Allium porrum Extract Decreases Effector Cell Degranulation and Modulates Airway Epithelial Cell Function

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material
2.3. Extract Preparation by Ultrasonication
2.4. Determination of Total Phenols Content
2.5. Antioxidant Activity
2.5.1. Determination of Total Antioxidant Activity
2.5.2. Determination of DPPH Free Radical Scavenging Activity
2.6. Gas Chromatography–Mass Spectrometry (GC–MS) Analysis
2.7. Liquid Chromatography–Mass Spectrometry (LC–IT/MS)
2.8. Cell Culture
2.9. Cytotoxicity Assay
2.10. Interleukin (IL) 6 Analysis
2.11. Epithelial Permeability Analysis
2.12. Degranulation Assay
2.13. RNA Extraction and Quantitative RT-PCR (qRT-PCR) Analysis
2.14. Statistical Analysis
3. Results
3.1. Characterization of Leek Extract
3.2. Antioxidant Activity of Leek Extract
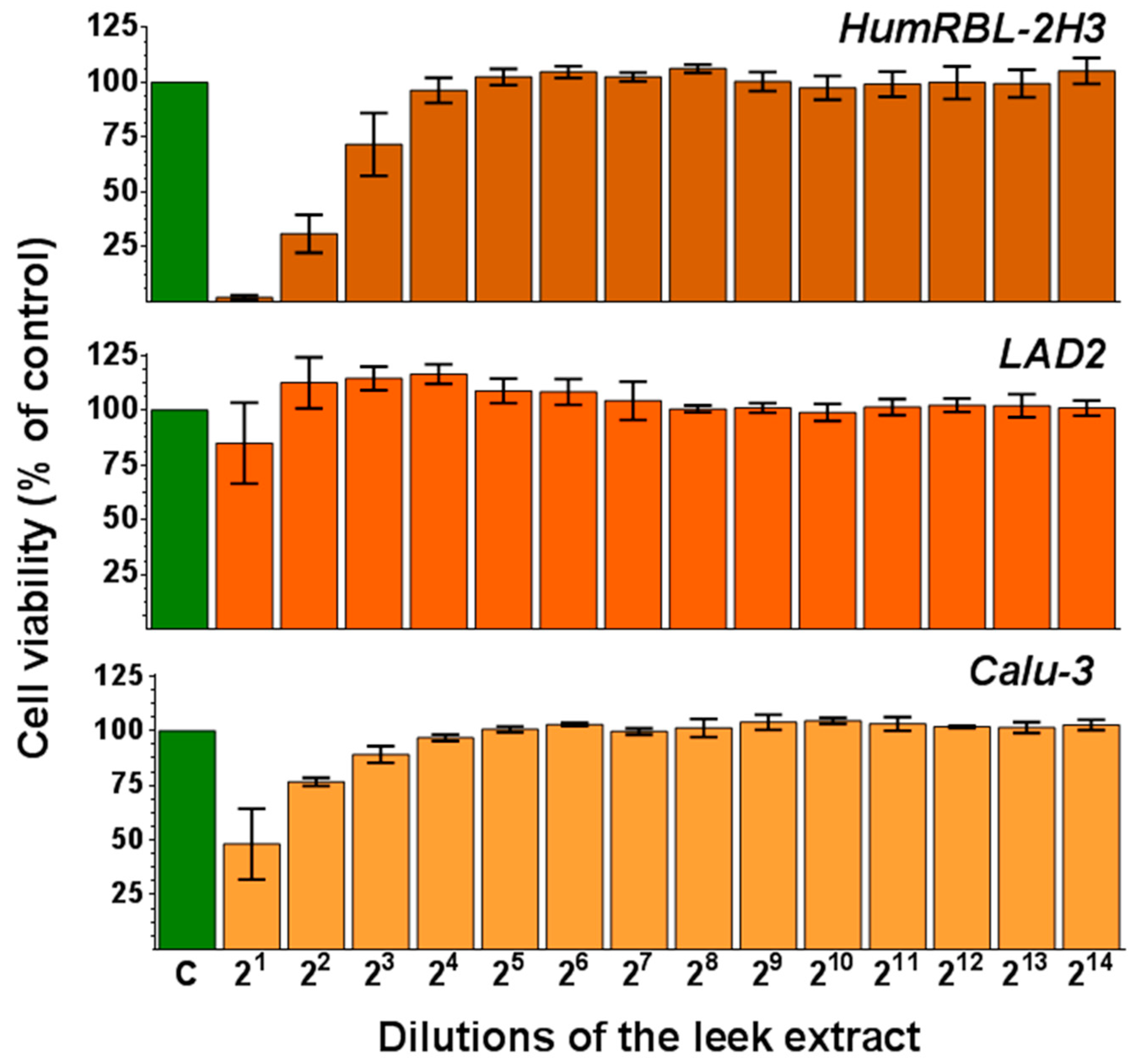
3.3. Leek Extract Exhibits Cytotoxic Activity in a Dose-Dependent Manner
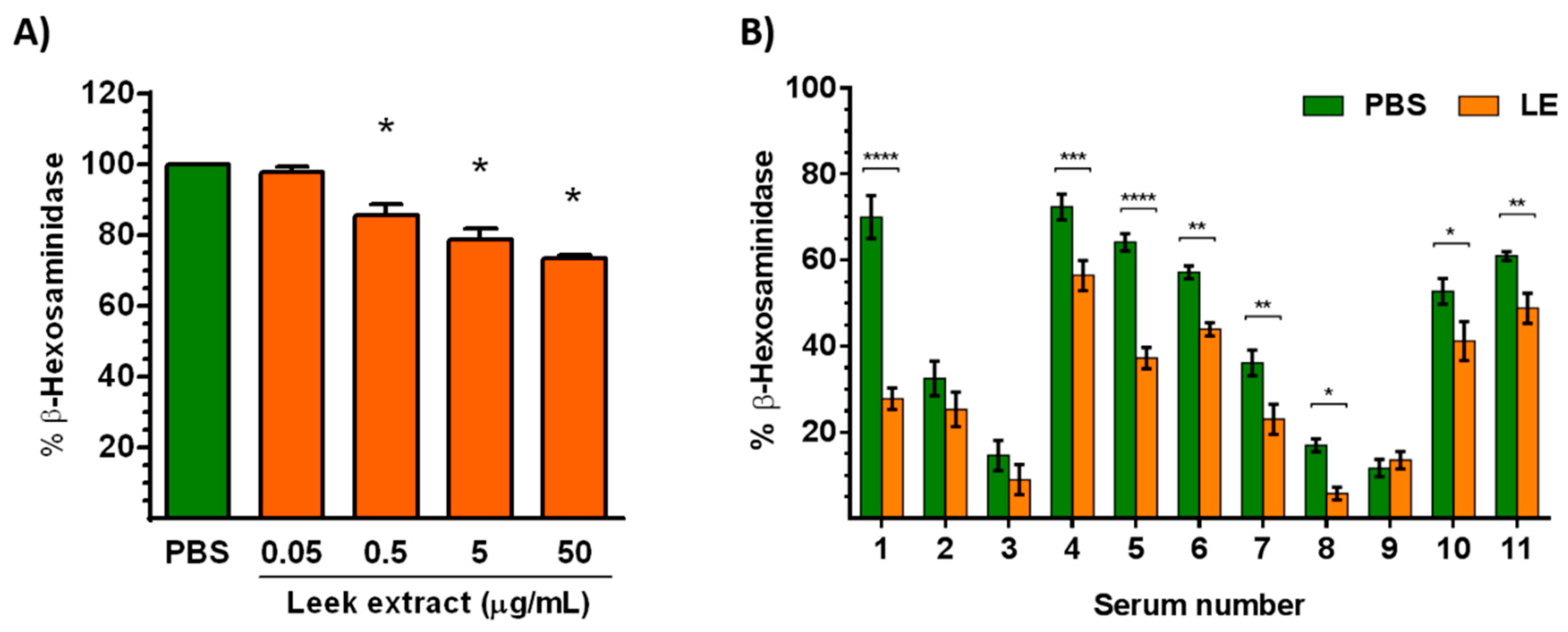
3.4. Leek Extract Decreases Degranulation of Mast Cells
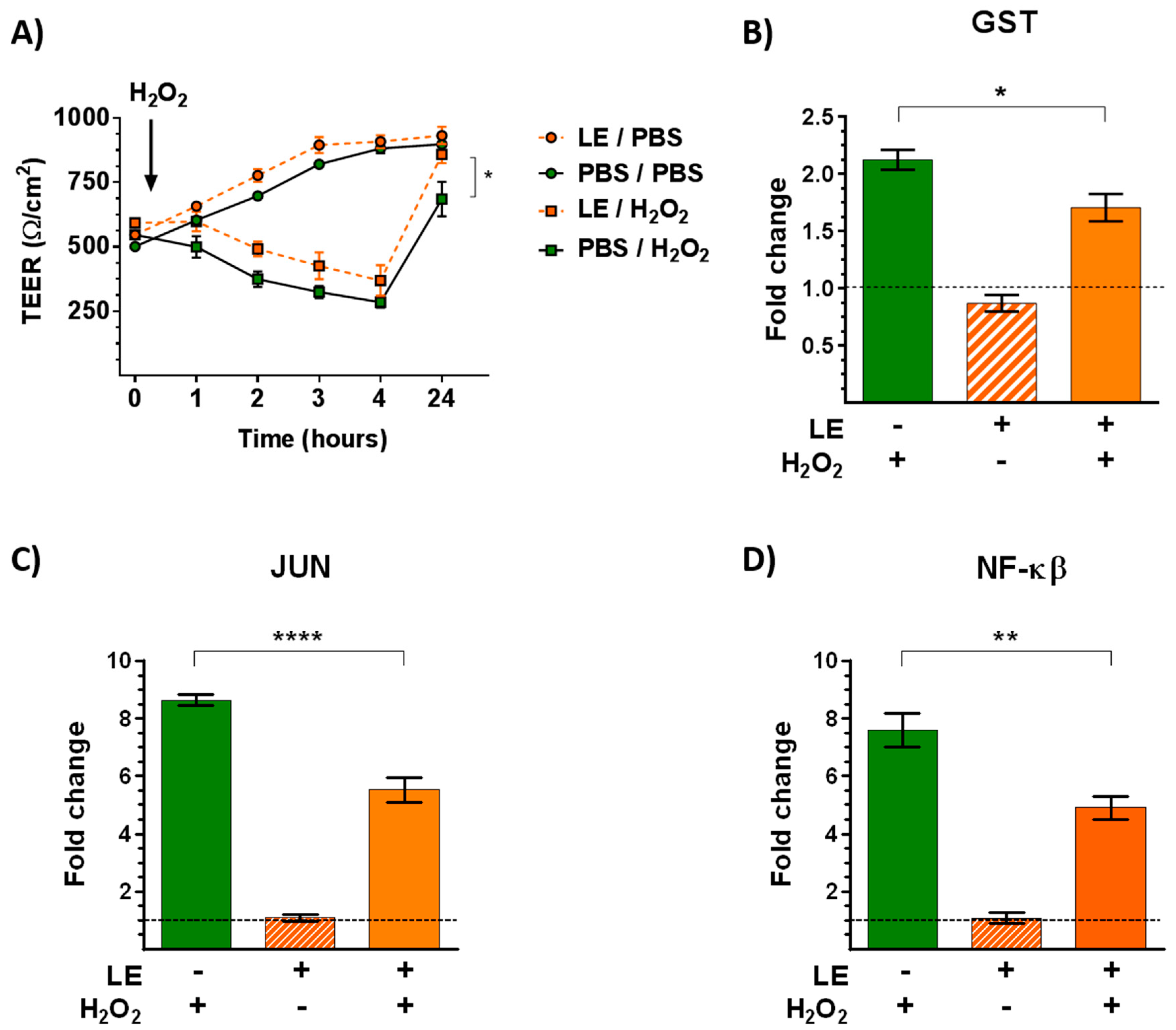
3.5. Leek Extract Prevents Both the Decrease of TEER and Gene Expression Induced by H2O2 Oxidative Stress/Inflammatory Stimulus
3.6. Leek Extract Decreases Epithelial Permeability to an Allergen
3.7. Leek Extract Decreases the Apical Release of IL-6 by Bronchial Epithelial Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. WHO Monographs On Selected Medicinal Plants; World Health Organization: Geneva, Switzerland, 1999. [Google Scholar]
- Pawankar, R.; Canonica, G.W.; Holgate, S.T.; Lockey, R.F.; Blaiss, M.S. The WAO White Book on Allergy: Update 2013; World Allergy Organization: Milwaukee, WI, USA, 2013. [Google Scholar]
- Durrani, S.R.; Viswanathan, R.K.; Busse, W.W. What effect does asthma treatment have on airway remodeling? Current perspectives. J. Allergy Clin. Immunol. 2011, 128, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Finn, D.F.; Walsh, J.J. Twenty-first century mast cell stabilizers. Br. J. Pharmacol. 2013, 170, 23–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, M.; Scott, N.W.; Mustafa, M.Z.; Mustafa, M.S.; Azuara-Blanco, A. Topical antihistamines and mast cell stabilisers for treating seasonal and perennial allergic conjunctivitis. Cochrane Libr. 2015, 6, CD009566. [Google Scholar] [CrossRef] [PubMed]
- Han, S.J.; Bae, E.A.; Trinh, H.T.; Yang, J.H.; Youn, U.J.; Bae, K.H.; Kim, D.H. Magnolol and honokiol: Inhibitors against mouse passive cutaneous anaphylaxis reaction and scratching behaviors. Biol. Pharm. Bull. 2007, 30, 2201–2203. [Google Scholar] [CrossRef] [PubMed]
- Kang, O.H.; Jang, H.J.; Chae, H.S.; Oh, Y.C.; Choi, J.G.; Lee, Y.S.; Kim, J.H.; Kim, Y.C.; Sohn, D.H.; Park, H.; et al. Anti-inflammatory mechanisms of resveratrol in activated HMC-1 cells: Pivotal roles of NFκB and MAPK. Pharmacol. Res. 2009, 59, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, J.W.; Ko, N.Y.; Mun, S.H.; Her, E.; Kim, B.K.; Han, J.W.; Lee, H.Y.; Beaven, M.A.; Kim, Y.M.; et al. Curcumin, a constituent of curry, suppresses IgE-mediated allergic response and mast cell activation at the level of Syk. J. Allergy Clin. Immunol. 2008, 121, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Kimata, M.; Inagaki, N.; Nagai, H. Effects of luteolin and other flavonoids on IgE-mediated allergic reactions. Planta Med. 2000, 66, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Jeong, H.J.; Kim, H.M. Theanine is a candidate amino acid for pharmacological stabilization of mast cells. Amino Acids 2012, 42, 1609–1618. [Google Scholar] [CrossRef]
- Penissi, A.B.; Vera, M.E.; Mariani, M.L.; Rudolph, M.I.; Ceñal, J.P.; de Rosas, J.C.; Fogal, T.H.; Tonn, C.E.; Favier, L.S.; Giordano, O.S.; et al. Novel anti-ulcer α, β-unsaturated lactones inhibit compound 48/80-induced mast cell degranulation. Eur. J. Pharmacol. 2009, 612, 122–130. [Google Scholar] [CrossRef]
- Huang, F.; Yamaki, K.; Tong, X.; Fu, L.; Zhang, R.; Cai, Y.; Yanagisawa, R.; Inoue, K.; Takano, H.; Yoshino, S. Inhibition of the antigen-induced activation of RBL-2H3 cells by sinomenine. Int. Immunopharmacol. 2008, 8, 502–507. [Google Scholar] [CrossRef]
- Qu, J.; Li, Y.; Zhong, W.; Gao, P.; Hu, C. Recent developments in the role of reactive oxygen species in allergic asthma. J. Thorac. Dis. 2017, 9, E32. [Google Scholar] [CrossRef] [PubMed]
- Corzo-Martinez, M.; Corzo, N.; Villamiel, M. Biological properties of onions and garlic. Trends Food Sci. Technol. 2007, 18, 609–625. [Google Scholar] [CrossRef]
- Rizwani, G.H.; Shareef, H. Genus Allium: The potential nutritive and therapeutic source. JPANS 2011, 1, 158–165. [Google Scholar] [CrossRef]
- Lee, K.H.; Park, E.; Lee, H.J.; Kim, M.O.; Cha, Y.J.; Kim, J.M.; Lee, H.; Shin, M.J. Effects of daily quercetin-rich supplementation on cardiometabolic risks in male smokers. Nutr. Res. Pract. 2011, 5, 28–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, C.B.T.; Jadeja, G.C. Deep eutectic solvent-based extraction of polyphenolic antioxidants from onion (Allium cepa L.) peel. J. Sci. Food Agric. 2019, 99, 1969–1979. [Google Scholar] [CrossRef] [PubMed]
- Jalalvand, A.R.; Zhaleh, M.; Goorani, S.; Zangeneh, M.M.; Seydi, N.; Zangeneh, A.; Moradi, R. Chemical characterization and antioxidant, cytotoxic, antibacterial, and antifungal properties of ethanolic extract of Allium Saralicum, R.M. Fritsch leaves rich in linolenic acid, methyl ester. J. Photochem. Photobiol. B 2019, 192, 103–112. [Google Scholar] [CrossRef]
- Zare, A.; Farzaneh, P.; Pourpak, Z.; Zahedi, F.; Moin, M.; Shahabi, S.; Hassan, Z.M. Purified aged garlic extract modulates allergic airway inflammation in BALB/c mice. Iran J. Allergy Asthma Immunol. 2008, 7, 133–141. [Google Scholar] [PubMed]
- Di Donna, L.; Mazzotti, F.; Taverna, D.; Napoli, A.; Sindona, G. Structural characterisation of malonyl flavonols in leek (Allium porrum L.) using high-performance liquid chromatography and mass spectrometry. Phytochem. Anal. 2014, 25, 207–212. [Google Scholar] [CrossRef]
- Soininen, T.H.; Jukarainen, N.; Soininen, P.; Auriola, S.O.K.; Julkunen-Tiitto, R.; Oleszek, W.; Stochmal, A.; Karjalainen, R.O.; Vepsalainen, J.J. Metabolite profiling of leek (Allium porrum L) cultivars by H-1 NMR and HPLC-MS. Phytochem. Anal. 2014, 25, 220–228. [Google Scholar] [CrossRef]
- Radovanović, B.; Mladenović, J.; Radovanović, A.; Pavlović, R.; Nikolić, V. Phenolic composition, antioxidant, antimicrobial and cytotoxic activities of Allium porrum L. (Serbia) extracts. J. Food Nutr. Res. 2015, 3, 564–569. [Google Scholar]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C.L.W.T. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Acero, N.; Gradillas, A.; Beltran, M.; García, A.; Muñoz Mingarro, D. Comparison of phenolic compounds profile and antioxidant properties of different sweet cherry (Prunus avium L.) varieties. Food Chem. 2019, 279, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Villalba, M.; Batanero, E.; López-Otín, C.; Sánchez, L.M.; Monsalve, R.I.; González de la Peña, M.A.; Lahoz, C.; Rodríguez, R. Amino acid sequence of Ole e 1, the major allergen from olive tree polen (Olea europea). Eur. J. Biochem. 1993, 103, 147–153. [Google Scholar]
- Kirshenbaum, A.S.; Akin, C.; Wu, Y.; Rottem, M.; Goff, J.P.; Beaven, M.A.; Rao, V.K.; Metcalfe, D.D. Characterization of novel stem cell factor responsive human mast cell lines LAD 1 and 2 established from a patient with mast cell sarcoma/leukemia; activation following aggregation of FcepsilonRI or FcgammaRI. Leuk. Res. 2003, 27, 677–682. [Google Scholar] [CrossRef]
- Gadermaier, G.; Jahn-Schmid, B.; Vogel, L.; Egger, M.; Himly, M.; Briza, P.; Ebner, C.; Vieths, S.; Bohle, B.; Ferreira, F. Targeting the cysteine-stabilized fold of Art v 1 for immunotherapy of Artemisia pollen allergy. Mol. Immunol. 2010, 47, 1292–1298. [Google Scholar] [CrossRef] [PubMed]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef]
- Prado, N.; De Linares, C.; Sanz, M.L.; Gamboa, P.; Villalba, M.; Rodríguez, R.; Batanero, E. Pollensomes as natural vehicles for pollen allergens. J. Immunol. 2015, 195, 445–449. [Google Scholar] [CrossRef]
- Vogel, L.; Lüttkopf, D.; Hatahet, L.; Haustein, D.; Vieths, S. Development of a functional in vitro assay as a novel tool for the standardization of allergen extracts in the human system. Allergy 2005, 60, 1021–1028. [Google Scholar] [CrossRef]
- Kachlicki, P.; Piasecka, A.; Stobiecki, M.; Marczak, L. Structural Characterization of Flavonoid Glycoconjugates and Their Derivatives with Mass Spectrometric Techniques. Molecules 2016, 21, 1494. [Google Scholar] [CrossRef]
- García-Herrera, P.; Morales, P.; Fernández-Ruiz, V.; Sánchez-Mata, M.C.; Cámara, M.; Carvalho, A.M.; Ferreira, I.C.F.R.; Pardo-de-Santayana, M.; Molina, M.; Tardio, J. Nutrients, phytochemicals and antioxidant activity in wild populations of Allium ampeloprasum L., a valuable underutilized vegetable. Food Res. Int. 2014, 62, 272–279. [Google Scholar] [CrossRef]
- Bernaert, N.; De Paepe, D.; Bouten, C.; De Clercq, H.; Stewart, D.; Van Bockstaele, E.; De Loose, M.; Van Droogenbroeck, B. Antioxidant capacity, total phenolic and ascorbate content as a function of the genetic diversity of leek (Allium ampeloprasum var. porrum). Food Chem. 2012, 134, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Shen, X.; Cheng, S.; Li, P.; Du, J.; Chang, Y.; Meng, H. Evaluation of garlic cultivars for polyphenolic content and antioxidant properties. PLoS ONE 2013, 8, e79730. [Google Scholar] [CrossRef] [PubMed]
- Rogerio, A.P.; Kanashiro, A.; Fontanari, C.; Da Silva, E.V.G.; Lucisano-Valim, Y.M.; Soares, E.G.; Faccioli, L.H. Anti-inflammatory activity of quercetin and isoquercitrin in experimental murine allergic asthma. Inflamm. Res. 2007, 56, 402–408. [Google Scholar] [CrossRef]
- Kaiser, P.; Youssouf, M.S.; Tasduq, S.A.; Singh, S.; Sharma, S.C.; Singh, G.D.; Gupta, V.K.; Gupta, B.D.; Johri, R.K. Anti-allergic effects of herbal product from Allium cepa (bulb). J. Med. Food. 2009, 12, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Scherer, R.; Godoy, H.T. Effects of extraction methods of phenolic compounds from Xanthium strumarium L. and their antioxidant activity. Rev. Bras. Plantas Med. 2014, 16, 41–46. [Google Scholar] [CrossRef]
- Yamada, K.E.; Eckhert, C.D. Boric acid activation of eIF2α and Nrf2 Is PERK dependent: A mechanism that explains how boron prevents DNA damage and enhances antioxidant status. Biol. Trace Elem. Res. 2019, 188, 2–10. [Google Scholar] [CrossRef]
- Linker, R.A.; Lee, D.H.; Ryan, S.; van Dam, A.M.; Conrad, R.; Bista, P.; Zeng, W.; Hronowsky, X.; Buko, A.; Chollate, S.; et al. Fumaric acid esters exert neuroprotective effects in neuroinflammation via activation of the Nrf2 antioxidant pathway. Brain 2011, 134, 678–692. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.J.; Liang, C.L.; Li, G.M.; Yu, C.Y.; Yin, M. Stearic acid protects primary cultured cortical neurons against oxidative stress. Acta Pharmacol. Sin. 2007, 28, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.C.; Yen, P.L.; Chang, S.T.; Cheng, P.L.; Lo, Y.C.; Liao, V.H. Antioxidative activities of both oleic acid and Camellia tenuifolia seed oil are regulated by the transcription factor DAF-16/FOXO in Caenorhabditis elegans. PLoS ONE 2016, 11, e0157195. [Google Scholar] [CrossRef]
- Luo, J.B.; Feng, L.; Jiang, W.D.; Liu, Y.; Wu, P.; Jiang, J.; Kuang, S.Y.; Tang, L.; Zhang, Y.A.; Zhou, X.Q. The impaired intestinal mucosal immune system by valine deficiency for young grass carp (Ctenopharyngodon idella) is associated with decreasing immune status and regulating tight junction proteins transcript abundance in the intestine. Fish Shellfish Immunol. 2014, 40, 197–207. [Google Scholar] [CrossRef]
- Ren, W.; Zou, L.; Ruan, Z.; Li, N.; Wang, Y.; Peng, Y.; Liu, G.; Yin, Y.; Li, T.; Hou, Y.; et al. Dietary L-proline supplementation confers immunostimulatory effects on inactivated Pasteurella Multocida vaccine immunized mice. Amino Acids 2013, 45, 555–561. [Google Scholar] [CrossRef]
- Seabra, R.M.; Andrade, P.B.; Valentão, P.; Fernandes, E.; Carvalho, F.; Bastos, M.L. Anti-oxidant compounds extracted from several plant materials. In Biomaterials from Aquatic and Terrestrial Organisms, 1st ed.; Fingerman, M., Ed.; CRC Press: Boca Raton, FL, USA, 2006; Chapter 4. [Google Scholar] [CrossRef]
- Piluzza, G.; Bullitta, S. Correlations between phenolic content and antioxidant properties in twenty-four plant species of traditional ethnoveterinary use in the Mediterranean area. Pharm. Biol. 2011, 49, 240–247. [Google Scholar] [CrossRef]
- Grzesik, M.; Naparło, K.; Bartosz, G.; Sadowska-Bartosz, I. Antioxidant properties of catechins: Comparison with other antioxidants. Food Chem. 2018, 241, 480–492. [Google Scholar] [CrossRef]
- Qu, W.; Fan, L.; Kim, Y.C.; Ishikawa, S.; Iguchi-Ariga, S.M.; Pu, X.P.; Ariga, H. Kaempferol derivatives prevent oxidative stress-induced cell death in a DJ-1-dependent manner. J. Pharmacol. Sci. 2009, 110, 191–200. [Google Scholar] [CrossRef]
- Nile, S.H.; Nile, A.S.; Keum, Y.S.; Sharma, K. Utilization of quercetin and quercetin glycosides from onion (Allium cepa L.) solid waste as an antioxidant, urease and xanthine oxidase inhibitors. Food Chem. 2017, 235, 119–126. [Google Scholar] [CrossRef]
- Gunathilake, K.D.P.P.; Ranaweera, K.K.D.S. Antioxidative properties of 34 green leafy vegetables. J. Funct. Foods 2016, 26, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Assadpour, S.; Nabavi, S.M.; Nabavi, S.F.; Dehpour, A.A.; Ebrahimzadeh, M.A. In vitro antioxidant and antihemolytic effects of the essential oil and methanolic extract of Allium rotundum L. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 5210–5215. [Google Scholar]
- Yoo, J.M.; Sok, D.E.; Kim, M.R. Anti-allergic action of aged black garlic extract in RBL-2H3 cells and passive cutaneous anaphylaxis reaction in mice. J. Med. Food. 2014, 17, 92–102. [Google Scholar] [CrossRef]
- Lee, E.; Kim, S.G.; Park, N.Y.; Park, H.H.; Jeong, K.T.; Choi, J.; Lee, I.H.; Lee, H.; Kim, K.J.; Lee, E. KOTMIN13, a Korean herbal medicine alleviates allergic inflammation in vivo and in vitro. BMC Complement. Altern. Med. 2016, 16, 169. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, K.; Loridas, S. Pulmonary oxidative stress, inflammation and cancer: Respirable particulate matter, fibrous dusts and ozone as major causes of lung carcinogenesis through reactive oxygen species mechanisms. Int. J. Environ. Res. Public Health 2013, 10, 3886–3907. [Google Scholar] [CrossRef]
- Evans, M.D.; Dizdaroglu, M.; Cooke, M.S. Oxidative DNA damage and disease: Induction, repair and significance. Mutat. Res. 2004, 567, 1–61. [Google Scholar] [CrossRef]
- Jang, H.Y.; Kim, S.M.; Yuk, J.E.; Kwon, O.K.; Oh, S.R.; Lee, H.K.; Jeong, H.; Ahn, K.S. Capsicum annuum L. methanolic extract inhibits ovalbumin-induced airway inflammation and oxidative stress in a mouse model of asthma. J. Med. Food 2011, 14, 1144–1151. [Google Scholar] [CrossRef]
- Lee, M.Y.; Lee, N.H.; Seo, C.S.; Lee, J.A.; Jung, D.; Kim, J.H.; Shin, H.K. Alpinia katsumadai seed extract attenuate oxidative stress and asthmatic activity in a mouse model of allergic asthma. Food Chem. Toxicol. 2010, 48, 1746–1752. [Google Scholar] [CrossRef]
- Catanzaro, D.; Rancan, S.; Orso, G.; Dall’Acqua, S.; Brun, P.; Giron, M.C.; Carrara, M.; Castagliuolo, I.; Ragazzi, E.; Caparrotta, L.; et al. Boswellia serrata preserves intestinal epithelial barrier from oxidative and inflammatory damage. PLoS ONE 2015, 10, e0125375. [Google Scholar] [CrossRef]
- Bhalla, D.K. Ozone-induced lung inflammation and mucosal barrier disruption: Toxicology, mechanisms, and implications. J. Toxicol. Environ. Health B Crit. Rev. 1999, 2, 31–86. [Google Scholar] [CrossRef]
- Janssen, Y.M.; Matalon, S.; Mossman, B.T. Differential induction of c-fos, c-jun, and apoptosis in lung epithelial cells exposed to ROS or RNS. Am. J. Physiol. 1997, 273, 789–796. [Google Scholar] [CrossRef]
- Oakley, A. Glutathione transferases: A structural perspective. Drug Metab. Rev. 2011, 43, 13–151. [Google Scholar] [CrossRef]
- Marini, M.; Vittori, E.; Hollemborg, J.; Mattoli, S. Expression of the potent inflammatory cytokines, granulocyte-macrophage-colony-stimulating factor and interleukin-6 and interleukin-8, in bronchial epithelial cells of patients with asthma. J. Allergy Clin. Immunol. 1992, 89, 1001–1009. [Google Scholar] [CrossRef]
- Oliveira, T.T.; Campos, K.M.; Cerqueira-Lima, A.T.; Carneiro, T.C.B.; da Silva Velozo, E.; Ribeiro Melo, I.C.; Figueiredo, E.A.; de Jesus Oliveira, E.; de Vasconcelos, D.F.; Pontes-de-Carvalho, L.C.; et al. Potential therapeutic effect of Allium cepa L. and quercetin in a murine model of Blomia tropicalis induced asthma. DARU J. Pharm. Sci. 2015, 23, 18. [Google Scholar] [CrossRef]
- Kim, H.Y.; Nam, S.Y.; Hong, S.W.; Kim, M.J.; Jeong, H.J.; Kim, H.M. Protective effects of rutin through regulation of vascular endothelial growth factor in allergic rhinitis. Am. J. Rhinol. Allergy 2015, 29, e87–e94. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Sequence 5′-3′ | Reverse Primer 5′-3′ |
|---|---|---|
| GAPDH | AAAGGGTCATCATCTCTG | GCTGTTGTCATACTTCTC |
| GST | CGGGCAACTGAAGCCTTTTG | TCAGCGAAGGAGATCTGGTC |
| c-Jun | GCAAAGAACTTTCCCGGCTG | GGAGAAGCCTAAGACGCAGG |
| NFκB | TGAGGATGATGAGAATGGAT | CGGAACACAATGGCATAC |
| Compound | Retention Time (min) | Molecular Formula | Molecular Mass (g/mol) | R-Match |
|---|---|---|---|---|
| Organic acids | ||||
| Boric acid | 4.8 | H3BO3 | 61.8 | 814 |
| Propanoic acid | 5.8 | C3H6O2 | 74.1 | 899 |
| Acetic acid | 6.1 | C2H4O2 | 60.1 | 849 |
| Phosphoric acid | 8.9 | H3PO4 | 98.0 | 869 |
| Succinic acid | 9.5 | C4H6O4 | 118.09 | 918 |
| Fumaric acid | 10.0 | C4H4O4 | 116.1 | 776 |
| Nonanoic acid | 10.2 | C9H18O2 | 158.2 | 787 |
| Malic acid | 11.8 | C4H6O5 | 134.1 | 859 |
| Arabinonic acid | 13.5 | C5H10O6 | 166.1 | 729 |
| Palmitic acid | 17.8 | C16H32O2 | 256.4 | 705 |
| Stearic acid | 19.1 | C18H36O2 | 284.5 | 702 |
| Oleic acid | 19.7 | C18H34O2 | 282.5 | 808 |
| Carbohydrates | ||||
| Arabinofuranose | 8.6 | C5H10O | 150.1 | 777 |
| Glucofuranoside | 15.4 | C6H12O6 | 180.2 | 772 |
| D-fructose | 15.5 | C6H12O6 | 180.2 | 809 |
| Mannofuranoside | 15.6 | C7H14O6 | 194.2 | 781 |
| Glucose | 17.1 | C6H12O6 | 180.2 | 790 |
| Galactopyranose | 17.1 | C6H12O6 | 180.2 | 825 |
| Amino acids | ||||
| L-valine | 6.3 | C5H11NO2 | 117.2 | 798 |
| L-proline | 12.2 | C5H9NO2 | 115.1 | 850 |
| Mean ± SD | |
|---|---|
| Total phenol content | 2.3 ± 0.2 a |
| Total antioxidant activity | 60.2 ± 3.1 b |
| DPPH scavenging activity, IC50 | 289.6 ± 6.1 c |
| Polyphenol Subclass (Flavonoids) | Compound Name | Retention Time (min) | Positive Ions (m/z) | ||||
|---|---|---|---|---|---|---|---|
| MW | [M+H]+ | [M+Na]+ | [M+K]+ | In-Source Fragments | |||
| flavanols | (-)-Epicatechin a | 10.0 | 290 | 291 | -- | -- | -- |
| flavanols | (+)-Catechin a | 12.3 | 290 | 291 | -- | -- | -- |
| flavonols | Kaempferol derivative b | 19.4 | 742 | 743 | 765 | -- | -- [Aglycone+H]+ = 287, [Aglycone+H-H2O]+ = 269 |
| flavonols | Kaempferol derivative c | 19.8 | 902 | 903 | 925 | -- | [(M-Rham)+H]+ = 757 [(M-Rham-Glc)+H]+ = 595 [(M-Rham-2Glc)+H]+ = 433 [Aglycone+H]+ = 287, [Aglycone+H-H2O]+ = 269 |
| flavonols | Quercetin derivative b | 18.8 | 625 | 627 | -- | -- | [(M-Glc)+H]+ = 463 [Aglycone+H]+ = 303, [Aglycone+H-H2O]+ = 285 |
| flavonols | Quercetin derivative b | 20.5 | 609 | 611 | 633 | 649 | [Aglycone+H]+ = 303, [Aglycone+H-H2O]+ = 285 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benedé, S.; Gradillas, A.; Villalba, M.; Batanero, E. Allium porrum Extract Decreases Effector Cell Degranulation and Modulates Airway Epithelial Cell Function. Nutrients 2019, 11, 1303. https://doi.org/10.3390/nu11061303
Benedé S, Gradillas A, Villalba M, Batanero E. Allium porrum Extract Decreases Effector Cell Degranulation and Modulates Airway Epithelial Cell Function. Nutrients. 2019; 11(6):1303. https://doi.org/10.3390/nu11061303
Chicago/Turabian StyleBenedé, Sara, Ana Gradillas, Mayte Villalba, and Eva Batanero. 2019. "Allium porrum Extract Decreases Effector Cell Degranulation and Modulates Airway Epithelial Cell Function" Nutrients 11, no. 6: 1303. https://doi.org/10.3390/nu11061303
APA StyleBenedé, S., Gradillas, A., Villalba, M., & Batanero, E. (2019). Allium porrum Extract Decreases Effector Cell Degranulation and Modulates Airway Epithelial Cell Function. Nutrients, 11(6), 1303. https://doi.org/10.3390/nu11061303