Bovine κ-Casein Fragment Induces Hypo-Responsive M2-Like Macrophage Phenotype

Abstract
:1. Introduction
2. Experimental Section
2.1. Reagents and Materials
2.2. Animals and Ethics
2.3. Generation of Bone Marrow-Derived Macrophages
2.4. Generation of Monocyte Derived Human Macrophages
2.5. Cell Activation
2.6. CD4+ T-Cells Co-Culture
2.7. Flow Cytometry
2.8. Polymerase Chain Reaction
2.9. Sodium Dodecyl Sulphate Polyacrylamide Gel Electrophoresis
2.10. Cytotoxicity Assays
2.11. Statistical Analysis
3. Results
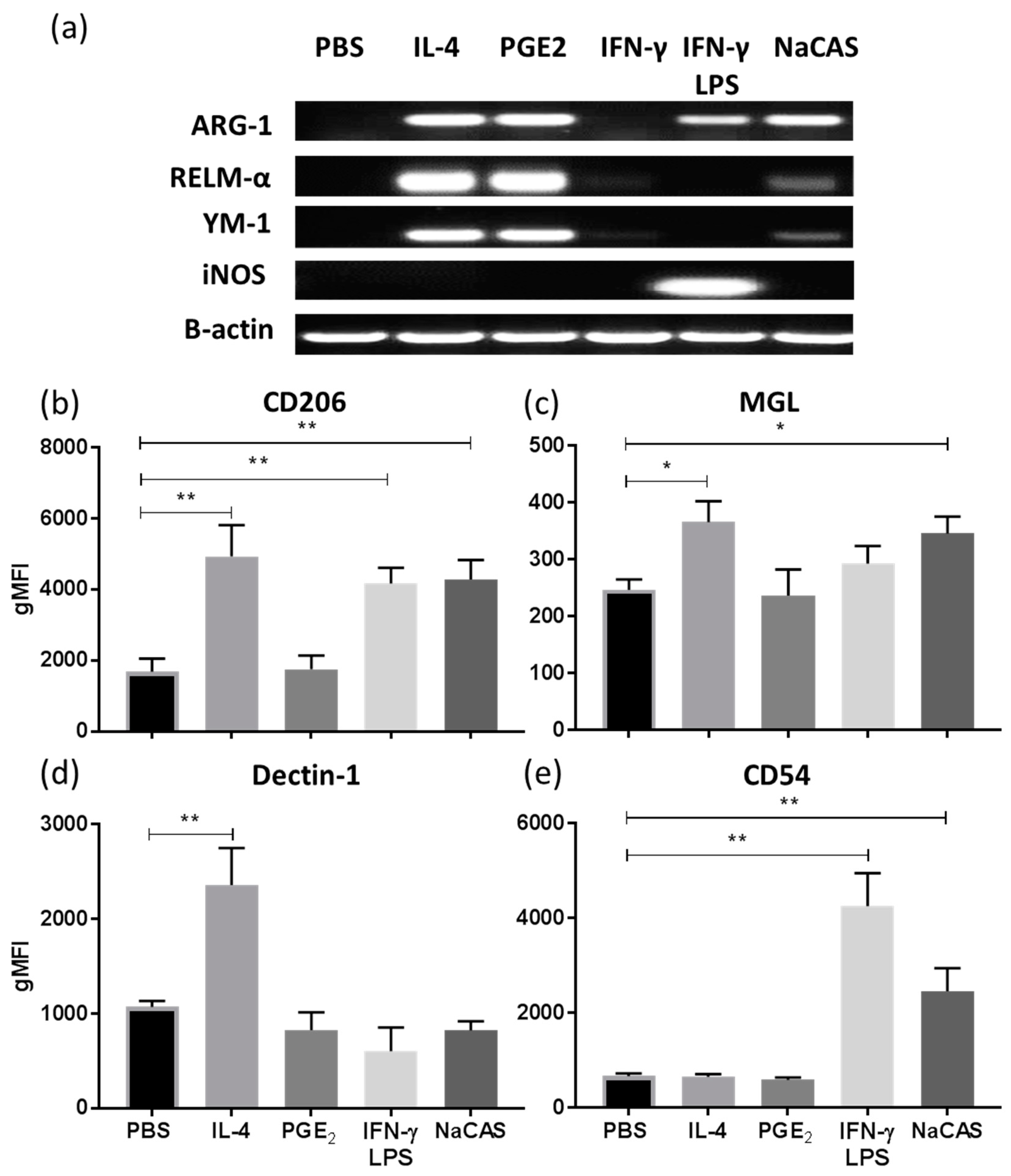
3.1. NaCAS Induces an M2-Like Macrophage Phenotype that Exhibits a Reduced Responsiveness to LPS Stimulation
3.2. κ-CAS Is the Subunit Responsible for the Suppression of LPS-Induced Cytokine Responses and the Induction of M2 Related Genes in BMMφ
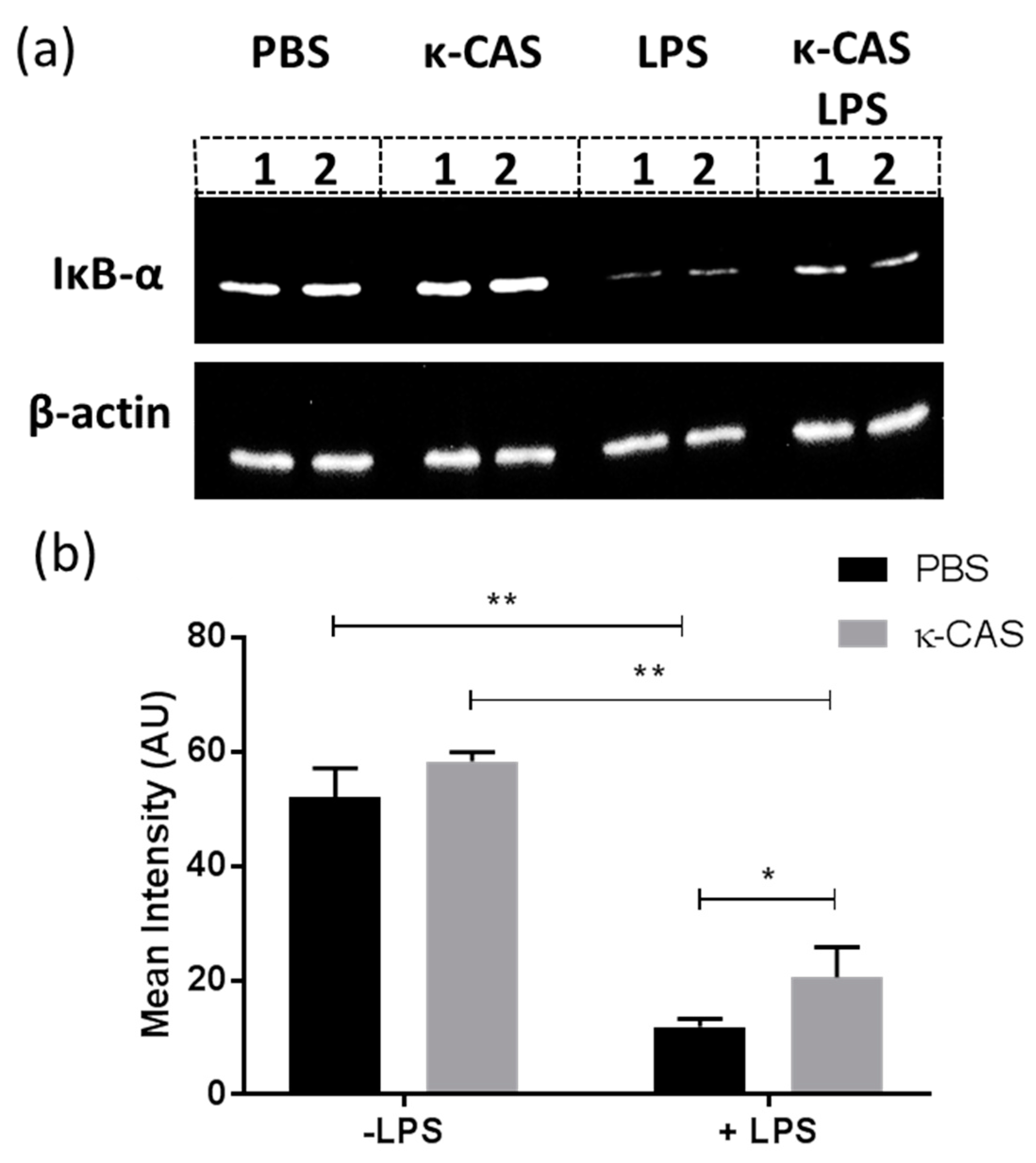
3.3. κ-CAS Abrogates NF-κB Activation
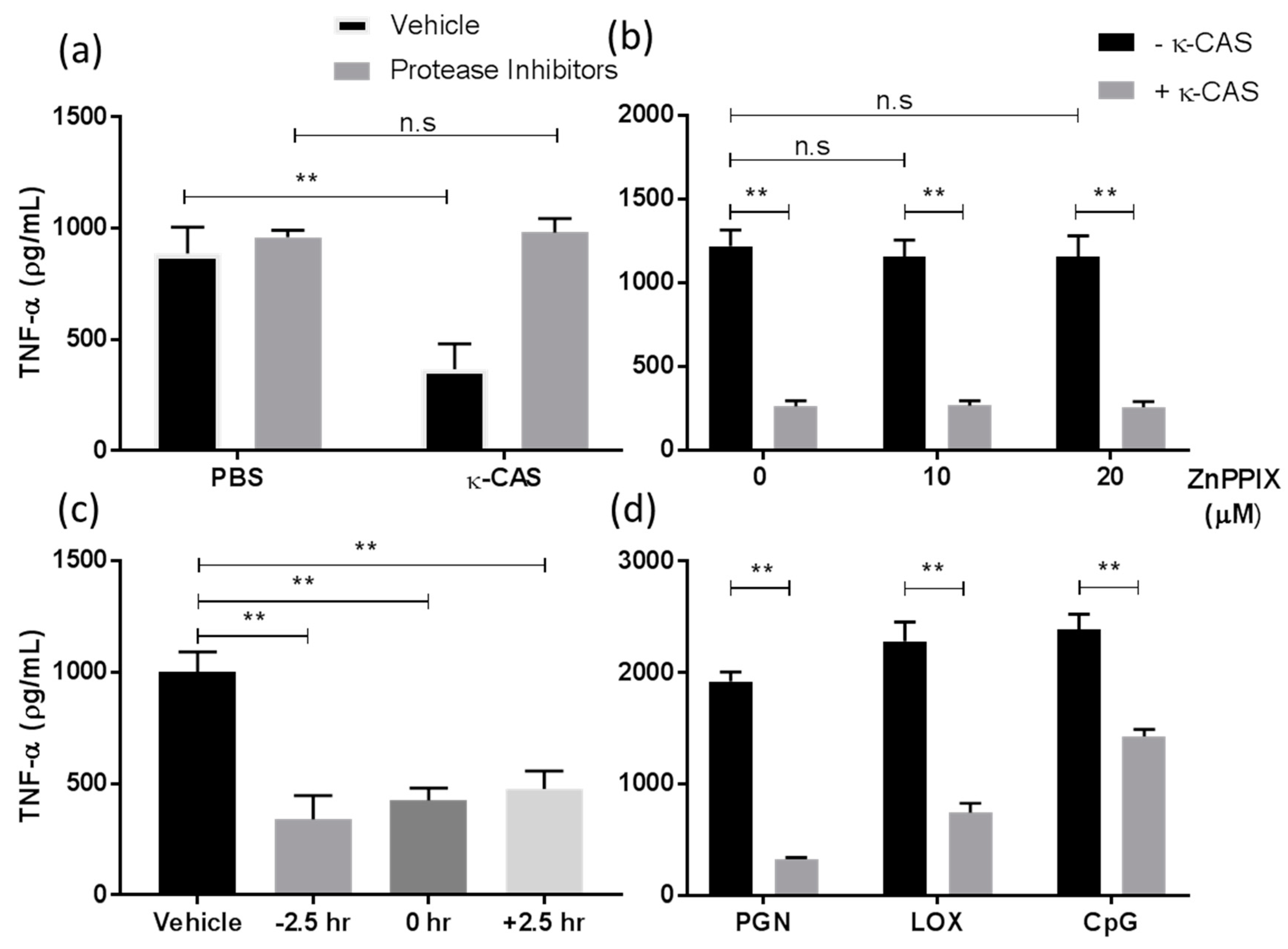
3.4. An Active Fragment of κ-CAS Is Responsible for Its Effect Which Targets Multiple TLRs Signalling Pathways
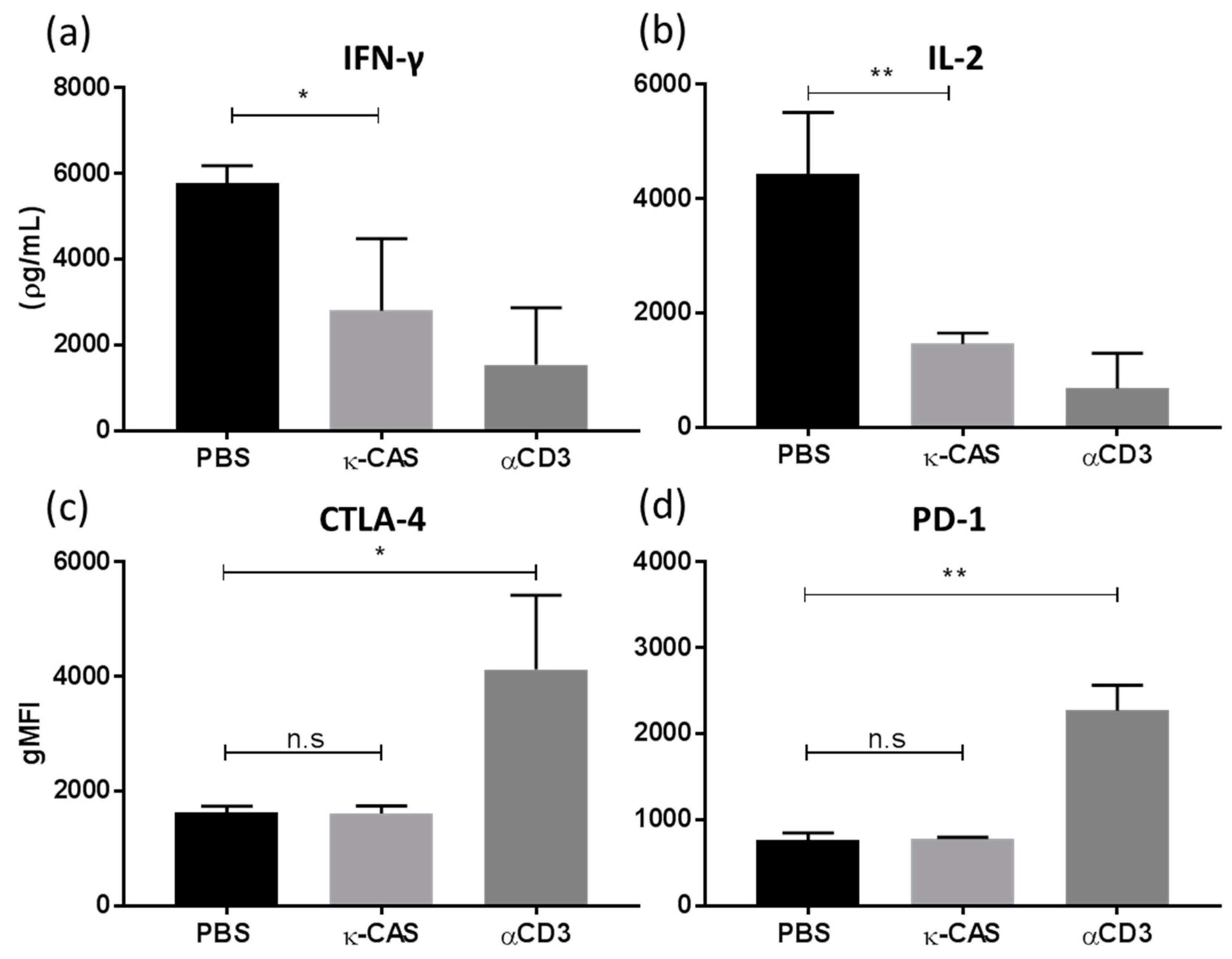
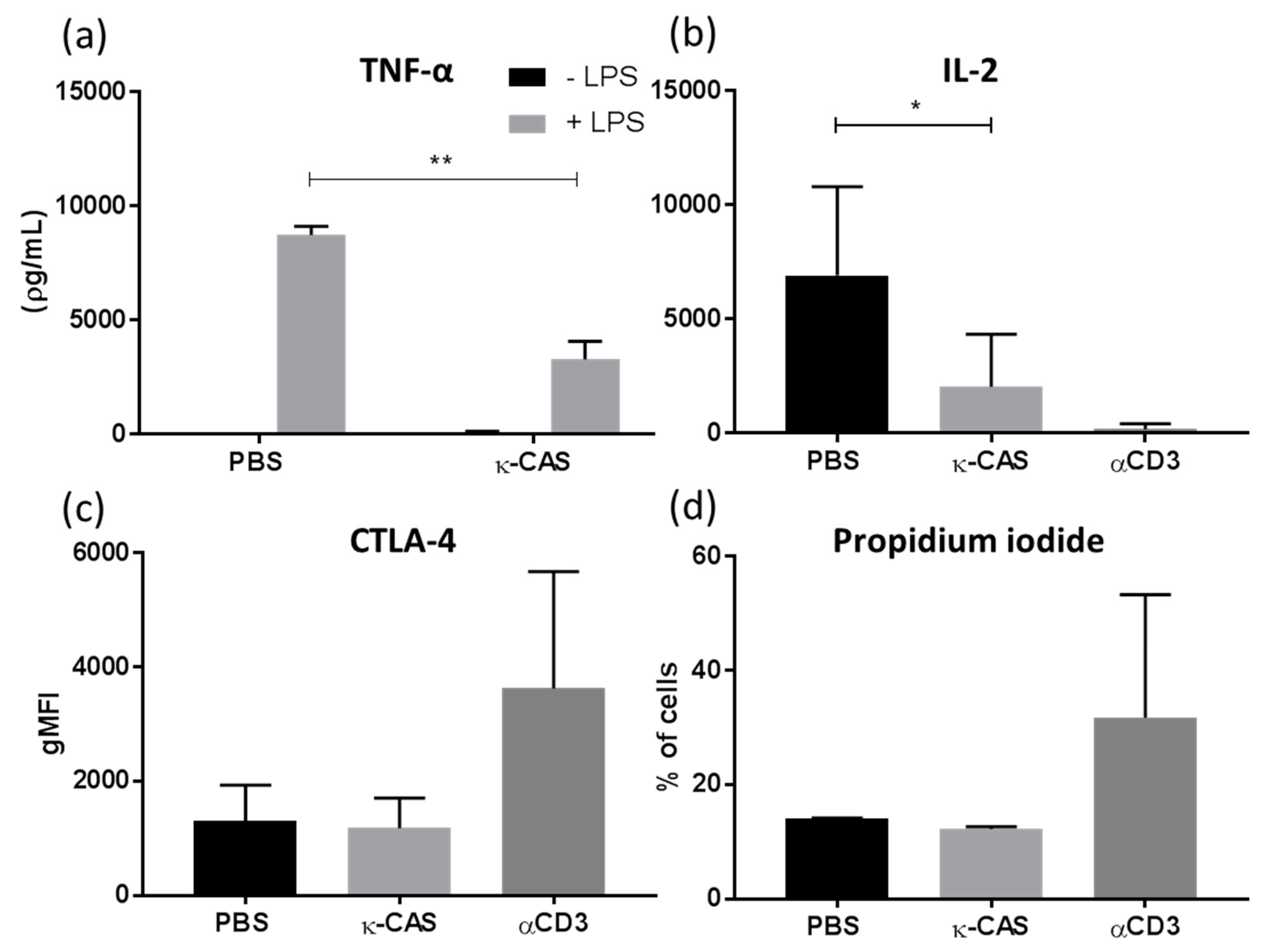
3.5. κ-CAS Sequesters the T-cell Priming Capacity of Macrophages
3.6. The Immunosuppressive Effects Exerted by κ-CAS Are Transferable in Human Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kalra, E.K. Nutraceutical--definition and introduction. AAPS PharmSci. 2003, 5, E25. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J. Nutraceuticals, nutritional therapy, phytonutrients, and phytotherapy for improvement of human health: A perspective on plant biotechnology application. Recent Pat. Biotechnol. 2007, 1, 75–97. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, D. The World Dairy Situation (2008)—By International Dairy Federation. Int. J. Dairy Technol. 2009, 62, 286–287. [Google Scholar] [CrossRef]
- Rutherfurd-Markwick, K.J.; Moughan, P.J. Bioactive peptides derived from food. J. AOAC Int. 2005, 88, 955–966. [Google Scholar] [PubMed]
- Dziuba, M.; Dziuba, B.; Iwaniak, A. A milk proteins as precursors of bioactive peptides. Acta Sci. Pol. Technol. Aliment. 2009, 8, 71–90. [Google Scholar] [CrossRef]
- Mohanty, D.P.; Mohapatra, S.; Misra, S.; Sahub, P.S. Milk derived bioactive peptides and their impact on human health—A review. Saudi J. Biol. Sci. 2016, 23, 577–583. [Google Scholar] [CrossRef]
- El-Gabalawy, H.; Guenther, L.C.; Bernstein, C.N. Epidemiology of immune-mediated inflammatory diseases: Incidence, prevalence, natural history, and comorbidities. J. Rheumatol. Suppl. 2010, 85, 2–10. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. The scientific evidence for the role of milk protein-derived bioactive peptides in humans. J. Funct. Foods 2015, 17, 640–656. [Google Scholar] [CrossRef]
- Gokhale, A.S.; Satyanarayanajois, S. Peptides and peptidomimetics as immunomodulators. Immunotherapy 2014, 6, 755–774. [Google Scholar] [CrossRef] [Green Version]
- Agyei, D.; Ongkudon, C.M.; Wei, C.Y.; Chan, A.S.; Danquah, M.K. Bioprocess challenges to the isolation and purification of bioactive peptides. Food Bioprod. Process. 2016, 98, 244–256. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Sahl, H.G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Marx, V. Watching peptides grow up. Chem. Eng. News 2005, 83, 17–24. [Google Scholar] [CrossRef]
- Mason, J.M. Design and development of peptides and peptide mimetics as antagonists for therapeutic intervention. Future Med. Chem. 2010, 2, 1813–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, H.S.; Doull, F.; Rutherfurd, K.J.; Cross, M.L. Immunoregulatory peptides in bovine milk. Br. J. Nutr. 2000, 84, S111–S117. [Google Scholar] [CrossRef] [PubMed]
- Requena, P.; Daddaoua, A.; Martínez-Plata, E.; González, M.; Zarzuelo, A.; Suárez, M.D.; Sánchez de Medina, F.; Martínez-Augustin, O. Bovine glycomacropeptide ameliorates experimental rat ileitis by mechanisms involving downregulation of interleukin 17. Br. J. Pharmacol. 2008, 154, 825–832. [Google Scholar] [CrossRef] [Green Version]
- López-Posadas, R.; Requena, P.; González, R.; Suárez, M.D.; Zarzuelo, A.; Sánchez de Medina, F.; Martínez-Augustin, O. Bovine glycomacropeptide has intestinal antiinflammatory effects in rats with dextran sulfate-induced colitis. J. Nutr. 2010, 140, 2014–2019. [Google Scholar] [CrossRef]
- Hvas, C.L.; Dige, A.; Bendix, M.; Wernlund, P.G.; Christensen, L.A.; Dahlerup, J.F.; Agnholt, J. Casein glycomacropeptide for active distal ulcerative colitis: A randomized pilot study. Eur. J. Clin. Investig. 2016, 46, 555–563. [Google Scholar] [CrossRef]
- Cobrin, G.M.; Abreu, M.T. Defects in mucosal immunity leading to Crohn’s disease. Immunol. Rev. 2005, 206, 277–295. [Google Scholar] [CrossRef]
- Bates, J.; Diehl, L. Dendritic cells in IBD pathogenesis: An area of therapeutic opportunity? J. Pathol. 2014, 232, 112–120. [Google Scholar] [CrossRef]
- Gren, S.T.; Grip, O. Role of Monocytes and Intestinal Macrophages in Crohn’s Disease and Ulcerative Colitis. Inflamm. Bowel Dis. 2016, 22, 1992–1998. [Google Scholar] [CrossRef]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Otani, H.; Futakami, M. Effects of Bovine Milk Proteins on the Phagocytic Property and Formation of Nitrite by Mouse Peritoneal Macrophages. Anim. Sci. Technol. 1994, 65, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Otani, H.; Futakami, M. Modification of nitrite production and phagocytosis of thioglycollate-elicited mouse peritoneal macrophages by Bovine Casein Digests. Food Agric. Immunol. 1996, 8, 59–69. [Google Scholar] [CrossRef]
- Li, T.; Cheng, X.; Du, M.; Chen, B.; Mao, X. Upregulation of heme oxygenase-1 mediates the anti-inflammatory activity of casein glycomacropeptide (GMP) hydrolysates in LPS-stimulated macrophages. Food Funct. 2017, 8, 2475–2484. [Google Scholar] [CrossRef] [PubMed]
- Ramos, G.; Weiss, B.; Córdova, Y.; Hernández, J.; Zambrano, I.; Santiago, E. Sodium caseinate induces expression and secretion of murine multipotent myeloid cell line 32D macrophage colony-stimulating factor. Arch. Med. Res. 2004, 35, 109–113. [Google Scholar] [CrossRef]
- Bamdad, F.; Shin, S.H.; Suh, J.W.; Nimalaratne, C.; Sunwoo, H. Anti-inflammatory and antioxidant properties of casein hydrolysate produced using high hydrostatic pressure combined with proteolytic enzymes. Molecules 2017, 22, 609. [Google Scholar] [CrossRef]
- Mikkelsen, T.L.; Bakman, S.; Sørensen, E.S.; Barkholt, V.; Frøkiaer, H. Sialic acid-containing milk proteins show differential immunomodulatory activities independent of sialic acid. J. Agric. Food Chem. 2005, 53, 7673–7680. [Google Scholar] [CrossRef]
- Mills, C.D.; Lenz, L.L.; Ley, K. Macrophages at the fork in the road to health or disease. Front. Immunol. 2015, 6, 59. [Google Scholar] [CrossRef]
- Weischenfeldt, J.; Porse, B. Bone marrow-derived macrophages (BMM): Isolation and applications. CSH Protoc. 2008. [Google Scholar] [CrossRef]
- Saghaeian-Jazi, M.; Mohammadi, S.; Sedighi, S. Culture and differentiation of monocyte derived macrophages using human serum: An optimized method. Zahedan J. Med Sci. 2016, 18, 29–32. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Dalgleish, D.G.; Law, A.J.R. Sodium caseinates? Composition and properties of different preparations. Int. J. Dairy Technol. 1998, 41, 1–4. [Google Scholar] [CrossRef]
- Lawrence, T. The Nuclear Factor NF-κB Pathway in Inflammation. CSH Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef]
- Monnai, M.; Otani, H. Effect of bovine kappa-caseinoglycopeptide on secretion of interleukin-1 family cytokines by P388D1 cells, a line derived from mouse monocyte/macrophage. Milchwissenshaft 1997, 52, 192–196. [Google Scholar]
- Cheng, X.; Gao, D.; Chen, B.; Mao, X. Endotoxin-binding peptides derived from casein glycomacropeptide inhibit lipopolysaccharide-stimulated inflammatory responses via blockade of NF-κB activation in macrophages. Nutrients 2015, 7, 3119–3137. [Google Scholar] [CrossRef] [PubMed]
- Hume, D.A. Macrophages as APC and the dendritic cell myth. J. Immunol. 2008, 181, 5829–5835. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Biswas, S.K.; Galdiero, M.R.; Sica, A.; Locati, M. Macrophage plasticity and polarization in tissue repair and remodelling. J. Pathol. 2013, 229, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, E.C.; Plevy, S.E. The role of macrophages and dendritic cells in the initiation of inflammation in IBD. Inflamm. Bowel Dis. 2014, 20, 166–175. [Google Scholar] [CrossRef]
- Stein, M.; Keshav, S.; Harris, N.; Gordon, S. Interleukin 4 potently enhances murine macrophage mannose receptor activity: A marker of alternative immunologic macrophage activation. J. Exp. Med. 1992, 176, 287–292. [Google Scholar] [CrossRef]
- Espagnolle, N.; Balguerie, A.; Arnaud, E.; Sensebé, L.; Varin, A. CD54-mediated interaction with pro-inflammatory macrophages increases the immunosuppressive function of human mesenchymal stromal cells. Stem Cell Rep. 2017, 8, 961–976. [Google Scholar] [CrossRef]
- Chirathaworn, C.; Kohlmeier, J.E.; Tibbetts, S.A.; Rumsey, L.M.; Chan, M.A.; Benedict, S.H. Stimulation through intercellular adhesion molecule-1 provides a second signal for T cell activation. J. Immunol. 2002, 168, 5530–5537. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage activation and polarization: Nomenclature and experimental guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Yao, L.; Li, L.; Zhang, J.; Place, A.T.; Minshall, R.D.; Liu, G. ICAM-1 regulates macrophage polarization by suppressing MCP-1 expression via miR-124 upregulation. Oncotarget 2017, 8, 111882–111901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheikh, N.A.; Jones, L.A. CD54 is a surrogate marker of antigen presenting cell activation. Cancer Immunol. Immunother. 2008, 57, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Soldano, S.; Pizzorni, C.; Paolino, S.; Trombetta, A.C.; Montagna, P.; Brizzolara, R.; Ruaro, B.; Sulli, A.; Cutolo, M. Alternatively activated (M2) macrophage phenotype is inducible by endothelin-1 in cultured human macrophages. PLoS ONE 2016, 11, e0166433. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, S.; O’Neill, S.M.; Sekiya, M.; Mulcahy, G.; Dalton, J.P. Thioredoxin peroxidase secreted by Fasciola hepatica induces the alternative activation of macrophages. Infect. Immun. 2005, 73, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.N.; Aldridge, A.; Vukman, K.V.; Donnelly, S.; O’Neill, S.M. Fasciola hepatica tegumental antigens indirectly induce an M2 macrophage-like phenotype in vivo. Parasite Immunol. 2014, 36, 531–539. [Google Scholar] [CrossRef]
- Mukhopadhya, A.; Noronha, N.; Bahar, B.; Ryan, M.T.; Murray, B.A.; Kelly, P.M.; Sweeney, T. Anti-inflammatory effects of a casein hydrolysate and its peptide-enriched fractions on TNFα-challenged Caco-2 cells and LPS-challenged porcine colonic explants. Food Sci. Nutr. 2014, 2, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Kiewiet, M.B.G.; Dekkers, R.; Gros, M.; van Neerven, R.J.J.; Groeneveld, A.; de Vos, P.; Faas, M.M. Toll-like receptor mediated activation is possibly involved in immunoregulating properties of cow’s milk hydrolysates. PLoS ONE 2017, 12, e0178191. [Google Scholar] [CrossRef]
- Bradley, J.R. TNF-mediated inflammatory disease. J. Pathol. 2008, 214, 149–160. [Google Scholar] [CrossRef]
- Wong, C.W.; Seow, H.F.; Liu, A.H.; Husband, A.J.; Smithers, G.W.; Watson, D.L. Modulation of immune responses by bovine beta-casein. Immunol. Cell Biol. 1996, 74, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Juillerat-Jeanneret, L.; Robert, M.C.; Juillerat, M.A. Peptides from Lactobacillus hydrolysates of bovine milk caseins inhibit prolylpeptidases of human colon cells. J. Agric. Food Chem. 2011, 59, 370–377. [Google Scholar] [CrossRef]
- Chatterton, D.E.W.; Nguyen, D.N.; Bering, S.B.; Sangild, P.T. Anti-inflammatory mechanisms of bioactive milk proteins in the intestine of newborns. Int. J. Biochem. Cell Biol. 2013, 45, 1730–1747. [Google Scholar] [CrossRef]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Smeltz, R.B.; Chen, J.; Ehrhardt, R.; Shevach, E.M. Role of IFN-gamma in Th1 differentiation: IFN-gamma regulates IL-18R alpha expression by preventing the negative effects of IL-4 and by inducing/maintaining IL-12 receptor beta 2 expression. J. Immunol. 2002, 168, 6165–6172. [Google Scholar] [CrossRef]
- Croft, M.; So, T.; Duan, W.; Soroosh, P. The significance of OX40 and OX40L to T-cell biology and immune disease. Immunol. Rev. 2009, 229, 173–191. [Google Scholar] [CrossRef] [Green Version]
- Grewal, I.S.; Flavell, R.A. CD40 and CD154 in cell-mediated immunity. Annu. Rev. Immunol. 1998, 16, 111–135. [Google Scholar] [CrossRef] [PubMed]
- Karulf, M.; Kelly, A.; Weinberg, A.D.; Gold, J.A. OX40 ligand regulates inflammation and mortality in the innate immune response to sepsis. J. Immunol. 2010, 185, 4856–4862. [Google Scholar] [CrossRef]
- Vukman, K.V.; Adams, P.N.; Metz, M.; Maurer, M.; O’Neill, S.M. Fasciola hepatica tegumental coat impairs mast cells’ ability to drive Th1 immune responses. J. Immunol. 2013, 190, 2873–2879. [Google Scholar] [CrossRef]
- Aldridge, A.; O’Neill, S.M. Fasciola hepatica tegumental antigens induce anergic-like T cells via dendritic cells in a mannose receptor-dependent manner. Eur. J. Immunol. 2016, 46, 1180–1192. [Google Scholar] [CrossRef]
- Lutz, M.B.; Schuler, G. Immature, semi-mature and fully mature dendritic cells: Which signals induce tolerance or immunity? Trends Immunol. 2002, 23, 445–449. [Google Scholar] [CrossRef]
- Wells, A.D.; Walsh, M.C.; Bluestone, J.A.; Turka, L.A. Signaling through CD28 and CTLA-4 controls two distinct forms of T cell anergy. J. Clin. Investig. 2001, 108, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Buchbinder, E.I.; Desai, A. CTLA-4 and PD-1 pathways: Similarities, differences, and implications of their inhibition. Am. J. Clin. Oncol. 2016, 39, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Otani, H.; Monnai, M.; Kawasaki, Y.; Kawakami, H.; Tanimoto, M. Inhibition of mitogen-induced proliferative responses of lymphocytes by bovine kappa-caseinoglycopeptides having different carbohydrate chains. J. Dairy Res. 1995, 62, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Otani, H.; Hata, I. Inhibition of proliferative responses of mouse spleen lymphocytes and rabbit Peyer’s patch cells by bovine milk caseins and their digests. J. Dairy Res. 1995, 62, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.F.; Oxenius, A. Interleukin 2: From immunostimulation to immunoregulation and back again. EMBO Rep. 2007, 8, 1142–1148. [Google Scholar] [CrossRef]
- Li, E.W.Y.; Mine, Y. Immunoenhancing effects of bovine glycomacropeptide and its derivatives on the proliferative response andphagocytic activities of human macrophage like cells U937. J. Agric. Food Chem. 2004, 52, 2704–2708. [Google Scholar] [CrossRef]
- Gong, J.; Chen, Q.; Yan, Y.; Pang, G. Effect of casein glycomacropeptide on subunit p65 of nuclear transcription factor-κB in lipopolysaccharide-stimulated human colorectal tumor HT-29 cells. Food Sci. Hum. Wellness 2014, 3, 51–55. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sense | Anti-Sense |
|---|---|---|
| Arg-1 | CAGAAGAATGGAAGAGTCAG | CAGATATGCAGG GAGTCACC |
| Ym-1 | TCACAGGTCTGGCAATTCTTCTG | TTTGTCCTTAGGAGGGCTTCCTC |
| iNOS | CCCTTCCGAAGTTTCTGGCAGCAGC | GGCTGTCAGAGAGCCTCGTGGCTTTGG |
| RELMα | GGTCCCAGTGCATATGGATGAGACCATAGA | CACCTCTTCACTCGAGGGACAGTTGGCAGC |
| B-actin | TGGAATCCTGTGGCATCCATGAAAC | TAAAACGCAGCTCAGTAACAGTCCG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lalor, R.; O’Neill, S. Bovine κ-Casein Fragment Induces Hypo-Responsive M2-Like Macrophage Phenotype. Nutrients 2019, 11, 1688. https://doi.org/10.3390/nu11071688
Lalor R, O’Neill S. Bovine κ-Casein Fragment Induces Hypo-Responsive M2-Like Macrophage Phenotype. Nutrients. 2019; 11(7):1688. https://doi.org/10.3390/nu11071688
Chicago/Turabian StyleLalor, Richard, and Sandra O’Neill. 2019. "Bovine κ-Casein Fragment Induces Hypo-Responsive M2-Like Macrophage Phenotype" Nutrients 11, no. 7: 1688. https://doi.org/10.3390/nu11071688
APA StyleLalor, R., & O’Neill, S. (2019). Bovine κ-Casein Fragment Induces Hypo-Responsive M2-Like Macrophage Phenotype. Nutrients, 11(7), 1688. https://doi.org/10.3390/nu11071688