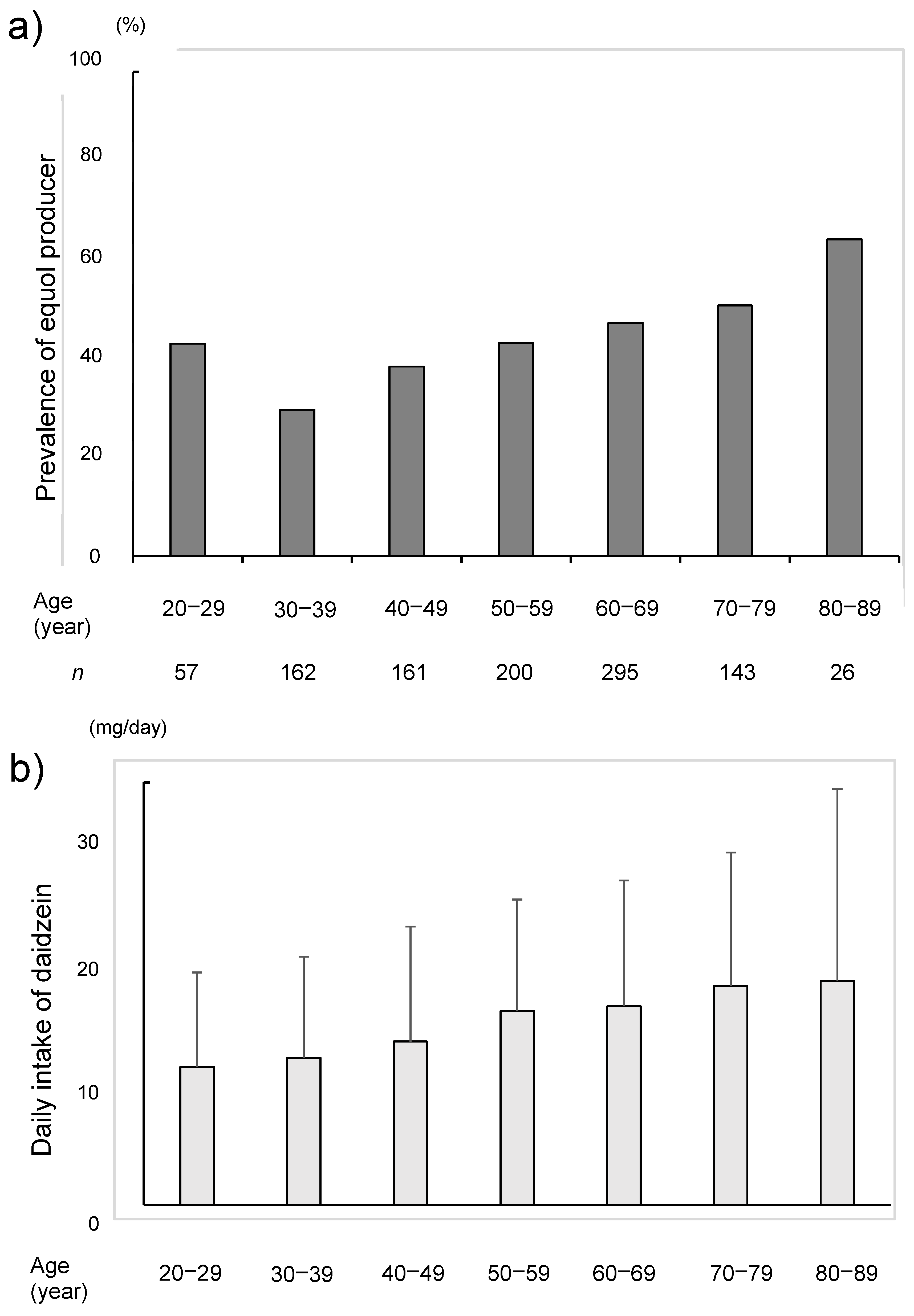
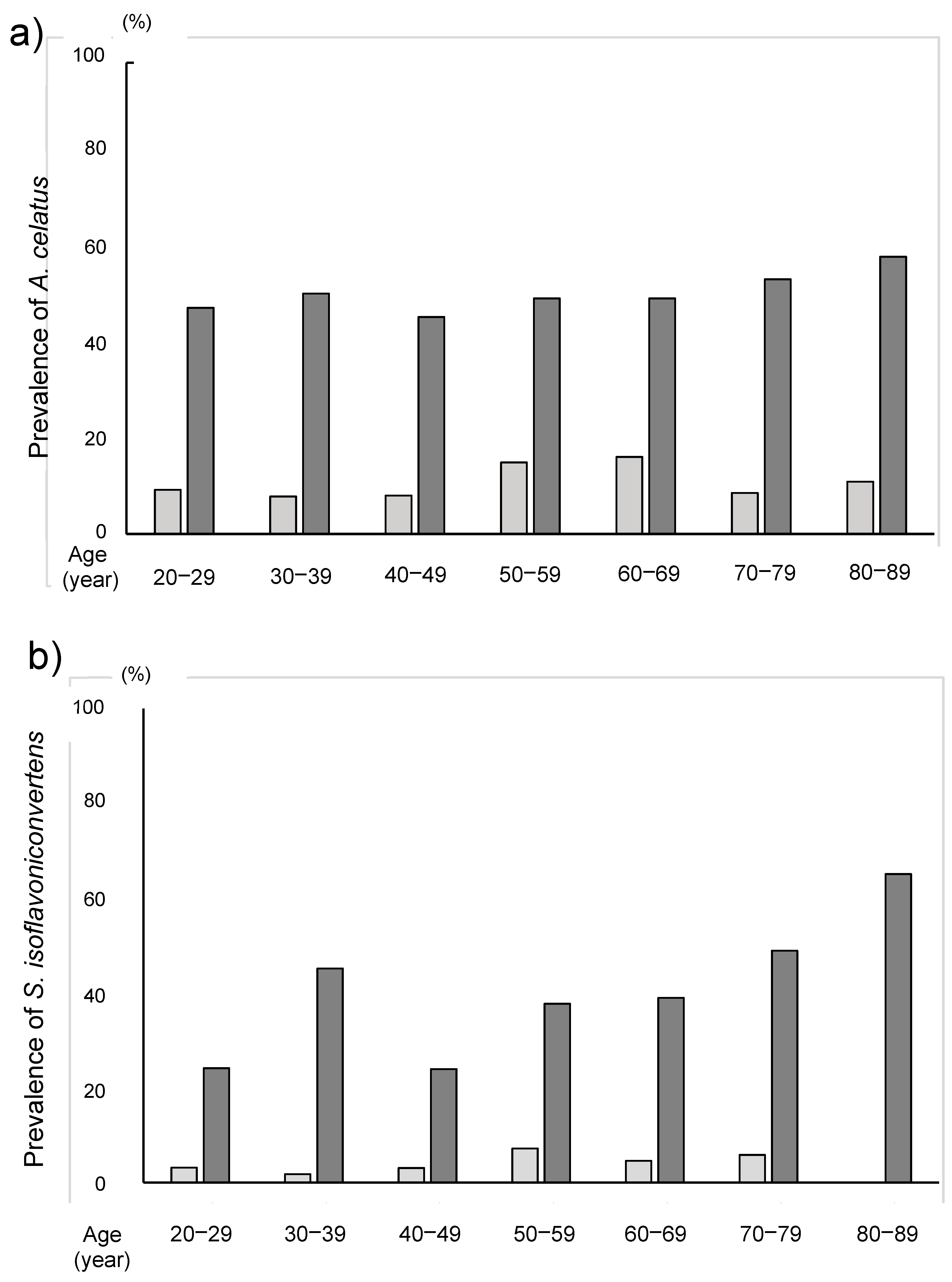
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Setchell, K.D.R.; Borriello, S.P.; Hulme, P.; Kirk, D.N.; Axelson, M. Nonsteroidal estrogens of dietary origin: Possible roles in hormone-dependent disease. Am. J. Clin. Nutr. 1984, 40, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Goodman, M.T.; Shvetsov, Y.B.; Wilkens, L.R.; Franke, A.A.; Le Marchand, L.; Kakazu, K.K.; Nomura, A.M.; Henderson, B.E.; Kolonel, L.N. Urinary phytoestrogen excretion and postmenopausal breast cancer risk: The multiethnic cohort study. Cancer Prev. Res. 2009, 2, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Akaza, H.; Miyanaga, N.; Takashima, N.; Naito, S.; Hirao, Y.; Tsukamoto, T.; Fujioka, T.; Mori, M.; Kim, W.J.; Song, J.M.; et al. Comparisons of percent equol producers between prostate cancer patients and controls: Case-controlled studies of isoflavones in Japanese, Korean and American residents. Jpn. J. Clin. Oncol. 2004, 34, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Spitznagel, E.L.; Bosland, M.C. Soy consumption and colorectal cancer risk in humans: A metaanalysis. Cancer Epidemiol. Biomark. Prev. 2010, 19, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Lydeking-Olsen, E.; Beck-Jensen, J.E.; Setchell, K.D.R.; Holm-Jensen, T. Soymilk or progesterone for prevention of bone loss—A 2 year randomized, placebo-controlled trial. Eur. J. Nutr. 2004, 43, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, A. Dietary phyto-oestrogens: Molecular mechanisms, bioavailability and importance to menopausal health. Nutr. Res. Rev. 2005, 18, 183–201. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.R.; Wiseman, H.; Sanders, T.A.; Adlercreutz, H.; Bowey, E.A. Interindividual variation in metabolism of soy isoflavones and lignans: Influence of habitual diet on equol production by the gut microflora. Nutr. Cancer 2000, 36, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, C.; Frankenfeld, C.L.; Lampe, J.W. Gut bacterial metabolism of the soy isoflavone daidzein: Exploring the relevance to human health. Exp. Biol. Med. 2005, 230, 155–170. [Google Scholar] [CrossRef]
- Lampe, J.W.; Karr, S.C.; Hutchins, A.M.; Slavin, J.L. Urinary equol excretion with a soy challenge: Influence of habitual diet. Proc. Soc. Exp. Biol. Med. 1998, 217, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Song, K.B.; Atkinson, C.; Frankenfeld, C.L.; Jokela, T.; Wähälä, K.; Thomas, W.K.; Lampe, J.W. Prevalence of daidzein-metabolizing phenotypes differs between Caucasian and Korean American women and girls. J. Nutr. 2006, 136, 1347–1351. [Google Scholar] [CrossRef] [PubMed]
- Ko, T.F.; Tsai, H.S.; Lin, S.M.; Liu, C.D.; Learn, S.P.; Chiou, R.Y. GC-MS determined distribution of urinary equol producers as affected by age, gender, and repeated ingestions of soymilk. J. Food Sci. 2010, 75, H306–H310. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D.; Faughnan, M.S.; Avades, T.; Zimmer-Nechemias, L.; Brown, N.M.; Wolfe, B.E.; Brashear, W.T.; Desai, P.; Oldfield, M.F.; Botting, N.P.; et al. Comparing the pharmacokinetics of daidzein and genistein with the use of 13C-labeled tracers in premenopausal women. Am. J. Clin. Nutr. 2003, 77, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Védrine, N.; Mathey, J.; Morand, C.; Brandolini, M.; Davicco, M.J.; Guy, L.; Rémésy, C.; Coxam, V.; Manach, C. One-month exposure to soy isoflavones did not induce the ability to produce equol in postmenopausal women. Eur. J. Clin. Nutr. 2006, 60, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Mathey, J.; Lamothe, V.; Coxam, V.; Potier, M.; Sauvant, P.; Bennetau-Pelissero, C. Concentrations of isoflavones in plasma and urine of post-menopausal women chronically ingesting high quantities of soy isoflavones. J. Pharm. Biomed. Anal. 2006, 41, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Frankenfeld, C.L.; Atkinson, C.; Thomas, W.K.; Gonzalez, A.; Jokela, T.; Wähälä, K.; Schwartz, S.M.; Li, S.S.; Lampe, J.W. High concordance of daidzein-metabolizing phenotypes in individuals measured 1 to 3 years apart. Br. J. Nutr. 2005, 94, 873–876. [Google Scholar] [CrossRef] [PubMed]
- Lundh, T.J.; Pettersson, H.; Kiessling, K.H. Liquid chromatographic determination of the estrogens daidzein, formononetin, coumestrol, and equol in bovine blood plasma and urine. J. Assoc. Off. Anal. Chem. 1988, 71, 938–941. [Google Scholar] [PubMed]
- Setchell, K.D.; Cole, S.J. Method of defining equol-producer status and its frequency among vegetarians. J. Nutr. 2006, 136, 2188–2193. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, S.; Yanagibori, R.; Amano, K. Self-administered diet history questionnaire developed for health education: A relative validation of the test-version by comparison with 3-day diet record in women. J. Epidemiol. 1998, 8, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Okubo, H.; Sasaki, S.; Rafamantanantsoa, H.H.; Ishikawa-Takata, K.; Okazaki, H.; Tabata, I. Validation of self-reported energy intake by a self-administered diet history questionnaire using the doubly labeled water method in 140 Japanese adults. Eur. J. Clin. Nutr. 2008, 62, 1343–1350. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Honda, S.; Murakami, K.; Sasaki, S.; Okubo, H.; Hirota, N.; Notsu, A.; Fukui, M.; Date, C. Both comprehensive and brief self-administered diet history questionnaires satisfactorily rank nutrient intakes in Japanese adults. J. Epidemiol. 2012, 22, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of Bacteria and Archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef] [PubMed]
- Hisada, T.; Endoh, K.; Kuriki, K. Inter- and intra-individual variations in seasonal and daily stabilities of the human gut microbiota in Japanese. Arch. Microbiol. 2015, 197, 919–934. [Google Scholar] [CrossRef] [PubMed]
- Maruo, T.; Sakamoto, M.; Ito, C.; Toda, T.; Benno, Y. Adlercreutzia equolifaciens gen. nov., sp. nov., an equol-producing bacterium isolated from human faeces, and emended description of the genus Eggerthella. Int. J. Syst. Evol. Microbiol. 2008, 58, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Minamida, K.; Ota, K.; Nishimukai, M.; Tanaka, M.; Abe, A.; Sone, T.; Tomita, F.; Hara, H.; Asano, K. Asaccharobacter celatus gen. nov., sp. nov., isolated from rat caecum. Int. J. Syst. Evol. Microbiol. 2008, 58, 1238–1240. [Google Scholar] [CrossRef] [PubMed]
- Ueno, T.; Uchiyama, S. Identification of the specific intestinal bacteria capable of metabolising soy isoflavone to equol. Ann. Nutr. Metab. 2002, 45, 114. [Google Scholar]
- Decroos, K.; Vanhemmens, S.; Cattoir, S.; Boon, N.; Verstraete, W. Isolation and characterization of an equol-producing mixed microbial culture from a human faecal sample and its activity under gastrointestinal conditions. Arch. Microbiol. 2005, 183, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.S.; Kitahara, M.; Sakamoto, M.; Hattori, M.; Benno, Y. Slackia equolifaciens sp. nov., a human intestinal bacterium capable of producing equol. Int. J. Syst. Evol. Microbiol. 2010, 60, 1721–1740. [Google Scholar] [CrossRef] [PubMed]
- Matthies, A.; Blaut, M.; Braune, A. Isolation of a human intestinal bacterium capable of daidzein and genistein conversion. Appl. Environ. Microbiol. 2009, 75, 1740–1744. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, K.; Tanaka, M.; Hirao, Y.; Nagata, Y.; Mori, M.; Miyanaga, N.; Akaza, H.; Kim, W.J. Age-stratified serum levels of isoflavones and proportion of equol producers in Japanese and Korean healthy men. Prostate Cancer Prostatic Dis. 2008, 11, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Ueno, T.; Abiru, Y.; Uchiyama, S.; Ishimi, Y. Distribution of 24-h urinary equol excretion as an indicator of the physiological range in healthy Japanese equol excretors. J. Funct. Foods 2014, 7, 129–135. [Google Scholar] [CrossRef]
- Franke, A.A.; Lai, J.F.; Pagano, I.; Morimoto, Y.; Maskarinec, G. Equol production changes over time in pre-menopausal women. Br. J. Nutr. 2012, 107, 1201–1206. [Google Scholar] [CrossRef] [PubMed]
- Nakatsu, C.H.; Armstrong, A.; Clavijo, A.P.; Martin, B.R.; Barnes, S.; Weaver, C.M. Fecal bacterial community changes associated with isoflavone metabolites in postmenopausal women after soy bar consumption. PLoS ONE 2014, 9, e108924. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Equol Producer | Non-Producer | p-Value | |
|---|---|---|---|
| number | 458 (43.8%) | 586 (56.2%) | |
| Sex, male | 177 (38.6%) | 234 (39.9%) | 0.673 |
| Age (year) | 56.8 ± 14.7 | 52.9 ± 15.0 | <0.001 |
| Body mass index (kg/m2) | 22.90 ± 3.69 | 22.56 ± 4.19 | 0.167 |
| Intake of daidzein (mg/day) | 15.0 (13.3) | 12.8 (12.3) | 0.004 |
| Urine equol concentration (μmol) | 6.49 (14.93) | 0.04 (0.05) | <0.001 |
| Urine daidzein concentration (μmol) | 5.95 (14.68) | 12.12 (23.43) | <0.001 |
| log (Urine equol/daidzein) | −0.03 (1.17) | −2.45 (0.69) | <0.001 |
| Equol Producer (n = 458) | Non-Producer (n = 586) | p Value | WAD | Rank | |
|---|---|---|---|---|---|
| Adlercreutzia equolifaciens | 4.05 ± 32.16 | 3.78 ± 29.16 | 0.121 | 0.000 | 6 |
| Asaccharobacter celatus | 1.71 ± 3.52 | 0.37 ± 14.56 | <0.001 | 0.094 | 3 |
| Bacteroides ovatus | 57.34 ± 112.37 | 117.78 ± 219.26 | 0.058 | 0.180 | 2 |
| Finegoldia magna | 0.27 ± 1.76 | 0.30 ± 2.04 | 0.124 | 0.001 | 5 |
| Lactobacillus mucosae | 3.75 ± 25.66 | 8.90 ± 83.56 | 0.481 | 0.011 | 4 |
| Slackia equolifaciens | 0.01 ± 0.06 | 0 | 0.808 | 0 | 8 |
| Slackia isoflavoniconvertens | 6.87 ± 15.06 | 0.72 ± 0.53 | <0.001 | 0.264 | 1 |
| Streptococcus intermedius | 0.04 ± 0.17 | 0.06 ± 0.03 | 0.860 | 0.000 | 7 |
| Equol Producer | Non-Producer | p | |||
|---|---|---|---|---|---|
| Adlercreutzia equolifaciens | 51 | (11.1) | 31 | (5.3) | <0.001 |
| Asaccharobacter celatus | 230 | (50.2) | 69 | (11.8) | <0.001 |
| Bacteroides ovatus | 406 | (88.6) | 445 | (75.9) | <0.001 |
| Finegoldia magna | 68 | (14.8) | 120 | (20.5) | 0.019 |
| Lactobacillus mucosae | 56 | (12.2) | 86 | (14.7) | 0.252 |
| Slackia equolifaciens | 4 | (0.8) | 0 | (0) | 0.023 |
| Slackia isoflavoniconvertens | 178 | (38.9) | 25 | (4.3) | <0.001 |
| Streptococcus intermedius | 34 | (7.4) | 47 | (8.0) | 0.721 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iino, C.; Shimoyama, T.; Iino, K.; Yokoyama, Y.; Chinda, D.; Sakuraba, H.; Fukuda, S.; Nakaji, S. Daidzein Intake Is Associated with Equol Producing Status through an Increase in the Intestinal Bacteria Responsible for Equol Production. Nutrients 2019, 11, 433. https://doi.org/10.3390/nu11020433
Iino C, Shimoyama T, Iino K, Yokoyama Y, Chinda D, Sakuraba H, Fukuda S, Nakaji S. Daidzein Intake Is Associated with Equol Producing Status through an Increase in the Intestinal Bacteria Responsible for Equol Production. Nutrients. 2019; 11(2):433. https://doi.org/10.3390/nu11020433
Chicago/Turabian StyleIino, Chikara, Tadashi Shimoyama, Kaori Iino, Yoshihito Yokoyama, Daisuke Chinda, Hirotake Sakuraba, Shinsaku Fukuda, and Shigeyuki Nakaji. 2019. "Daidzein Intake Is Associated with Equol Producing Status through an Increase in the Intestinal Bacteria Responsible for Equol Production" Nutrients 11, no. 2: 433. https://doi.org/10.3390/nu11020433
APA StyleIino, C., Shimoyama, T., Iino, K., Yokoyama, Y., Chinda, D., Sakuraba, H., Fukuda, S., & Nakaji, S. (2019). Daidzein Intake Is Associated with Equol Producing Status through an Increase in the Intestinal Bacteria Responsible for Equol Production. Nutrients, 11(2), 433. https://doi.org/10.3390/nu11020433