A Comprehensive Assessment of Apigenin as an Antiproliferative, Proapoptotic, Antiangiogenic and Immunomodulatory Phytocompound

 , ,
, ,  ,
,  ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Preparation
2.2. Cell Viability Assays
2.3. Cell Cycle Analysis by Flow Cytometry
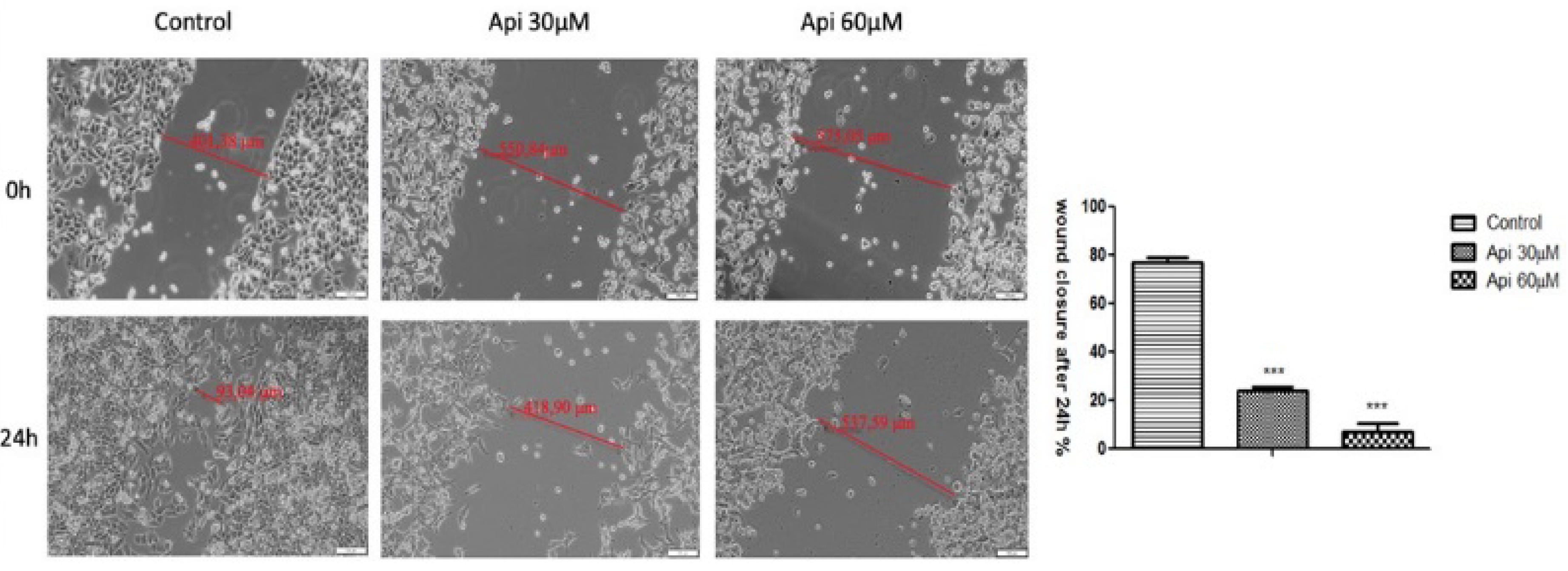
2.4. Anti-Migratory Potential—Scratch Assay Method
2.5. Determination of In Situ Caspase Activity
2.6. Annexin V—Propidium Iodide Assay
2.7. Assessment of Cytotoxicity via LDH Released
2.8. Fluorescence Microscopy
2.9. Extracellular Flux (XF) Analysis
2.10. Chorioallantoic Membrane Assay (CAM)
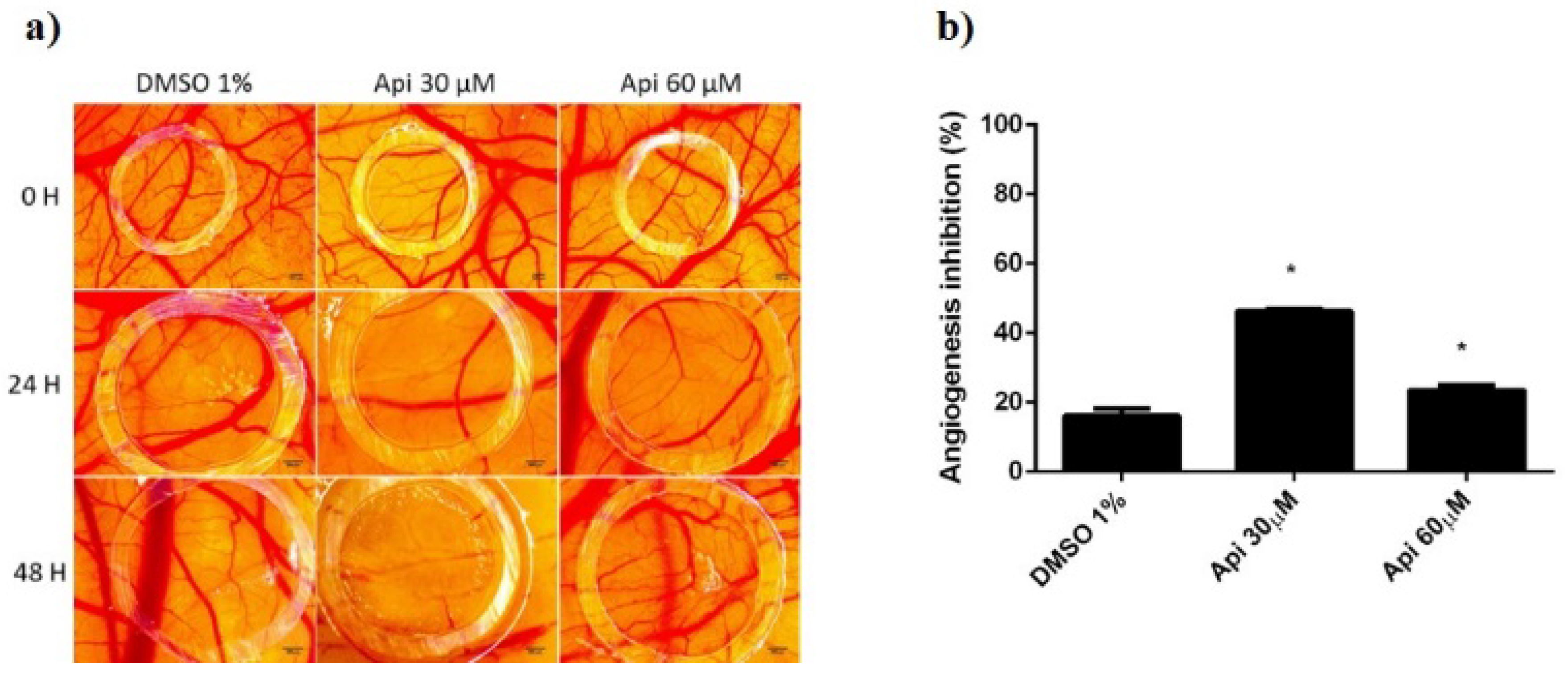
2.11. Normal Angiogenesis Assessment on the Chorioallantoic Membrane
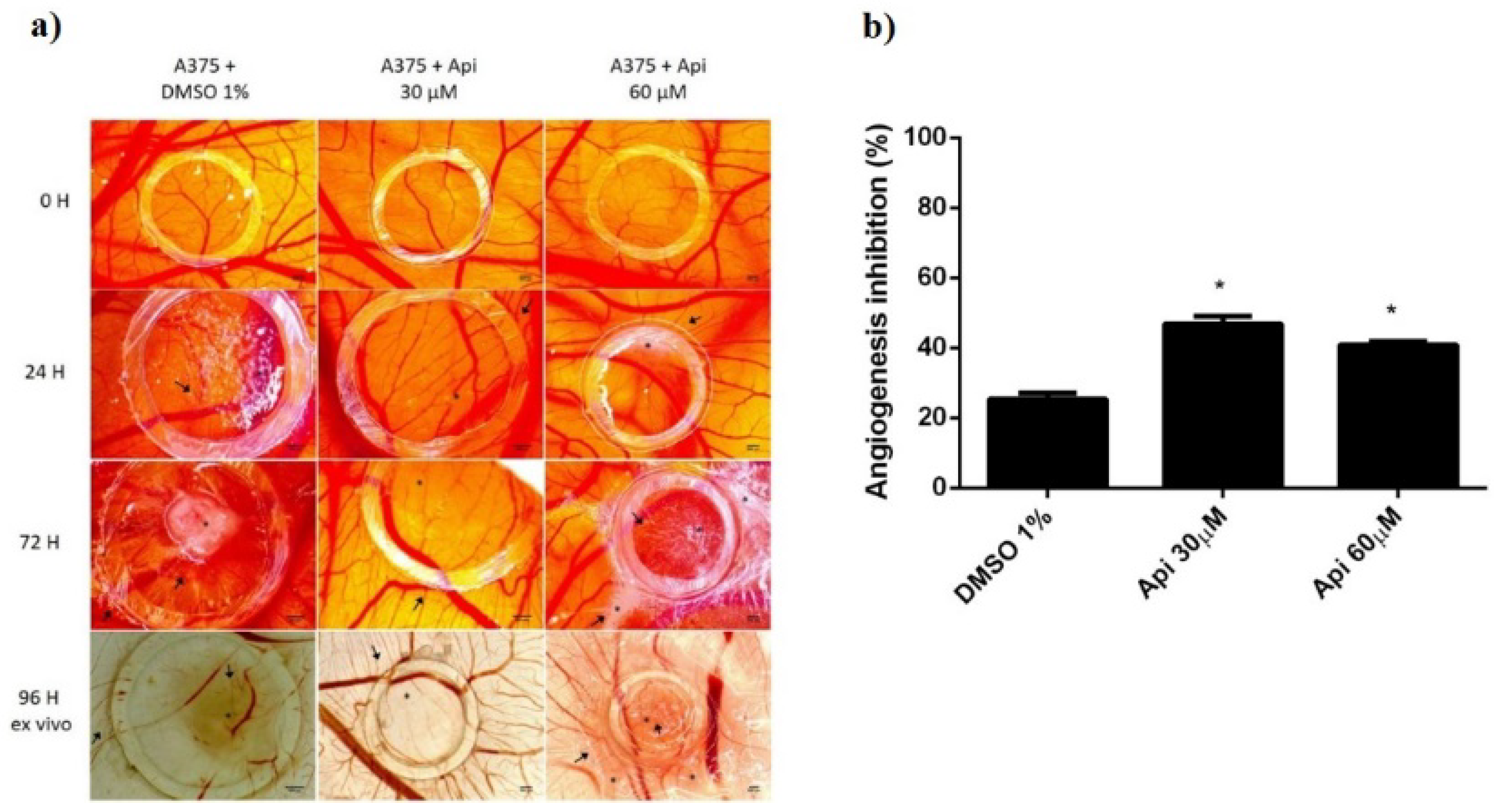
2.12. Tumour Angiogenesis Assessment on the Chorioallantoic Membrane
2.13. Statistics
3. Results
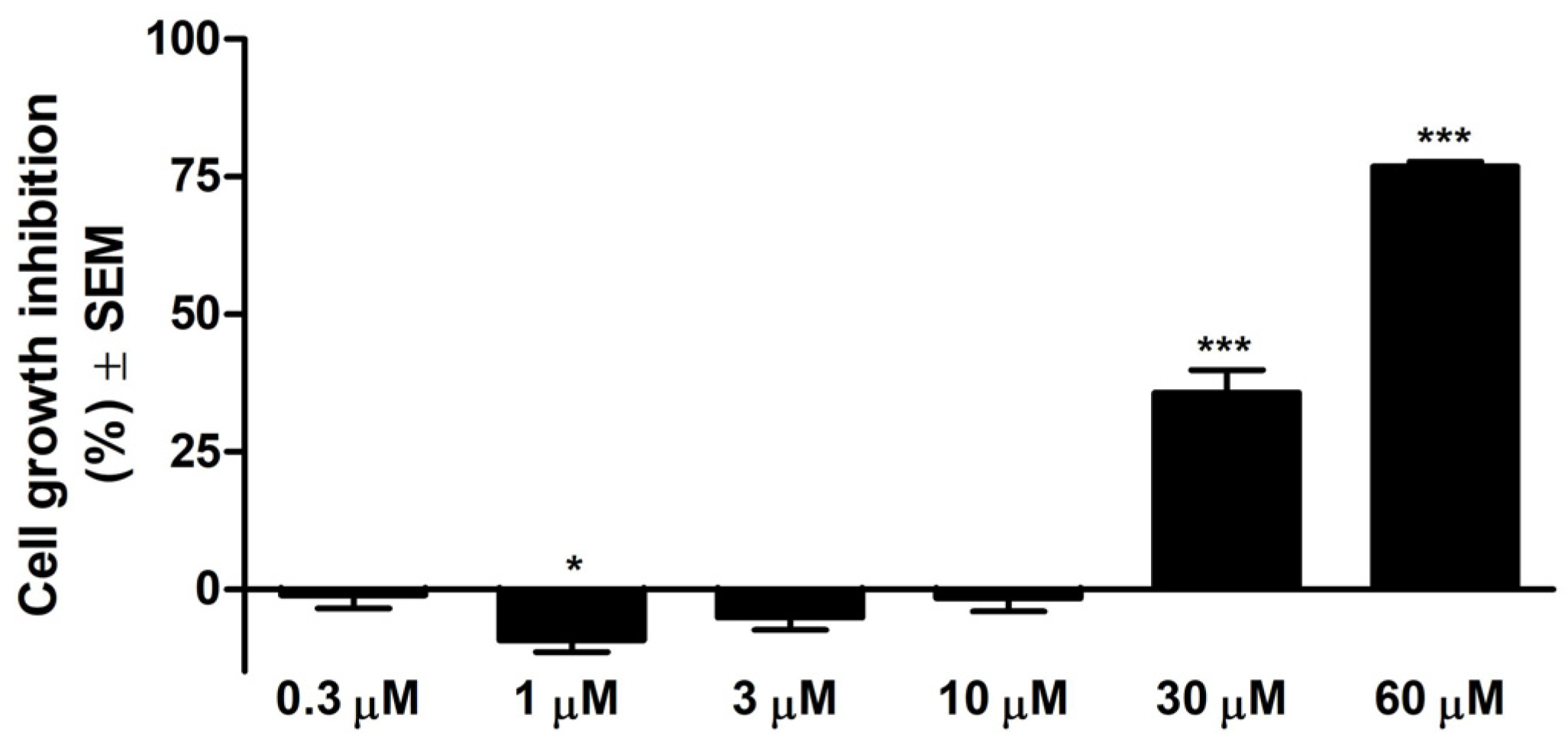
3.1. Cell Growth Inhibition
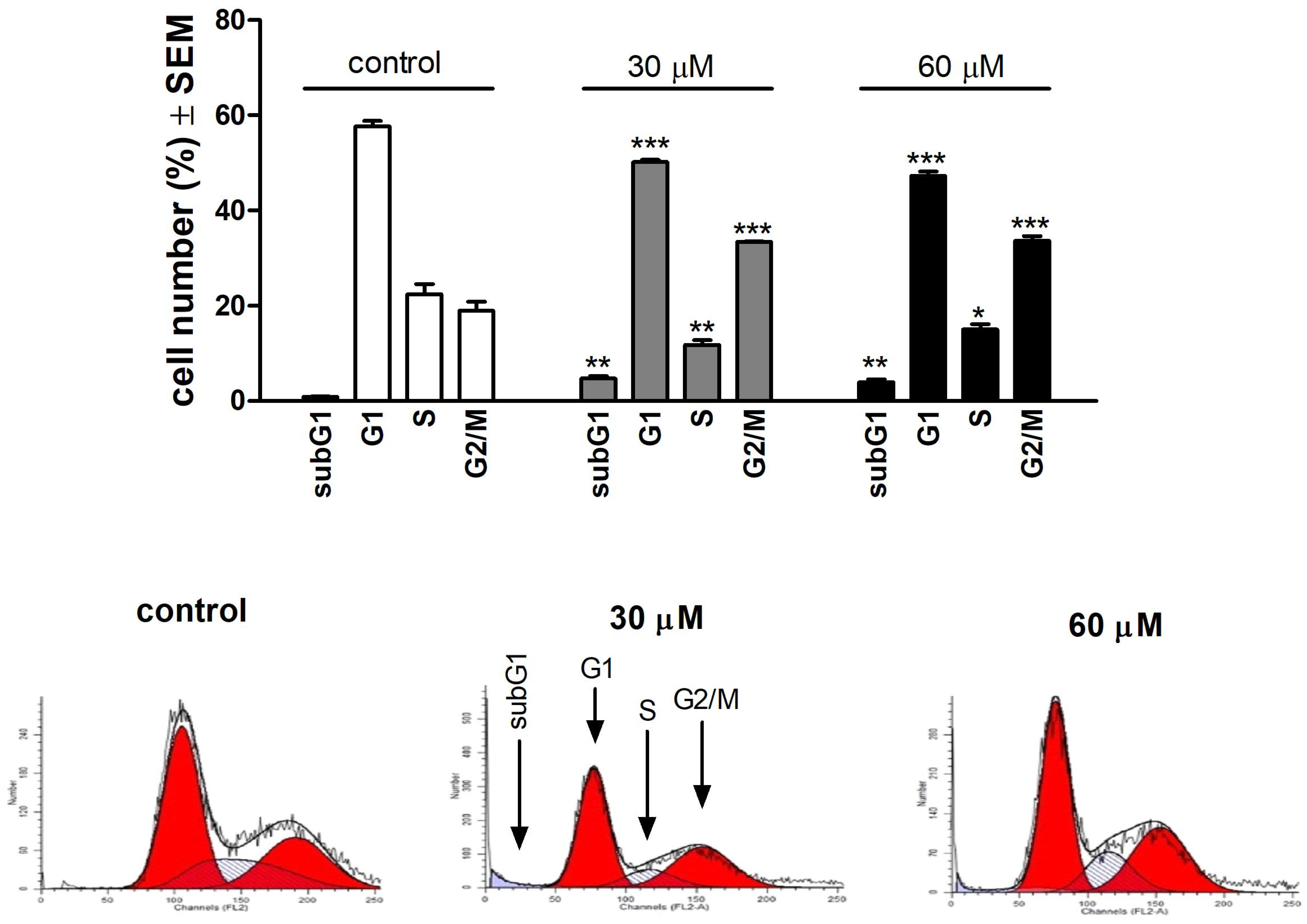
3.2. Api Effects on Cell Cycle Phases
3.3. Antiproliferative Effect of Api
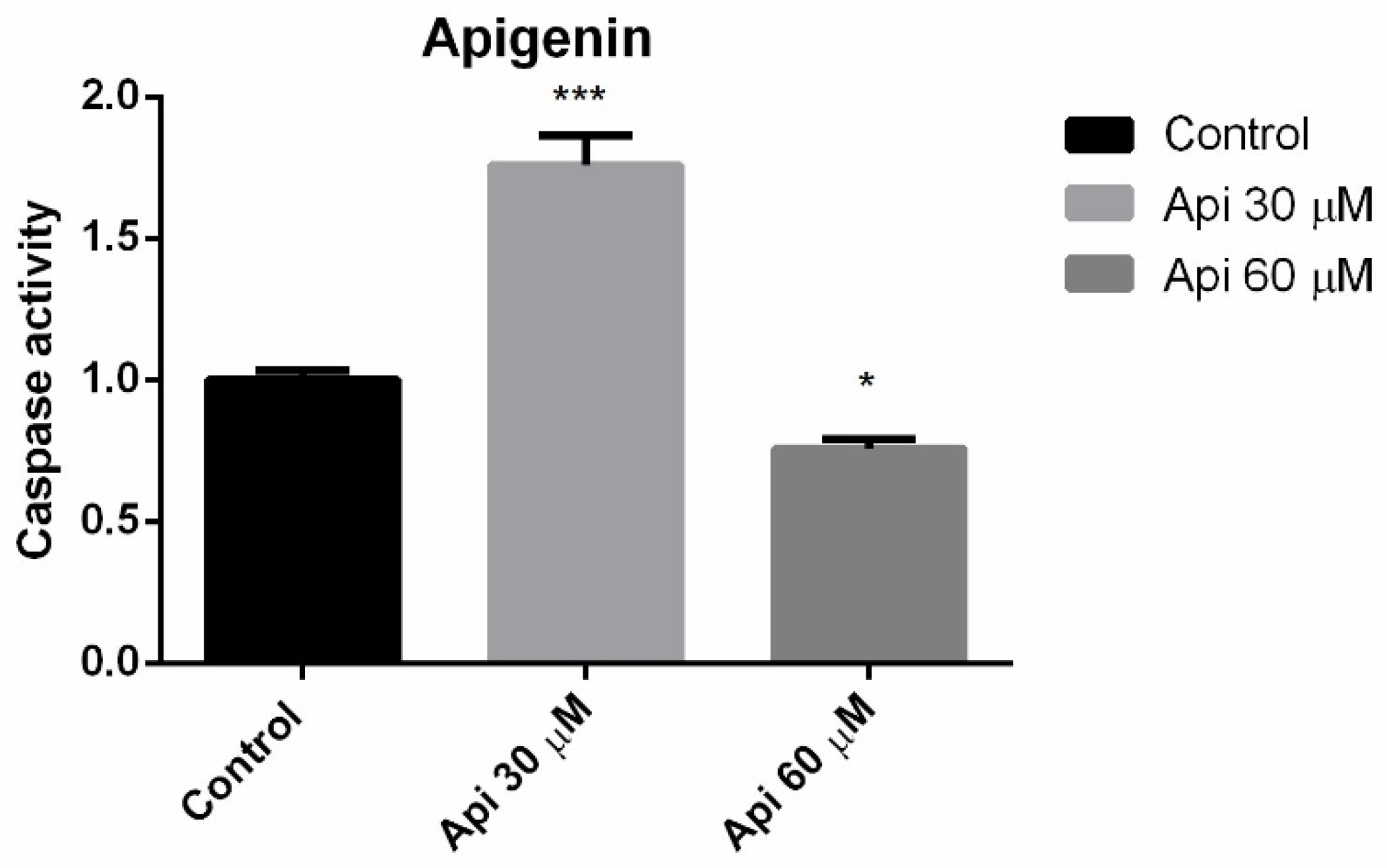
3.4. Caspase 3 Activity
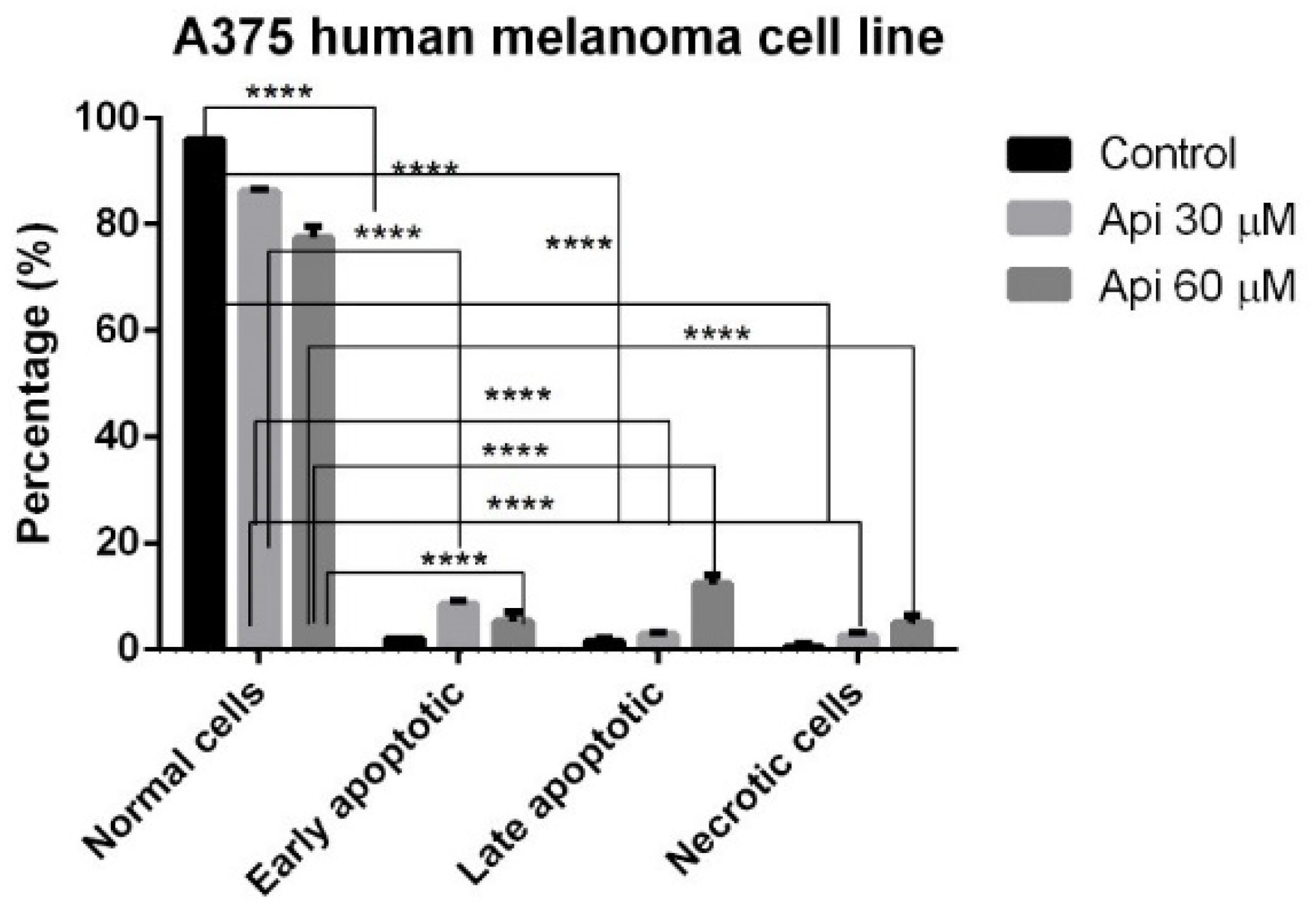
3.5. Annexin V-PI
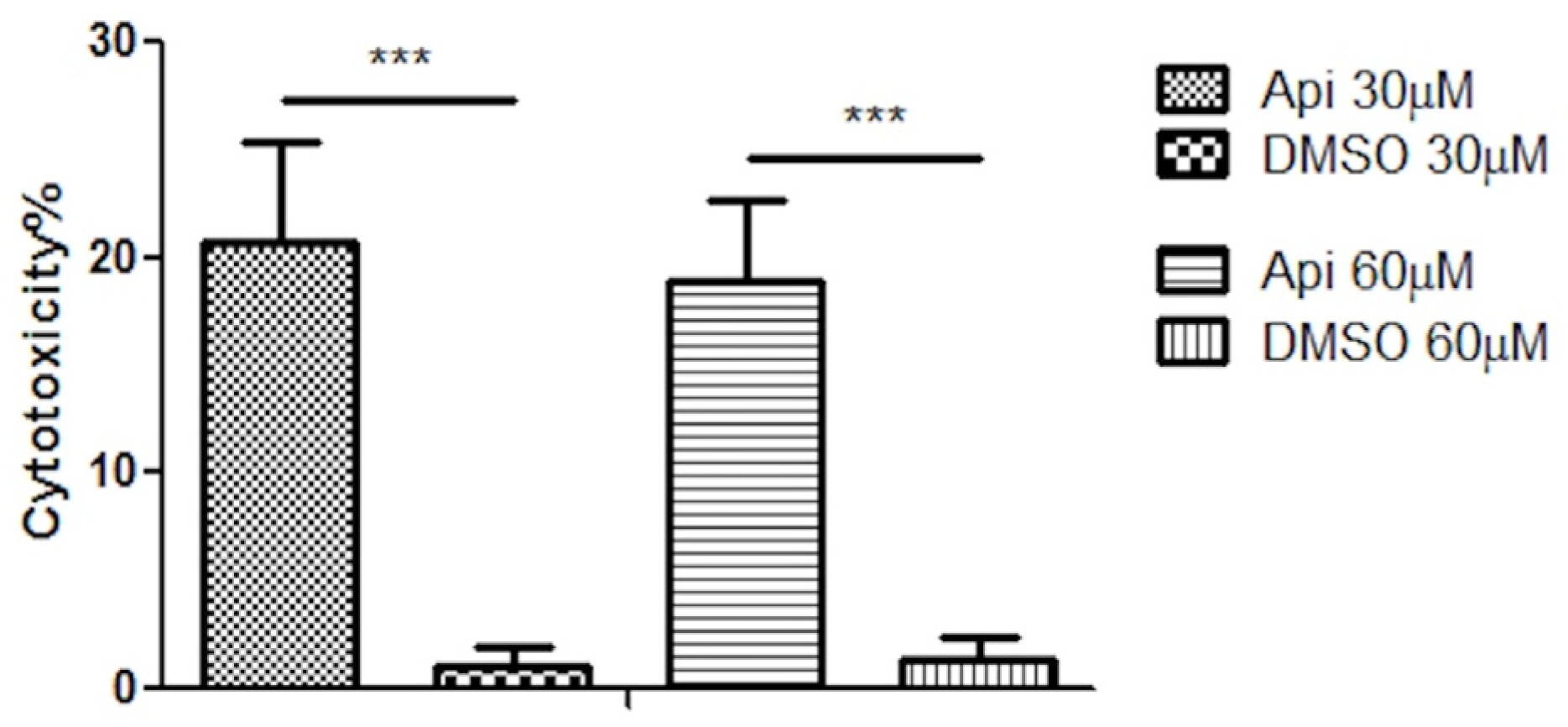
3.6. LDH Assay
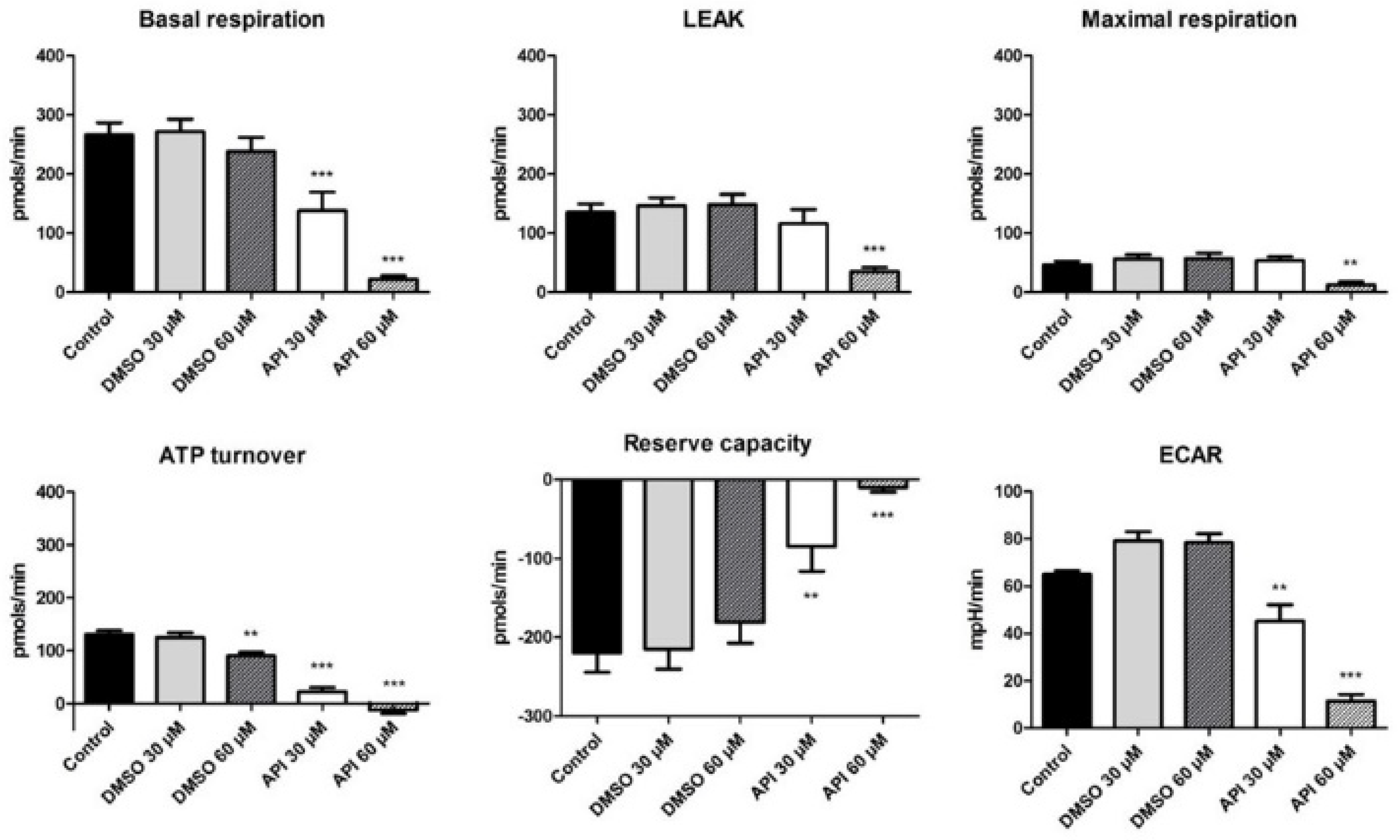
3.7. Bioenergetic Profile of A375 Human Melanoma Cells
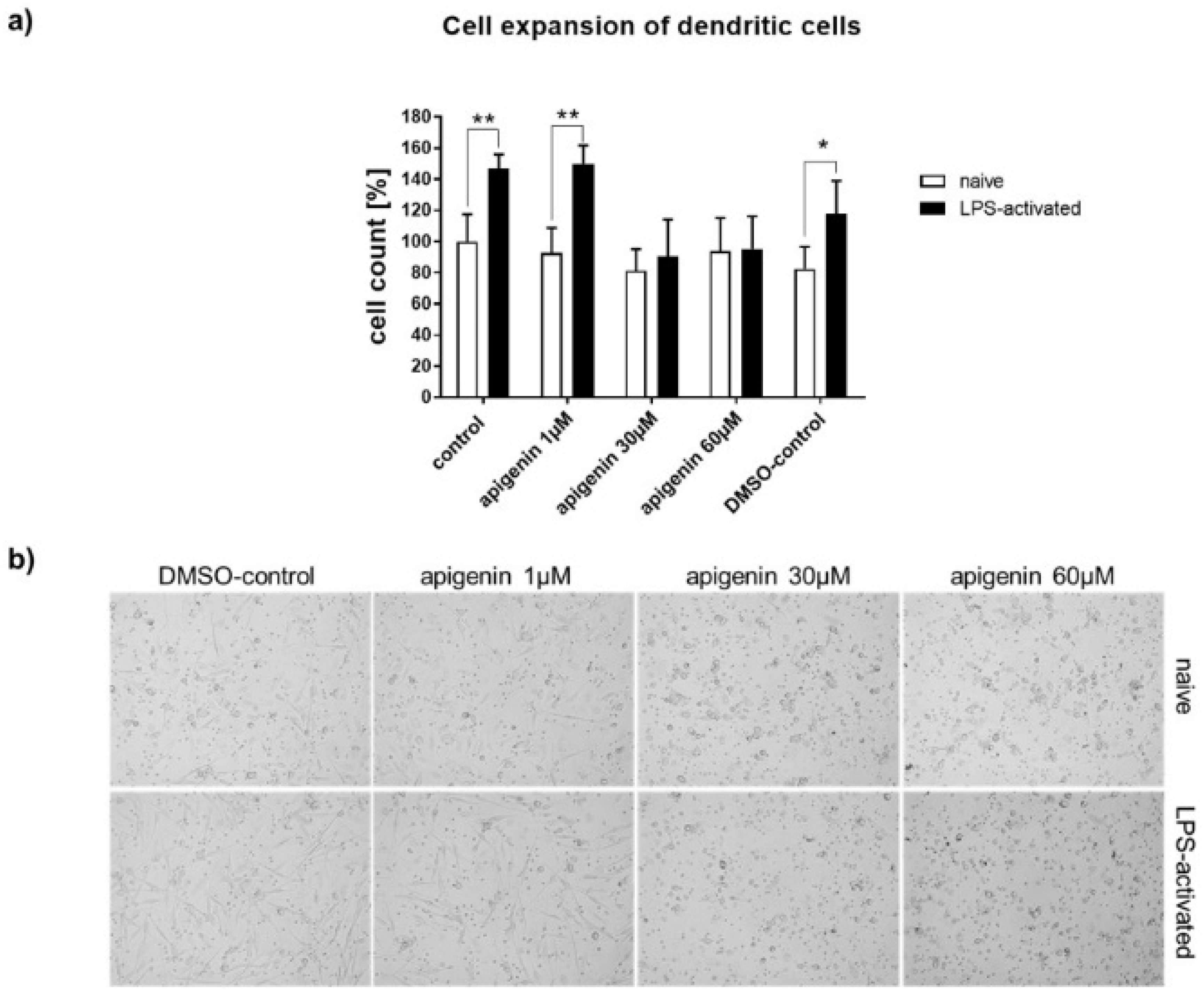
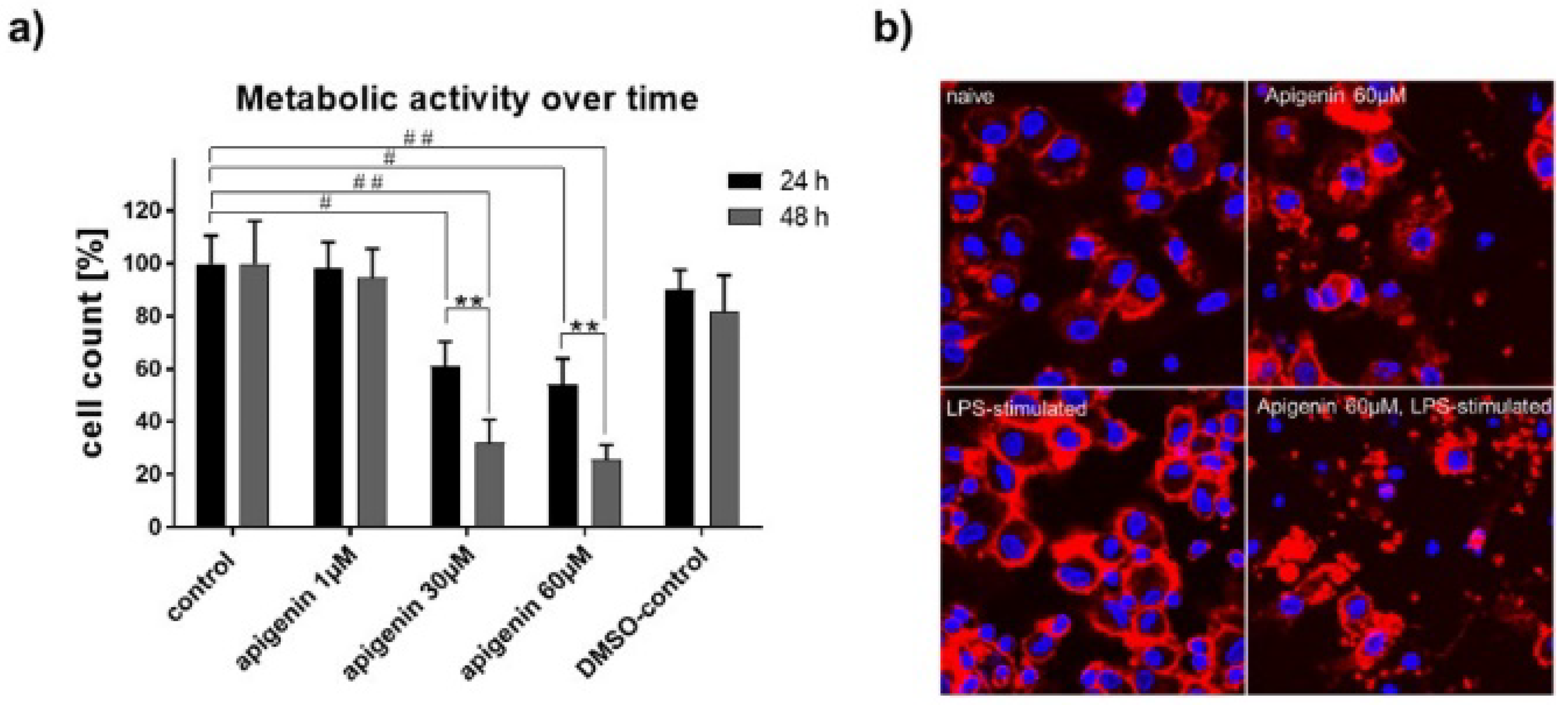
3.8. Immunomodulatory Effects of Api
3.9. XTT Assay for Metabolic Activity
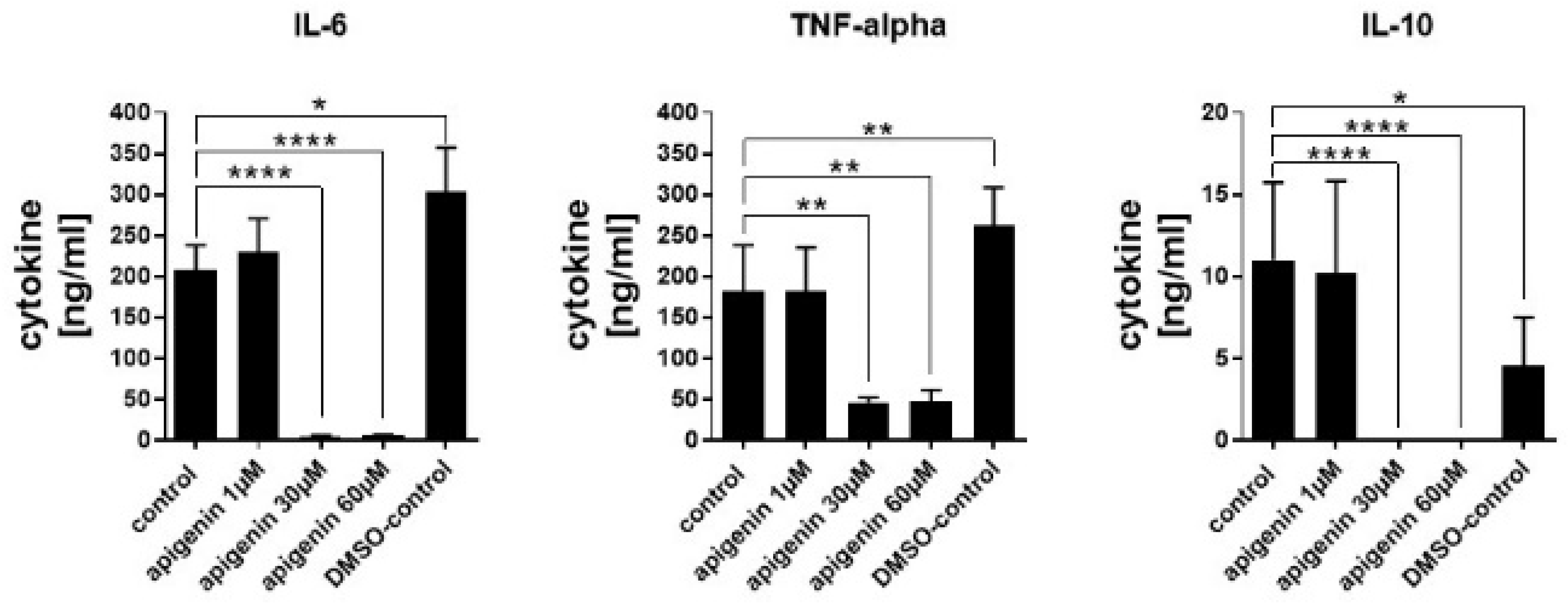
3.10. Cytokine Quantification
3.11. Chorioallantoic Membrane Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nobili, S.; Lippi, D.; Witort, E.; Donnini, M.; Bausi, L.; Mini, E.; Capaccioli, S. Natural Compounds for Cancer Treatment and Prevention. Pharmacol. Res. 2009, 59, 365–378. [Google Scholar] [CrossRef] [PubMed]
- Hallberg, Ö.; Johansson, O. Increasing Melanoma—Too Many Skin Cell Damages or Too Few Repairs? Cancers 2013, 5, 184–204. [Google Scholar] [CrossRef] [PubMed]
- HHS. Available online: https://www.hhs.gov/ (accessed on 1 February 2019).
- Shain, A.H.; Bastian, B.C. From Melanocytes to Melanomas. Nat. Rev. Cancer 2016, 16, 345–358. [Google Scholar] [CrossRef]
- Chahar, M.K.; Sharma, N.; Dobhal, M.P.; Joshi, Y.C. Flavonoids: A Versatile Source of Anticancer Drugs. Pharmacogn. Rev. 2011, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J.; Kim, G.H. Apigenin Induces Apoptosis through a Mitochondria/Caspase-Pathway in Human Breast Cancer MDA-MB-453 Cells. J. Clin. Biochem. Nutr. 2009, 44, 260–265. [Google Scholar] [CrossRef] [Green Version]
- Moga, M.A.; Dimienescu, O.G.; Arvatescu, C.A.; Mironescu, A.; Dracea, L.; Ples, L. The Role of Natural Polyphenols in the Prevention and Treatment of Cervical Cancer-An Overview. Molecules 2016, 21, 1055. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, F.; Zhou, R.; Song, X.; Xie, M. Apigenin: A Current Review on its beneficial Biological Activities. J. Food Biochem. 2017, 41, e12376. [Google Scholar] [CrossRef]
- Venigalla, M.; Gyengesi, E.; Münch, G. Curcumin and Apigenin—Novel and Promising Therapeutics Against Chronic Neuroinflammation in Alzheimer′s Disease. Neural Regen. Res. 2015, 10, 1181–1185. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.; Kanwal, R.; Bhaskaran, N.; Gupta, S. Plant Flavone Apigenin Binds to Nucleic Acid Bases and Reduces Oxidative DNA Damage in Prostate Epithelial Cells. PLoS ONE 2014, 9, e91588. [Google Scholar] [CrossRef]
- Yan, X.; Qi, M.; Li, P.; Zhan, Y.; Shao, H. Apigenin in Cancer Therapy: Anti-cancer Effects and Mechanisms of Action. Cell Biosci. 2017, 7, 50. [Google Scholar] [CrossRef]
- Madunić, J.; Madunić, I.V.; Gajski, G.; Popić, J.; Garaj-Vrhovac, V. Apigenin: A Dietary Flavonoid with Diverse Anticancer Properties. Cancer Lett. 2018, 413, 11–22. [Google Scholar] [CrossRef]
- Rakoff-Nahoum, S. Why Cancer and Inflammation? Yale J. Biol. Med. 2006, 79, 123–130. [Google Scholar]
- Perrott, K.M.; Wiley, C.D.; Desprez, P.Y.; Campisi, J. Apigenin Suppresses the Senescence-Associated Secretory Phenotype and Paracrine Effects on Breast Cancer Cells. GeroScience 2017, 39, 161–173. [Google Scholar] [CrossRef]
- Zhao, G.; Han, X.; Cheng, W.; Ni, J.; Zhang, Y.; Lin, J.; Song, Z. Apigenin Inhibits Proliferation and Invasion, and Induces Apoptosis and Cell Cycle Arrest in Human Melanoma Cells. Oncol. Rep. 2017, 37, 2277–2285. [Google Scholar] [CrossRef] [PubMed]
- Caltagirone, S.; Rossi, C.; Poggi, A.; Ranelletti, F.O.; Natali, P.G.; Brunetti, M.; Aiello, F.B.; Piantelli, M. Flavonoids Apigenin and Quercetin Inhibit Melanoma Growth and Metastatic Potential. Int. J. Cancer 2000, 87, 595–600. [Google Scholar] [CrossRef]
- Piantelli, M.; Rossi, C.; Iezzi, M.; La Sorda, R.; Iacobelli, S.; Alberti, S.; Natali, P.G. Flavonoids Inhibit Melanoma Lung Metastasis by Impairing Tumour Cells Endothelium Interactions. J. Cell Physiol. 2006, 207, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.H.; Chu, J.H.; Kwan, H.Y.; Su, T.; Yu, H.; Cheng, C.Y.; Fu, X.Q.; Guo, H.; Li, T.; Tse, A.K.; et al. Inhibition of the STAT3 signaling pathway contributes to Apigenin-Mediated Anti-Metastatic Effect in Melanoma. Sci. Rep. 2016, 6, 21731. [Google Scholar] [CrossRef] [PubMed]
- Schwiebs, A.; Thomas, D.; Kleuser, B.; Pfeilschifter, J.M.; Radeke, H.H. Nuclear Translocation of SGPP-1 and Decrease of SGPL-1 Activity Contribute to Sphingolipid Rheostat Regulation of Inflammatory Dendritic Cells. Mediat. Inflamm. 2017, 2017, 5187368. [Google Scholar] [CrossRef]
- Danciu, C.; Zupko, I.; Bor, A.; Schwiebs, A.; Radeke, H.; Hancianu, M.; Cioanca, O.; Alexa, E.; Oprean, C.; Bojin, F.; et al. Botanical Therapeutics: Phytochemical Screening and Biological Assessment of Chamomile, Parsley and Celery Extracts against A375 Human Melanoma and Dendritic Cells. Int. J. Mol. Sci. 2018, 19, 3624. [Google Scholar] [CrossRef]
- Danciu, C.; Muntean, D.; Alexa, E.; Farcas, C.; Oprean, C.; Zupko, I.; Bor, A.; Minda, D.; Proks, M.; Buda, V.; et al. Phytochemical Characterization and Evaluation of the Antimicrobial, Antiproliferative and Proapoptotic Potential of Ephedra Alata Decne. Hydroalcoholic Extract against the MCF-7 Breast Cancer Cell Line. Molecules 2018, 24, 13. [Google Scholar] [CrossRef]
- Duicu, O.M.; Scurtu, I.; Popescu, R.; Sturza, A.; Coricovac, D.; Dănilă, M.D.; Privistirescu, A.; Muntean, D.M. Assessment of the Effects of Methylene Blue on Cellular Bioenergetics in H9c2 Cells. Rev. Chim. 2015, 66, 519–522. [Google Scholar]
- Zuo, Y.H.; Han, Q.B.; Dong, G.T.; Yue, R.Q.; Ren, X.C.; Liu, J.X.; Liu, L.; Luo, P.; Zhou, H. Panax ginseng Polysaccharide Protected H9c2 Cardiomyocyte from Hypoxia/Reoxygenation Injury through Regulating Mitochondrial Metabolism and RISK Pathway. Front. Physiol. 2018, 9, 699. [Google Scholar] [CrossRef] [PubMed]
- Yadava, N.; Nicholls, D.G. Spare Respiratory Capacity Rather than Oxidative Stress Regulates Glutamate Excitotoxicity after Partial Respiratory Inhibition of Mitochondrial Complex I with Rotenone. J. Neurosci. 2007, 27, 7310–7317. [Google Scholar] [CrossRef]
- Ribatti, D.; Vacca, A.; Roncali, L.; Dammacco, F. The Chick Embryo Chorioallantoic Membrane as a Model for In Vivo Research on Anti-Angiogenesis. Curr. Pharm. Biotechnol. 2000, 1, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. The Chick Embryo Chorioallantoic Membrane in the Study of Tumour Angiogenesis. Rom. J. Morphol. Embryol. 2008, 49, 131–135. [Google Scholar] [PubMed]
- Demir, R.; Peros, G.; Hohenberger, W. Definition of the “Drug-Angiogenic-Activity-Index” that Allows the Quantification of the Positive and Negative Angiogenic Active Drugs: A Study Based on the Chorioallantoic Membrane Model. Pathol. Oncol. Res. 2011, 17, 309–313. [Google Scholar] [CrossRef]
- Özcetin, A.; Aigner, A.; Bakowsky, U. A Chorioallantoic Membrane Model for the Determination of Antiangiogenic Effects of Imatinib. Eur. J. Pharm. Biopharm. 2013, 85, 711–715. [Google Scholar] [CrossRef]
- Zhong, L.; Guo, X.N.; Zhang, X.H.; Sun, Q.M.; Tong, L.J.; Wu, Z.X.; Luo, X.M.; Jiang, H.L.; Nan, F.J.; Zhang, X.W.; et al. TKI-31 Inhibits Angiogenesis by Combined Suppression Signaling Pathway of VEGFR2 and PDGFRbeta. Cancer Biol. Ther. 2006, 5, 323–330. [Google Scholar] [CrossRef]
- Avram, S.; Coricovac, D.E.; Pavel, I.Z.; Pinzaru, I.; Ghiulai, R.; Baderca, F.; Soica, C.; Muntean, D.; Branisteanu, D.E.; Spandidos, D.A.; et al. Standardization of A375 human melanoma models on Chicken Embryo Chorioallantoic Membrane and Balb/c Nude Mice. Oncol. Rep. 2017, 38, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.P.; Chou, T.H.; Ding, H.Y.; Chen, P.R.; Chiang, F.Y.; Kuo, P.L.; Liang, C.H. Apigenin Induces Apoptosis via Tumour Necrosis Factor Receptor- and Bcl-2-Mediated Pathway and Enhances Susceptibility of Head and Neck Squamous Cell Carcinoma to 5-Fluorouracil and Cisplatin. Biochim. Biophys. Acta 2012, 1820, 1081–1091. [Google Scholar] [CrossRef]
- Hasnat, M.A.; Pervin, M.; Lim, J.H.; Lim, B.O. Apigenin Attenuates Melanoma Cell Migration by Inducing Anoikis through Integrin and Focal Adhesion Kinase Inhibition. Molecules 2015, 20, 21157–21166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, F.; Moriwaki, K.; De Rosa, M. Detection of Necrosis by Release of Lactate Dehydrogenase Activity. In Immune Homeostasis: Methods in Molecular Biology (Methods and Protocols); Snow, A., Lenardo, M., Eds.; Humana Press: Totowa, NJ, USA, 2013; Volume 979, pp. 65–70. [Google Scholar]
- Renz, A.; Berdel, W.E.; Kreuter, M.; Belka, C.; Schulze-Osthoff, K.; Los, M. Rapid Extracellular Release of Cytochrome c Is Specific for Apoptosis and Marks Cell Death In Vivo. Blood 2001, 98, 1542–1548. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Wunder, M.B.; Norris, D.A.; Shellman, Y.G. A Simple Protocol for Using a LDH-Based Cytotoxicity Assay to Assess the Effects of Death and Growth Inhibition at the Same Time. PLoS ONE 2011, 6, e26908. [Google Scholar] [CrossRef] [PubMed]
- Chao, S.C.; Huang, S.C.; Hu, D.N.; Lin, H.Y. Subtoxic Levels of Apigenin Inhibit Expression and Secretion of VEGF by Uveal Melanoma Cells via Suppression of ERK1/2 and PI3K/Akt Pathways. Evid.-Based Complement Altern. Med. 2013, 2013, 817674. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Das, J.; Samadder, A.; Boujedaini, N.; Khuda-Bukhsh, A.R. Apigenin-Induced Apoptosis in A375 and A549 Cells through Selective Action and Dysfunction of Mitochondria. Exp. Biol. Med. 2012, 237, 1433–1448. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.; Staub, R.E.; Baggett, S.; Chimmani, R.; Tagliaferri, M.; Cohen, I.; Shtivelman, E. Identification and Analysis of the Active Phytochemicals from the anti-Cancer Botanical Extract Bezielle. PLoS ONE 2012, 7, e30107. [Google Scholar] [CrossRef] [PubMed]
- Seydi, E.; Rasekh, H.R.; Salimi, A.; Mohsenifar, Z.; Pourahmad, J. Selective Toxicity of Apigenin on Cancerous Hepatocytes by Directly Targeting their Mitochondria. Anticancer Agents Med. Chem. 2016, 16, 1576–1586. [Google Scholar] [CrossRef]
- Salmani, J.M.M.; Zhang, X.P.; Jacob, J.A.; Chen, B.A. Apigenin’s anticancer properties And Molecular Mechanisms of Action: Recent Advances and Future Prospectives. Chin. J. Nat. Med. 2017, 15, 321–329. [Google Scholar] [CrossRef]
- Nicholls, D.G.; Darley-Usmar, V.M.; Wu, M.; Jensen, P.B.; Rogers, G.W.; Ferrick, D.A. Bioenergetic Profile Experiment using C2C12 Myoblast Cells. J. Vis. Exp. 2010, 46, e2511. [Google Scholar] [CrossRef]
- Nicholson, L.B. The Immune System. Essays Biochem. 2016, 60, 275–301. [Google Scholar] [CrossRef]
- Spel, L.; Boelens, J.J.; Nierkens, S.; Boes, M. Antitumor Immune Responses Mediated by Dendritic Cells: How Signals Derived from Dying Cancer Cells Drive Antigen Cross-presentation. Oncoimmunology 2013, 2, e26403. [Google Scholar] [CrossRef] [PubMed]
- Shankar, E.; Goel, A.; Gupta, K.; Gupta, S. Plant Flavone Apigenin: An Emerging Anticancer Agent. Curr. Pharmacol. Rep. 2017, 3, 423–446. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Zhou, Q.; Liu, L.Z.; Xia, C.; Hu, X.; Shi, X.; Jiang, B.H. Apigenin Inhibits Tumour Angiogenesis through Decreasing HIF-1alpha and VEGF Expression. Carcinogenesis 2007, 28, 858–864. [Google Scholar] [CrossRef] [PubMed]












© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghițu, A.; Schwiebs, A.; Radeke, H.H.; Avram, S.; Zupko, I.; Bor, A.; Pavel, I.Z.; Dehelean, C.A.; Oprean, C.; Bojin, F.; et al. A Comprehensive Assessment of Apigenin as an Antiproliferative, Proapoptotic, Antiangiogenic and Immunomodulatory Phytocompound. Nutrients 2019, 11, 858. https://doi.org/10.3390/nu11040858
Ghițu A, Schwiebs A, Radeke HH, Avram S, Zupko I, Bor A, Pavel IZ, Dehelean CA, Oprean C, Bojin F, et al. A Comprehensive Assessment of Apigenin as an Antiproliferative, Proapoptotic, Antiangiogenic and Immunomodulatory Phytocompound. Nutrients. 2019; 11(4):858. https://doi.org/10.3390/nu11040858
Chicago/Turabian StyleGhițu, Alexandra, Anja Schwiebs, Heinfried H. Radeke, Stefana Avram, Istvan Zupko, Andrea Bor, Ioana Zinuca Pavel, Cristina Adriana Dehelean, Camelia Oprean, Florina Bojin, and et al. 2019. "A Comprehensive Assessment of Apigenin as an Antiproliferative, Proapoptotic, Antiangiogenic and Immunomodulatory Phytocompound" Nutrients 11, no. 4: 858. https://doi.org/10.3390/nu11040858
APA StyleGhițu, A., Schwiebs, A., Radeke, H. H., Avram, S., Zupko, I., Bor, A., Pavel, I. Z., Dehelean, C. A., Oprean, C., Bojin, F., Farcas, C., Soica, C., Duicu, O., & Danciu, C. (2019). A Comprehensive Assessment of Apigenin as an Antiproliferative, Proapoptotic, Antiangiogenic and Immunomodulatory Phytocompound. Nutrients, 11(4), 858. https://doi.org/10.3390/nu11040858