Extract Methods, Molecular Characteristics, and Bioactivities of Polysaccharide from Alfalfa (Medicago sativa L.)


 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
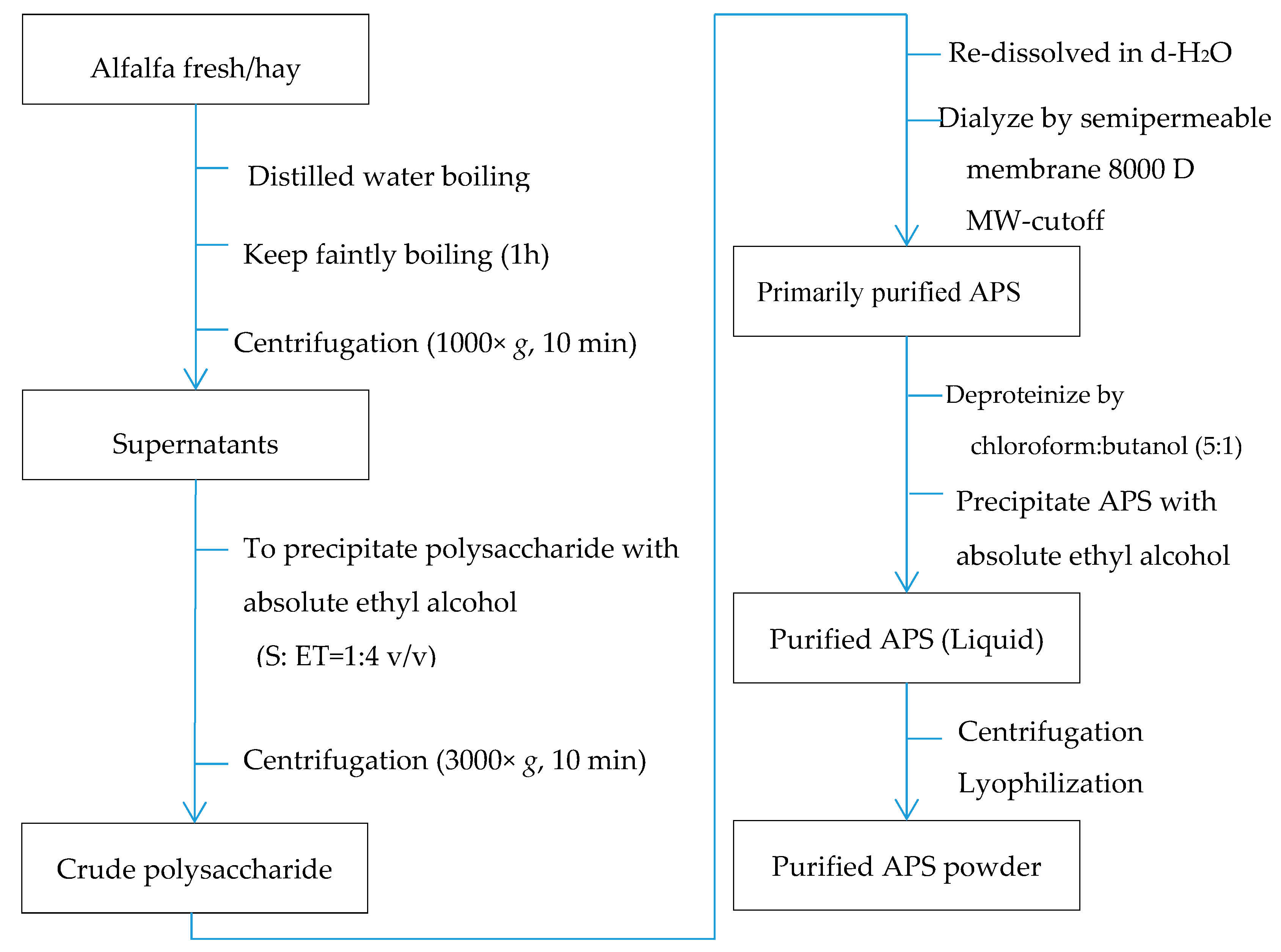
2.1. The Extraction and Purification of Polysaccharides from Alfalfa
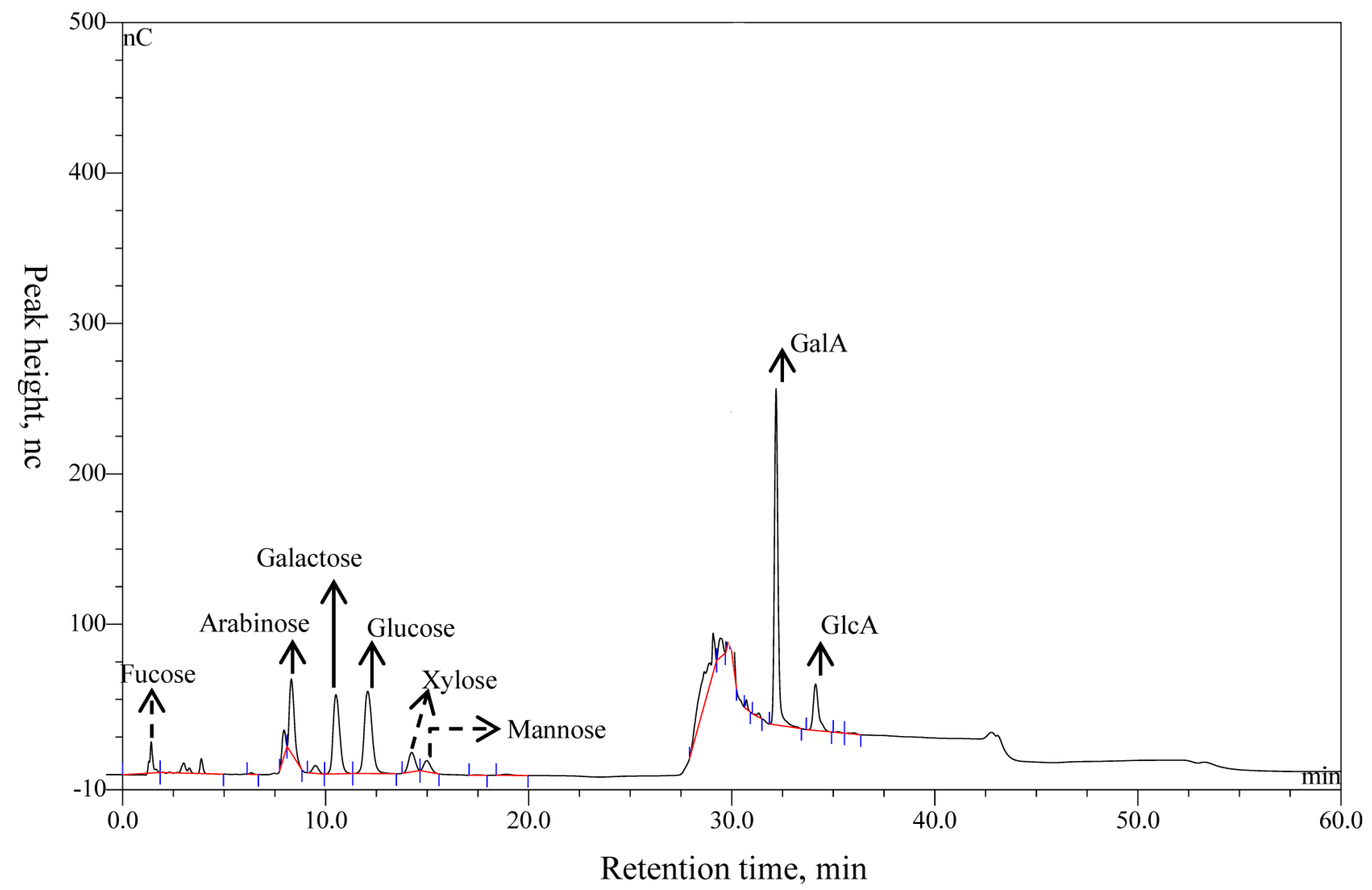
2.2. Determination of Monosaccharide Components of APS
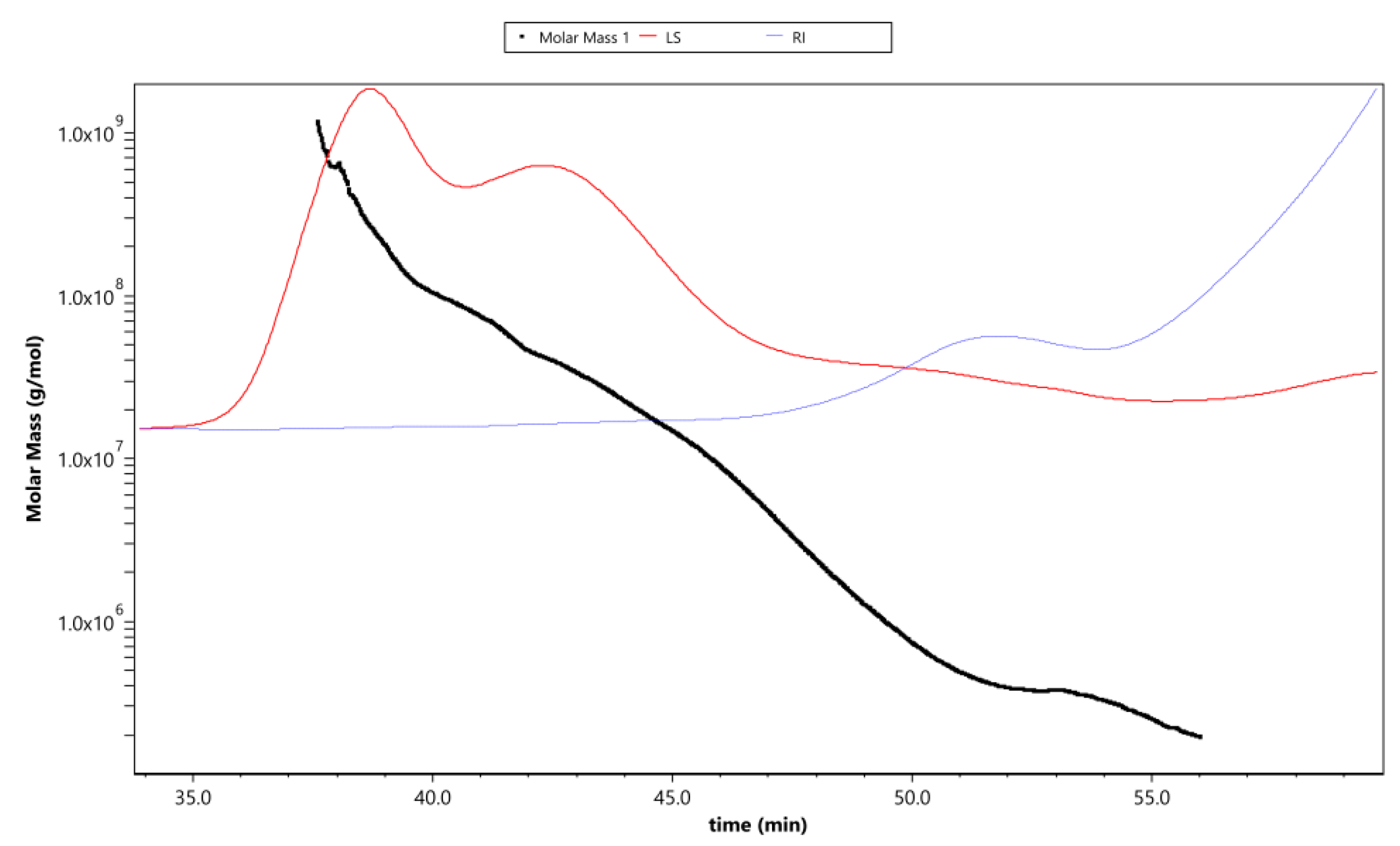
2.3. Determination of Molecular Weight of APS
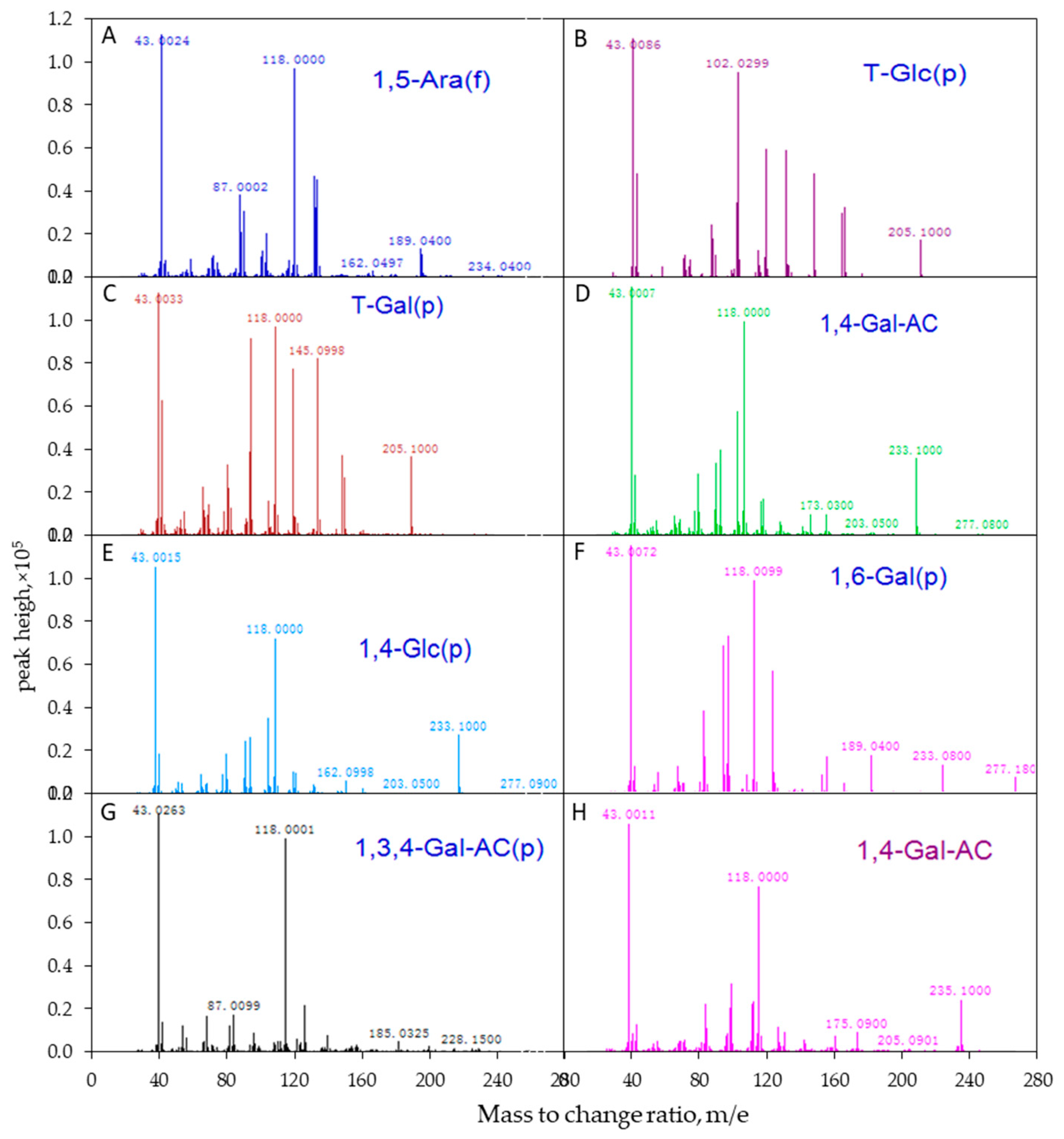
2.4. Determination of Glycosidic Linkages of APS
2.5. The Immunomodulation to Mouse Splenic T and B Lymphocytes
2.5.1. Proliferation Assay of Mouse Splenic T and B Cells
2.5.2. IgM Production of B Cells
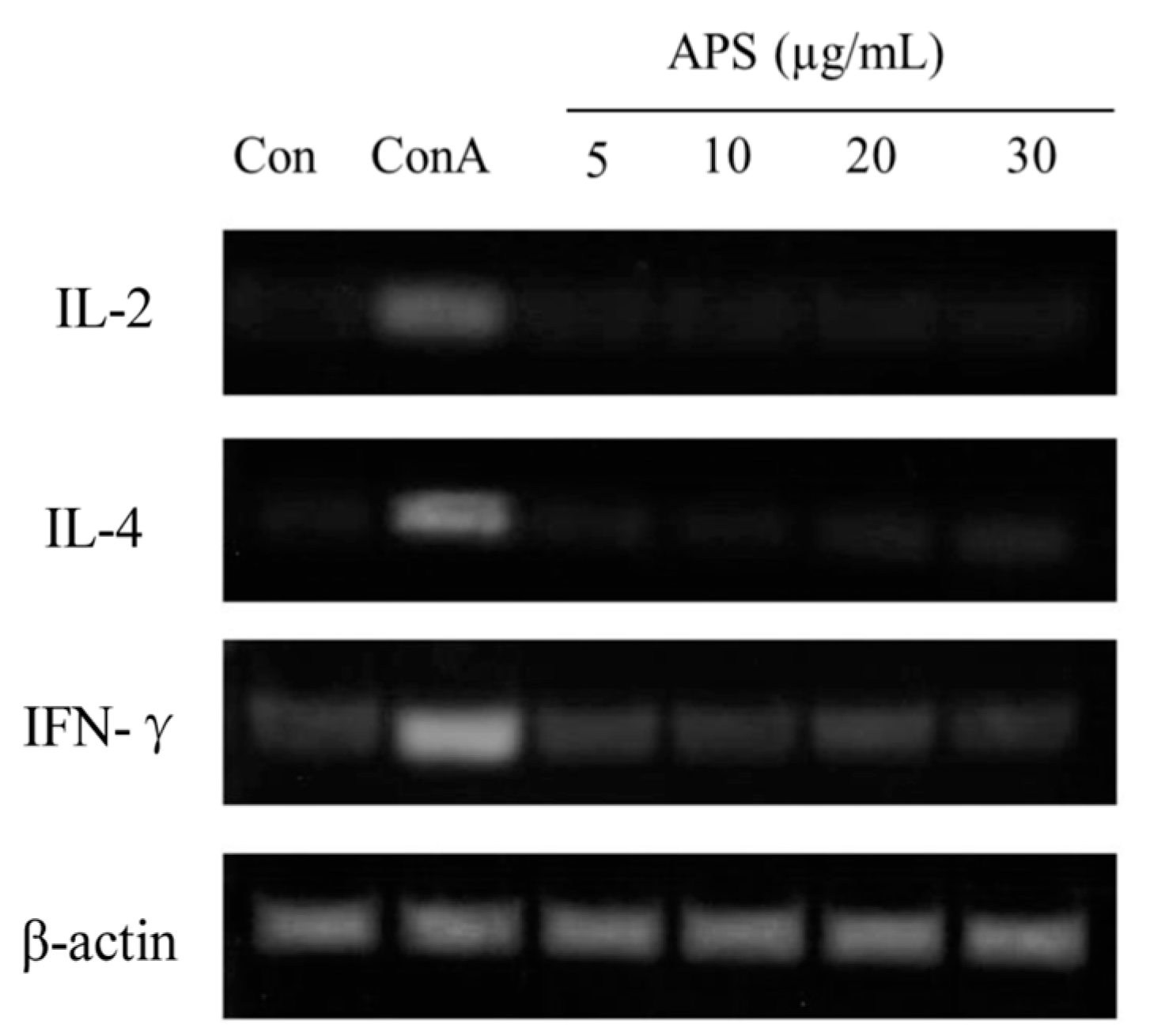
2.5.3. The mRNA Expression of IL-2, IL-4, and IFN-γ in T Cells
2.6. Statistical Analysis
3. Results
3.1. The Monosaccharide Compositions of APS
3.2. Molecular Weight
3.3. Glycosidic Linkages between Monosaccharides
3.4. APS Increased Proliferation and Immune Functions of B but Not T Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| APS | Alfalfa polysaccharides |
| EI-MS | Electron ionization mass spectrometry |
| GalA | Glacturonic acid |
| GlcA | Glucuronic acid |
| GPC-RI-MALS | Gel permeation chromatography–refractive index–multiangle laser light scattering |
| IC | Ion chromatography |
| MEFs | Mouse embryonic fibroblasts |
| Mn | Number-average molecular weight |
| Mw | Weight-average molecular weight |
| Mz | Z-average molecular weight |
| nc | nano coulomb (the unit of quantity of electric charge) |
| Rn | Number-average radius |
| Rw | Weight-average radius |
| Rz | Z-average radius |
| SCFAs | Short-chain fatty acids |
References
- Zhang, J.; Yin, B.; Xie, Y.; Li, J.; Yang, Z.; Zhang, G. Legume-Cereal Intercropping Improves Forage Yield, Quality and Degradability. PLoS ONE 2015, 10, e144813. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.C.; Kinsella, J.E. Functional properties of novel proteins: Alfalfa leaf protein. J. Food Sci. 1976, 41, 286–292. [Google Scholar] [CrossRef]
- Chen, X.; Nie, W.; Fan, S.; Zhang, J.; Wang, Y.; Lu, J.; Jin, L. A polysaccharide from Sargassum fusiforme protects against immunosuppression in cyclophosphamide-treated mice. Carbohyd. Polym. 2012, 90, 1114–1119. [Google Scholar] [CrossRef] [PubMed]
- Ramberg, J.E.; Nelson, E.D.; Sinnott, R.A. Immunomodulatory dietary polysaccharides: A systematic review of the literature. Nutr. J. 2010, 9, 54. [Google Scholar] [CrossRef]
- Chen, L.; Liu, J.; Zhang, Y.; Dai, B.; An, Y.; Yu, L.L. Structural, thermal, and anti-inflammatory properties of a novel pectic polysaccharide from alfalfa (Medicago sativa L.) stem. J. Agric. Food Chem. 2015, 63, 3219–3228. [Google Scholar] [CrossRef]
- Wang, L.; Xie, Y.; Yang, W.; Yang, Z.; Jiang, S.; Zhang, C.; Zhang, G. Alfalfa polysaccharide prevents H2O2-induced oxidative damage in MEFs by activating MAPK/Nrf2 signaling pathways and suppressing NF-κB signaling pathways. Sci. Rep.-UK 2019, 9, 1782. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, Q.; Liu, X.; Ma, L.; Lai, F. Extraction of polysaccharides and its antitumor activity on Magnolia kwangsiensis Figlar & Noot. Carbohyd. Polym. 2016, 142, 98–104. [Google Scholar]
- Zhang, C.Y.; Gan, L.P.; Du, M.Y.; Shang, Q.H.; Xie, Y.H.; Zhang, G.G. Effects of dietary supplementation of alfalfa polysaccharides on growth performance, small intestinal enzyme activities, morphology, and large intestinal selected microbiota of piglets. Livest. Sci. 2019, 223, 47–52. [Google Scholar] [CrossRef]
- Chen, H.L.; Li, D.F.; Chang, B.Y.; Gong, L.M.; Dai, J.G.; Yi, G.F. Effects of Chinese herbal polysaccharides on the immunity and growth performance of young broilers. Poult. Sci. 2003, 82, 364–370. [Google Scholar] [CrossRef]
- Zhang, T.; Yang, Y.; Liang, Y.; Jiao, X.; Zhao, C. Beneficial Effect of Intestinal Fermentation of Natural Polysaccharides. Nutrients 2018, 10, 1055. [Google Scholar] [CrossRef]
- Lahouar, L.; Pochart, P.; Salem, H.B.; El Felah, M.; Mokni, M.; Magne, F.; Mangin, I.; Suau, A.; Pereira, E.; Hammami, M. Effect of dietary fibre of barley variety ‘Rihane’on azoxymethane-induced aberrant crypt foci development and on colonic microbiota diversity in rats. Br. J. Nutr. 2012, 108, 2034–2042. [Google Scholar] [CrossRef] [PubMed]
- Deetz, D.A.; Jung, H.G.; Buxton, D.R. Water-deficit effects on cell-wall composition and in vitro degradability of structural polysaccharides from alfalfa stems. Crop Sci. 1996, 36, 383–388. [Google Scholar] [CrossRef]
- Zou, Y.; Xie, H.B.; Yu, Q.F.; Li, J.C.; He, S.P.; He, J.H. Plant Polysaccharides: Effects on lymphocytes proliferation and cytokines secretion of weaner piglets. Chin. J. Anim. Nutr. 2014, 26, 210–218. [Google Scholar]
- Lamsal, B.P.; Koegel, R.G.; Gunasekaran, S. Some physicochemical and functional properties of alfalfa soluble leaf proteins. LWT-Food Sci. Technol. 2007, 40, 1520–1526. [Google Scholar] [CrossRef]
- Aspinall, G.O.; Gestetner, B.; Molloy, J.A.; Uddin, M. Pectic substances from lucerne (Medicago sativa). Part II. Acidic oligosaccharides from partial hydrolysis of leaf and stem pectic acids. J. Chem. Soc. C Org. 1968, 1, 2554–2559. [Google Scholar] [CrossRef]
- Li, J.; Tang, Y.; Meng, X.; Guan, N.; Xiao, H.; Liu, T.; Yu, Y. The proliferative effects of alfalfa polysaccharides on the mouse immune cells. Life Sci. J. 2013, 10, 868–873. [Google Scholar]
- Samac, D.A.; Litterer, L.; Temple, G.; Jung, H.G.; Somers, D.A. Expression of UDP-glucose dehydrogenase reduces cell-wall polysaccharide concentration and increases xylose content in alfalfa stems. Appl. Biochem. Biotechnol. 2004, 116, 1167–1182. [Google Scholar] [CrossRef]
- Wei, C.; Zhang, Y.; Zhang, H.; Li, J.; Tao, W.; Linhardt, R.J.; Chen, S.; Ye, X. Physicochemical properties and conformations of water-soluble peach gums via different preparation methods. Food Hydrocoll. 2018, 95, 571–579. [Google Scholar] [CrossRef]
- Wang, S.; Dong, X.; Tong, J. Optimization of enzyme-assisted extraction of polysaccharides from alfalfa and its antioxidant activity. Int. J. Biol. Macromol. 2013, 62, 387–396. [Google Scholar] [CrossRef]
- Khatua, S.; Acharya, K. Alkali treated antioxidative crude polysaccharide from Russula alatoreticula potentiates murine macrophages by tunning TLR/NF-κB pathway. Sci. Rep. UK 2019, 9, 1713. [Google Scholar] [CrossRef]
- Li, M.; Yan, Y.; Yu, Q.; Deng, Y.; Wu, D.; Wang, Y.; Ge, Y.; Li, S.; Zhao, J. Comparison of Immunomodulatory Effects of Fresh Garlic and Black Garlic Polysaccharides on RAW 264.7 Macrophages. J. Food Sci. 2017, 82, 765–772. [Google Scholar] [CrossRef]
- Kanwal, S.; Joseph, T.P.; Owusu, L.; Xiaomeng, R.; Meiqi, L.; Yi, X. A Polysaccharide Isolated from Dictyophora indusiata Promotes Recovery from Antibiotic-Driven Intestinal Dysbiosis and Improves Gut Epithelial Barrier Function in a Mouse Model. Nutrients 2018, 10, 1003. [Google Scholar] [CrossRef]
- Park, J.Y.; Shin, M.S.; Kim, S.N.; Kim, H.Y.; Kim, K.H.; Shin, K.S.; Kang, K.S. Polysaccharides from Korean Citrus hallabong peels inhibit angiogenesis and breast cancer cell migration. Int. J. Biol. Macromol. 2016, 85, 522–529. [Google Scholar] [CrossRef]
- Park, H.; Hong, J.; Kwon, H.; Kim, Y.; Lee, K.; Kim, J.; Song, S.K. TLR4-mediated activation of mouse macrophages by Korean mistletoe lectin-C (KML-C). Biochem. Biophys. Res. Commun. 2010, 396, 721–725. [Google Scholar] [CrossRef]
- Sevag, M.G.; Lackman, D.B.; Smolens, J. The isolation of the components of streptococcal nucleoproteins in serologically active form. J. Biol. Chem. 1938, 124, 425–436. [Google Scholar]
- Albrecht, K.A.; Marten, G.C.; Halgerson, J.L.; Wedin, W.F. Analysis of Cell-Wall Carbohydrates and Starch in Alfalfa by near Infrared Reflectance Spectroscopy 1. Crop Sci. 1987, 27, 586–588. [Google Scholar] [CrossRef]
- Roe, J.H. The determination of sugar in blood and spinal fluid with anthrone reagent. J. Biol. Chem. 1955, 212, 335–343. [Google Scholar]
- Oberlerchner, J.T.; Rosenau, T.; Potthast, A. Overview of Methods for the Direct Molar Mass Determination of Cellulose. Molecules (Basel Switzerland) 2015, 20, 10313–10341. [Google Scholar] [CrossRef]
- Huang, Y.; Jhou, B.; Chen, S.; Khoo, K. Identifying Specific and Differentially Linked Glycosyl Residues in Mammalian Glycans by Targeted LC-MS Analysis. Anal. Sci. Int. J. Jpn. Soc. Anal. Chem. 2018, 34, 1049–1054. [Google Scholar] [CrossRef] [Green Version]
- Řezanka, T.; Sigler, K. Structural analysis of a polysaccharide fromChlorella kessleri by means of gas chromatography—Mass spectrometry of its saccharide alditols. Folia Microbiol. 2007, 52, 246. [Google Scholar] [CrossRef]
- Han, S.B.; Yoon, Y.D.; Ahn, H.J.; Lee, H.S.; Lee, C.W.; Yoon, W.K.; Park, S.K.; Kim, H.M. Toll-like receptor-mediated activation of B cells and macrophages by polysaccharide isolated from cell culture of Acanthopanax senticosus. Int. Immunopharmacol. 2003, 3, 1301–1312. [Google Scholar] [CrossRef]
- Feng, W.; Ao, H.; Peng, C. Gut Microbiota, Short-Chain Fatty Acids, and Herbal Medicines. Front. Pharmacol. 2018, 9, 1354. [Google Scholar] [CrossRef] [PubMed]
- Chahal, D.S.; Sivamani, R.K.; Rivkah Isseroff, R.; Dasu, M.R. Plant-Based Modulation of Toll-like Receptors: An Emerging Therapeutic Model. Phytother. Res. 2013, 27, 1423–1438. [Google Scholar] [CrossRef]
- Wei, C.; Zhang, Y.; He, L.; Cheng, J.; Li, J.; Tao, W.; Mao, G.; Zhang, H.; Linhardt, R.J.; Ye, X.; et al. Structural characterization and anti-proliferative activities of partially degraded polysaccharides from peach gum. Carbohyd. Polym. 2019, 203, 193–202. [Google Scholar] [CrossRef]
- Wang, S.; Dong, X.; Ma, H.; Cui, Y.; Tong, J. Purification, characterisation and protective effects of polysaccharides from alfalfa on hepatocytes. Carbohydr. Polym. 2014, 112, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Anonye, B.O. General Commentary on: Alternatives to Antibiotic Growth Promoters in Animals. Front Vet Sci. 2016, 3, 74. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Huang, G. Preparation and immunological activity of polysaccharides and their derivatives. Int. J. Biol. Macromol. 2018, 112, 211–216. [Google Scholar] [CrossRef]
- Fujisaka, S.; Avila-Pacheco, J.; Soto, M.; Kostic, A.; Dreyfuss, J.M.; Pan, H.; Ussar, S.; Altindis, E.; Li, N.; Bry, L.; et al. Diet, Genetics, and the Gut Microbiome Drive Dynamic Changes in Plasma Metabolites. Cell Rep. 2018, 22, 3072–3086. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
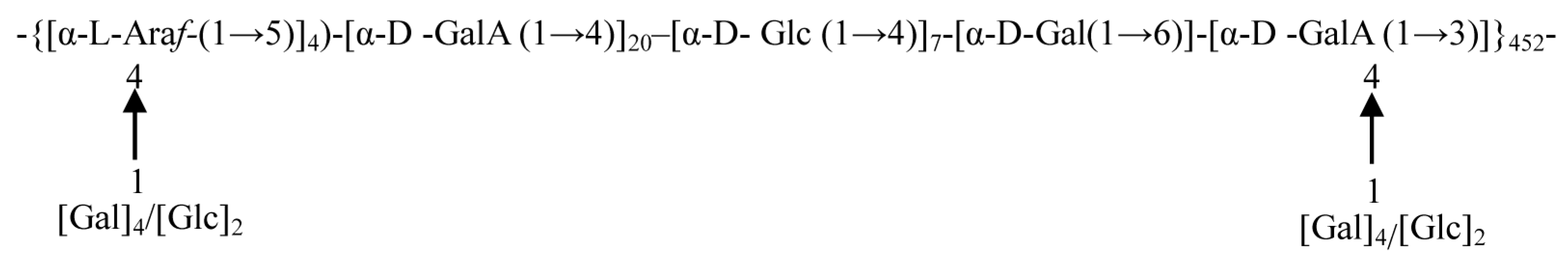{kind=link}
{kind=link}
| Chemical Component | Neutral Sugar (%) | Protein (%) | Yield of Crude Polysaccharide (w/w) % | |||
|---|---|---|---|---|---|---|
| 96.38 ± 4.35 | 3.62 ± 0.48 | 15.76 ± 1.67 | ||||
| Component | RT, min | nc | nc × time | Relative Peak Area | μg/mg | mol/mol, % |
| Fucose | 3.82 ± 0.08 | 9.84 ± 0.08 | 3.27 ± 0.19 | 1.75 ± 0.05 | 4.38 ± 0.06 | 2.02 ± 0.02 |
| Arabinose | 8.29 ± 0.02 | 49.45 ± 0.03 | 13.28 ± 0.04 | 6.79 ± 0.10 | 12.40 ± 0.12 | 6.25 ± 0.06 |
| Galactose | 10.51 ± 0.01 | 52.83 ± 0.21 | 20.27 ± 0.41 | 10.26 ± 0.20 | 8.72 ± 0.07 | 3.66 ± 0.02 |
| Glucose | 12.05 ± 0.08 | 54.68 ± 0.08 | 24.44 ± 0.35 | 12.35 ± 0.12 | 39.20 ± 0.09 | 16.46 ± 0.03 |
| Xylose | 14.22 ± 0.03 | 12.68 ± 0.37 | 4.45 ± 0.33 | 2.48 ± 0.21 | 4.69 ± 0.11 | 2.36 ± 0.05 |
| Mannose | 14.91 ± 0.11 | 7.58 ± 0.14 | 2.80 ± 0.06 | 1.68 ± 0.21 | 6.74 ± 0.16 | 0.77 ± 0.02 |
| GalA | 32.16 ± 0.06 | 223.51 ± 9.31 | 47.11 ± 0.92 | 23.75 ± 0.13 | 146.63 ± 0.18 | 57.13 ± 0.03 |
| GlcA | 34.17 ± 0.04 | 30.83 ± 0.12 | 9.27 ± 0.08 | 4.63 ± 0.20 | 29.16 ± 0.22 | 11.36 ± 0.08 |
| Item | Values | |
|---|---|---|
| Molar mass (g/mol) | Mw | (3.301 ± 0.012) × 106 |
| Mn | (4.057 ± 0.013) × 105 | |
| Mz | (1.433 ± 0.036) × 108 | |
| Polydispersity | Mw/Mn | 8.14 ± 0.02 |
| Mz/Mn | 353.31 ±13.77 | |
| Radius mean square (R.M.S.), nm | Rn | 53.4 ± 3.3 |
| Rw | 51.8 ± 3.3 | |
| Rz | 59.0 ± 3.0 | |
| Serial Number | Retention Time | nc | nc × min | Glycosidic Linkages | mol % |
|---|---|---|---|---|---|
| 1 | 11.64 | 9,757,235 | 2,036,177 | 1,5-Ara(f) | 10.28 |
| 2 | 12.88 | 4,013,392 | 749,606 | T-Glc(p) | 4.02 |
| 3 | 13.55 | 10,293,423 | 2,283,312 | T-Gal(p) | 10.30 |
| 4 | 15.83 | 56,765,889 | 10,600,206 | 1,4-Gal(p)NAC | 52.29 |
| 5 | 16.02 | 18,470,747 | 4,355,016 | 1,4-Glc(p) | 17.02 |
| 6 | 17.51 | 3,820,551 | 722,873 | 1,6-Gal(p) | 3.52 |
| 7 | 17.89 | 3,016,060 | 450,777 | 1,3,4-Gal(p)NAC | 2.57 |
| Incubation Time, h | CON | LPS/ConA | APS concentration, µg/ mL | |||
|---|---|---|---|---|---|---|
| 5 | 10 | 20 | 30 | |||
| B cells | ||||||
| 12 | 0.018b ± 0.002 | 0.033 ** ± 0.003 | 0.021 ± 0.001 | 0.027 * ± 0.002 | 0.024 ± 0.002 | 0.024 ± 0.001 |
| 24 | 0.026 ± 0.003 | 0.0558 ** ± 0.003 | 0.031 ± 0.002 | 0.037 * ± 0.003 | 0.029 ± 0.003 | 0.033 ± 0.003 |
| 48 | 0.012 ± 0.001 | 0.024 * ± 0.004 | 0.017 ± 0.002 | 0.023 * ± 0.002 | 0.019 ± 0.002 | 0.018 ± 0.002 |
| T cells | ||||||
| 12 | 0.048 ± 0.0105 | 0.152 ** ± 0.015 | 0.061 ± 0.014 | 0.059 ± 0.008 | 0.056 ± 0.012 | 0.058 ± 0.009 |
| 24 | 0.081 ± 0.011 | 0.163 ** ± 0.019 | 0.085 ± 0.006 | 0.089 ± 0.004 | 0.09 ± 0.005 | 0.089 ± 0.006 |
| 48 | 0.028 ± 0.007 | 0.139 ** ± 0.024 | 0.031 ± 0.007 | 0.034 ± 0.007 | 0.032 ± 0.006 | 0.04 ± 0.008 |
| IgM (cytokine of B cells) | ||||||
| 24 | 0.391 ± 0.014 | 0.4107 ± 0.012 | 0.391 ± 0.011 | 0.412 ± 0.007 | 0.401 ± 0.006 | 0.401 ± 0.010 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Li, Z.; Zhang, C.-Y.; Li, M.; Lee, Y.; Zhang, G.-G. Extract Methods, Molecular Characteristics, and Bioactivities of Polysaccharide from Alfalfa (Medicago sativa L.). Nutrients 2019, 11, 1181. https://doi.org/10.3390/nu11051181
Zhang C, Li Z, Zhang C-Y, Li M, Lee Y, Zhang G-G. Extract Methods, Molecular Characteristics, and Bioactivities of Polysaccharide from Alfalfa (Medicago sativa L.). Nutrients. 2019; 11(5):1181. https://doi.org/10.3390/nu11051181
Chicago/Turabian StyleZhang, Chen, Zemin Li, Chong-Yu Zhang, Mengmeng Li, Yunkyoung Lee, and Gui-Guo Zhang. 2019. "Extract Methods, Molecular Characteristics, and Bioactivities of Polysaccharide from Alfalfa (Medicago sativa L.)" Nutrients 11, no. 5: 1181. https://doi.org/10.3390/nu11051181
APA StyleZhang, C., Li, Z., Zhang, C. -Y., Li, M., Lee, Y., & Zhang, G. -G. (2019). Extract Methods, Molecular Characteristics, and Bioactivities of Polysaccharide from Alfalfa (Medicago sativa L.). Nutrients, 11(5), 1181. https://doi.org/10.3390/nu11051181