Greater Efficacy of Black Ginseng (CJ EnerG) over Red Ginseng against Lethal Influenza A Virus Infection

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of BG (CJ EnerG) Extract
2.2. Analysis of Acid Polysaccharides
2.3. Measurements of Ginsenosides
2.4. Virus
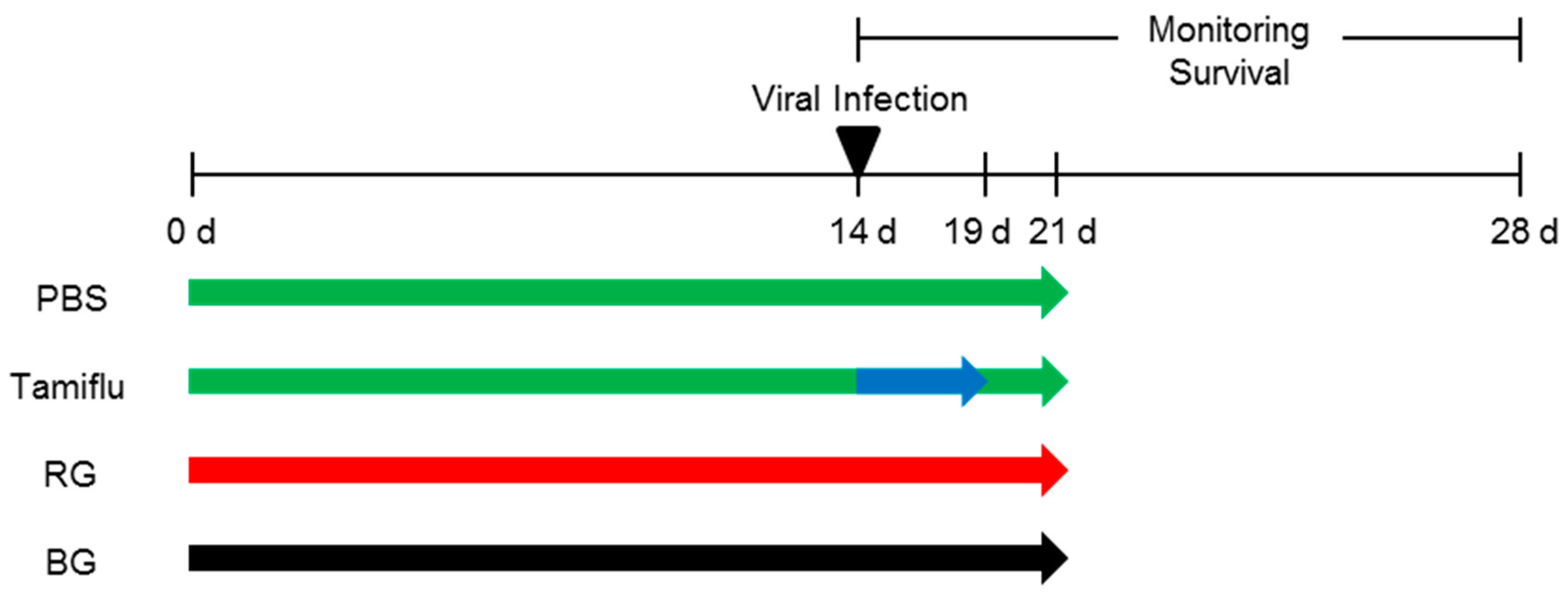
2.5. Mice and Treatments
2.6. Measurement of Survival
2.7. Determination of Lung Viral Titers
2.8. Histopathological Assays
2.9. Measurements of Cytokines
2.10. IgG Assay
2.11. Hemagglutination Inhibition (HI) Assay and Plaque Reduction Assay
2.12. Statistical Analysis
3. Results
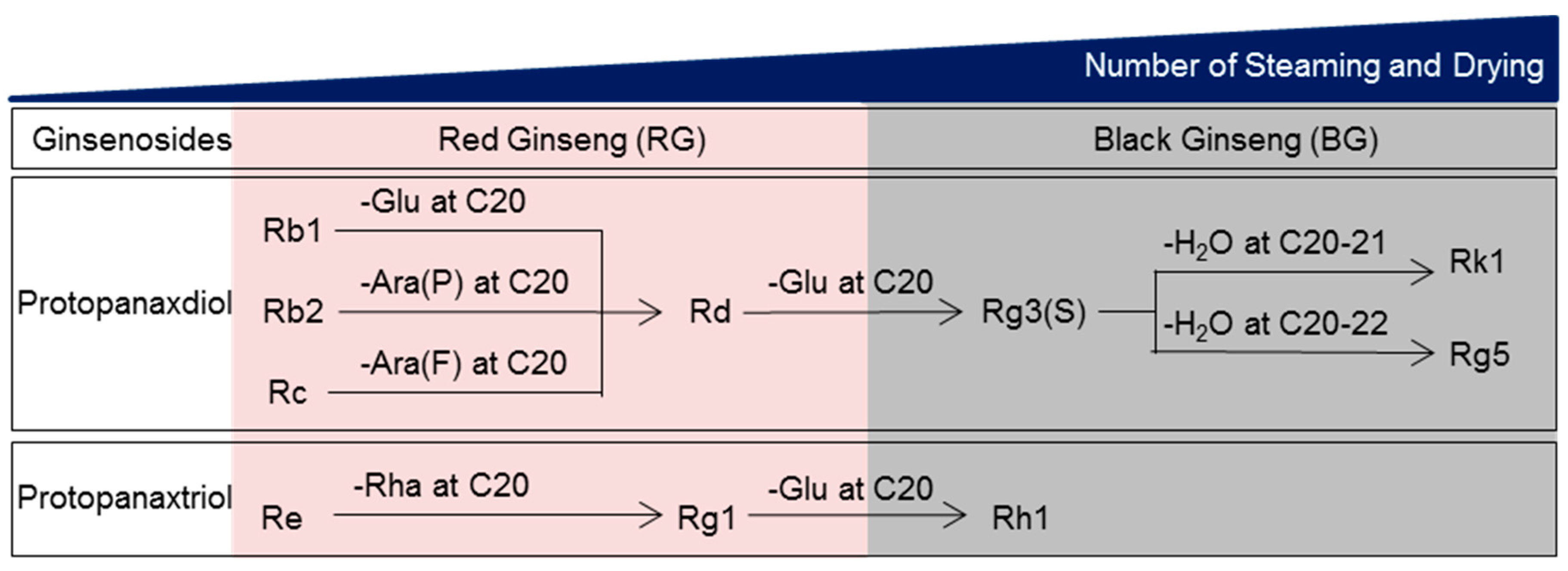
3.1. The Components of BG Differ from Those of RG
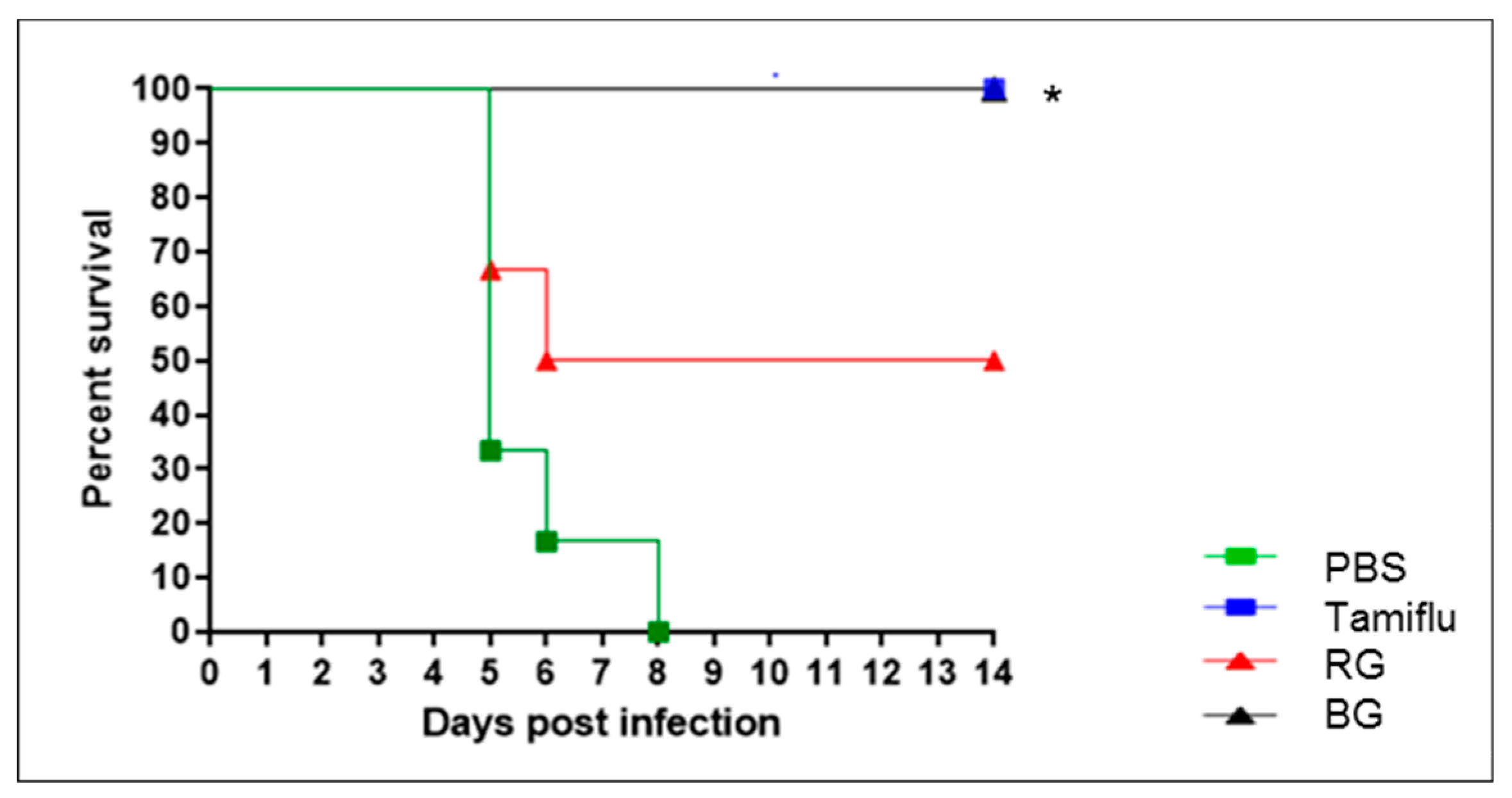
3.2. BG Protects Mice Against Lethal Influenza A Virus Infection
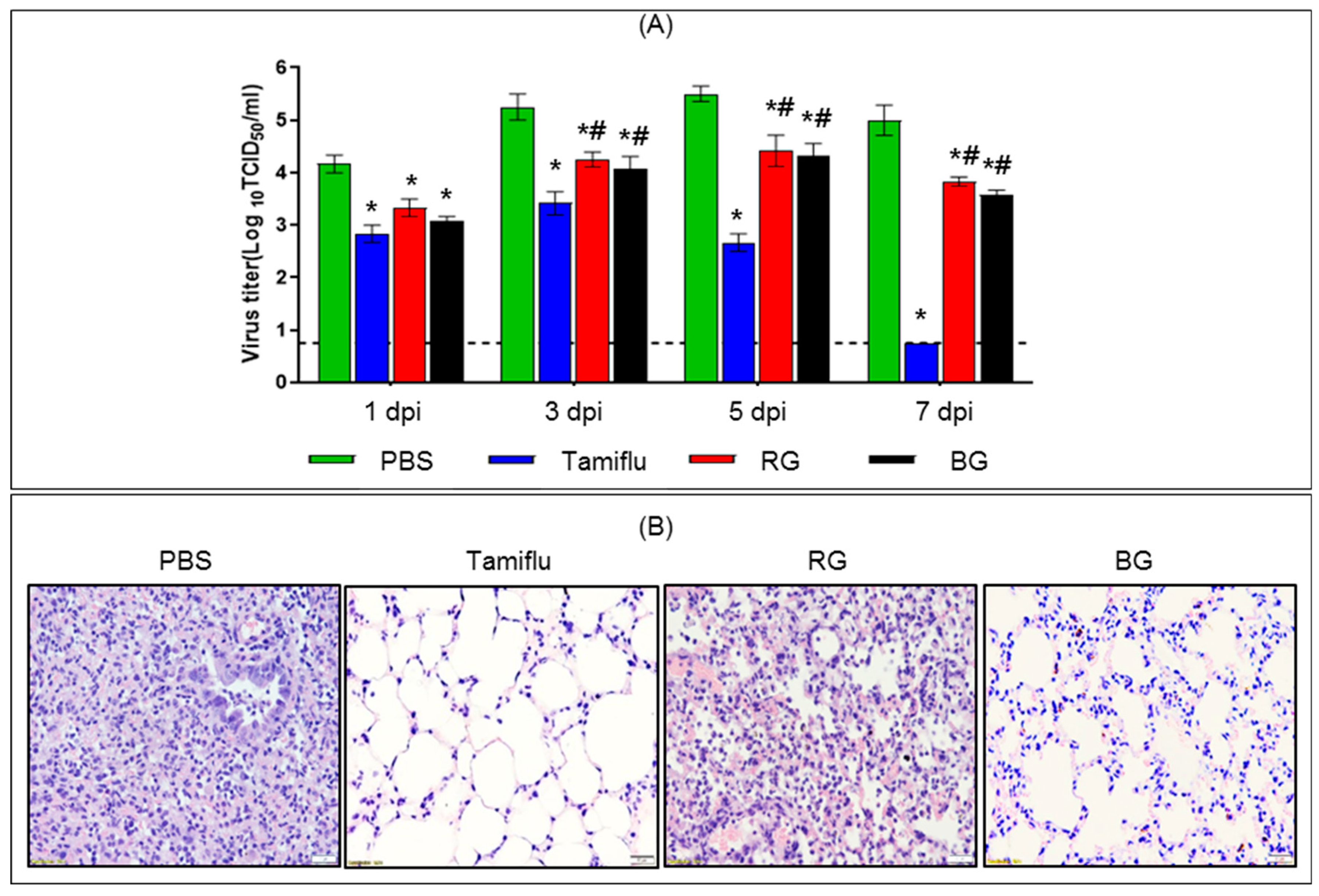
3.3. Administration of BG Results in Reduced Viral Burden and Lung Histopathology Following Lethal Influenza A Virus Infection
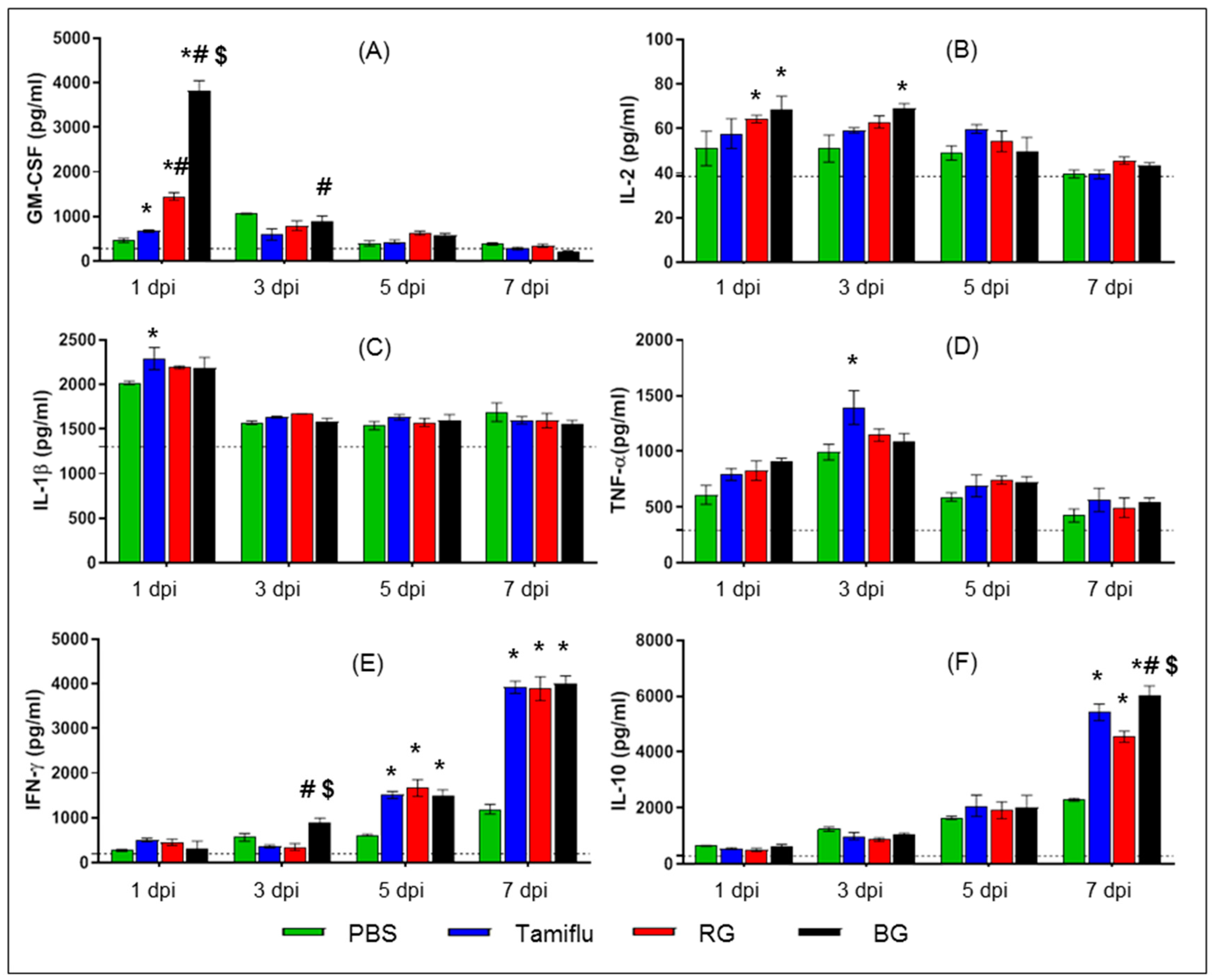
3.4. BG Induces Cytokine Production in Infected Mice
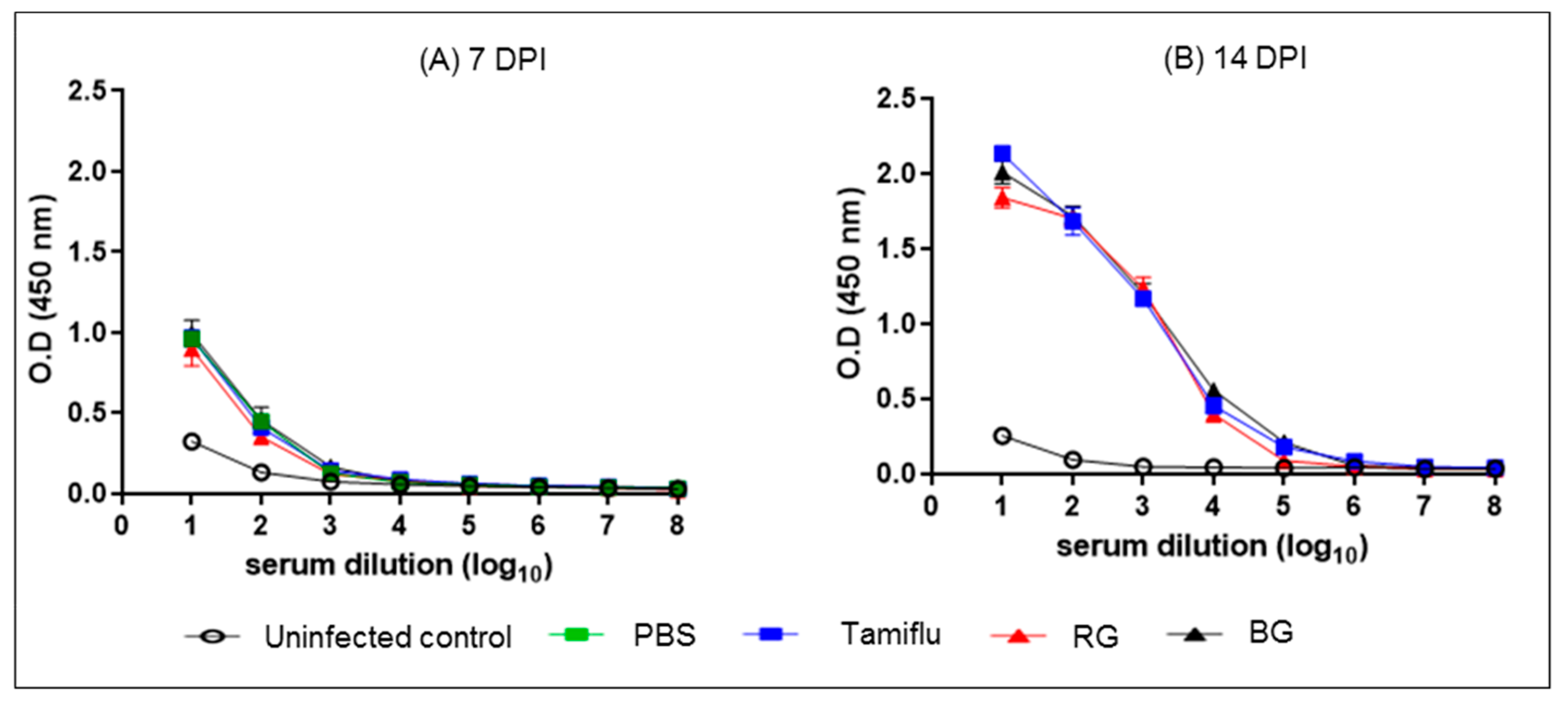
3.5. BG Does Not Affect the Development of Normal Influenza-Specific Antibodies
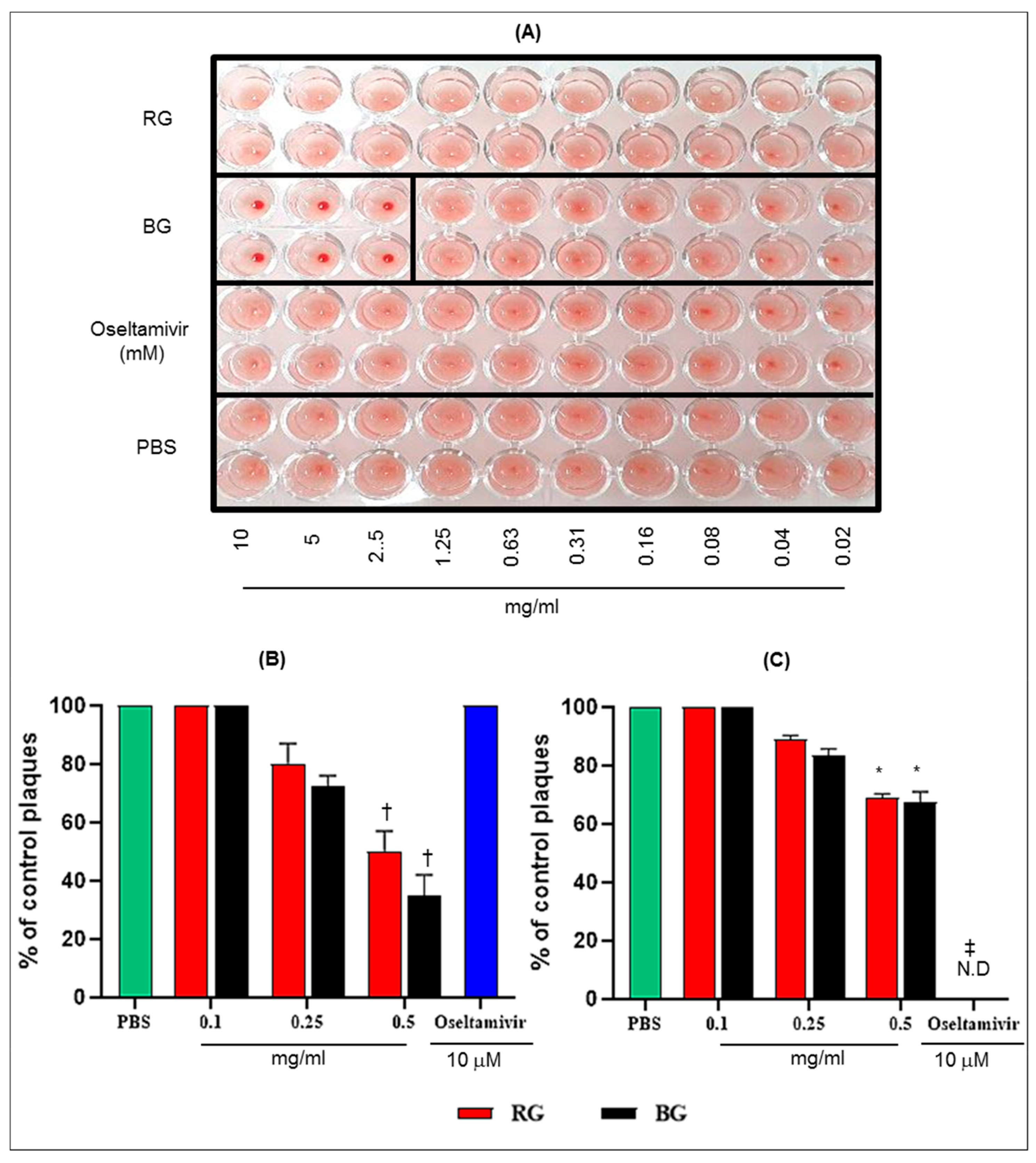
3.6. BG Inhibits the Hemagglutination Activity of Influenza A Virus and Virus Replication in Vitro
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Blut, A. Influenza virus. Transfus. Med. Hemother. 2009, 36, 32–39. [Google Scholar]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Mol. Biol. Rev. 1992, 56, 152–179. [Google Scholar]
- Girard, M.P.; Tam, J.S.; Assossou, O.M.; Kieny, M.P. The 2009 A (H1N1) influenza virus pandemic: A review. Vaccine 2010, 28, 4895–4902. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Vijaykrishna, D.; Bahl, J.; Zhu, H.; Wang, J.; Smith, G.J. The emergence of pandemic influenza viruses. Protein Cell 2010, 1, 9–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, G.J.; Bahl, J.; Vijaykrishna, D.; Zhang, J.; Poon, L.L.; Chen, H.; Webster, R.G.; Peiris, J.M.; Guan, Y. Dating the emergence of pandemic influenza viruses. Proc. Natl. Acad. Sci. USA 2009, 106, 11709–11712. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Blanton, L.; Elal, A.I.A.; Alabi, N.; Barnes, J.; Biggerstaff, M.; Brammer, L.; Budd, A.P.; Burns, E.; Cummings, C.N. Update: Influenza Activity in the United States During the 2018–19 Season and Composition of the 2019–20 Influenza Vaccine. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 544–551. [Google Scholar] [CrossRef]
- Lambert, L.C.; Fauci, A.S. Influenza vaccines for the future. N. Engl. J. Med. 2010, 363, 2036–2044. [Google Scholar] [CrossRef]
- Dreitlein, W.B.; Maratos, J.; Brocavich, J. Zanamivir and oseltamivir: Two new options for the treatment and prevention of influenza. Clin. Ther. 2001, 23, 327–355. [Google Scholar] [CrossRef]
- Gubareva, L.V.; Kaiser, L.; Hayden, F.G. Influenza virus neuraminidase inhibitors. Lancet 2000, 355, 827–835. [Google Scholar] [CrossRef]
- Moscona, A. Neuraminidase inhibitors for influenza. N. Engl. J. Med. 2005, 353, 1363–1373. [Google Scholar] [CrossRef]
- Wang, C.; Takeuchi, K.; Pinto, L.; Lamb, R. Ion channel activity of influenza A virus M2 protein: Characterization of the amantadine block. J. Virol. 1993, 67, 5585–5594. [Google Scholar]
- De Clercq, E. Antiviral agents active against influenza A viruses. Nat. Rev. Drug Discov. 2006, 5, 1015–1025. [Google Scholar] [CrossRef]
- Baranovich, T.; Wong, S.-S.; Armstrong, J.; Marjuki, H.; Webby, R.J.; Webster, R.G.; Govorkova, E.A. T-705 (favipiravir) induces lethal mutagenesis in influenza A H1N1 viruses In Vitro. J. Virol. 2013, 87, 3741–3751. [Google Scholar] [CrossRef]
- Jassim, S.A.; Naji, M.A. Novel antiviral agents: A medicinal plant perspective. J. Appl. Microbiol. 2003, 95, 412–427. [Google Scholar] [CrossRef]
- Kubo, T.; Nishimura, H. Antipyretic effect of Mao-to, a Japanese herbal medicine, for treatment of type A influenza infection in children. Phytomedicine 2007, 14, 96–101. [Google Scholar] [CrossRef]
- Wang, X.; Jia, W.; Zhao, A.; Wang, X. Anti-influenza agents from plants and traditional Chinese medicine. Phytother. Res. 2006, 20, 335–341. [Google Scholar] [CrossRef]
- Kim, E.-H.; Pascua, P.N.Q.; Song, M.-S.; Baek, Y.H.; Kwon, H.-I.; Park, S.-J.; Lim, G.-J.; mi Kim, S.; Decano, A.; Lee, K.J. Immunomodulaton and attenuation of lethal influenza A virus infection by oral administration with KIOM-C. Antivir. Res. 2013, 98, 386–393. [Google Scholar] [CrossRef]
- Dong, W.; Farooqui, A.; Leon, A.J.; Kelvin, D.J. Inhibition of influenza A virus infection by ginsenosides. PLoS ONE 2017, 12, e0171936. [Google Scholar] [CrossRef]
- Lee, J.; Hwang, H.; Ko, E.-J.; Lee, Y.-N.; Kwon, Y.-M.; Kim, M.-C.; Kang, S.-M. Immunomodulatory activity of red ginseng against influenza A virus infection. Nutrients 2014, 6, 517–529. [Google Scholar] [CrossRef]
- Wang, Y.; Jung, Y.-J.; Kim, K.-H.; Kwon, Y.; Kim, Y.-J.; Zhang, Z.; Kang, H.-S.; Wang, B.-Z.; Quan, F.-S.; Kang, S.-M. Antiviral activity of fermented ginseng extracts against a broad range of influenza viruses. Viruses 2018, 10, 471. [Google Scholar] [CrossRef]
- Xu, M.L.; Kim, H.J.; Choi, Y.R.; Kim, H.-J. Intake of Korean red ginseng extract and saponin enhances the protection conferred by vaccination with inactivated influenza A virus. J. Ginseng Res. 2012, 36, 396–402. [Google Scholar] [CrossRef]
- Yin, S.Y.; Kim, H.J.; Kim, H.-J. A comparative study of the effects of whole red ginseng extract and polysaccharide and saponin fractions on influenza A (H1N1) virus infection. Biol. Pharm. Bull. 2013, 36, 1002–1007. [Google Scholar] [CrossRef]
- Yoo, D.-G.; Kim, M.-C.; Park, M.-K.; Park, K.-M.; Quan, F.-S.; Song, J.-M.; Wee, J.J.; Wang, B.-Z.; Cho, Y.-K.; Compans, R.W. Protective effect of ginseng polysaccharides on influenza viral infection. PLoS ONE 2012, 7, e33678. [Google Scholar] [CrossRef]
- Lee, C.-S.; Lee, J.-H.; Oh, M.; Choi, K.-M.; Jeong, M.R.; Park, J.-D.; Kwon, D.Y.; Ha, K.-C.; Park, E.-O.; Lee, N. Preventive effect of Korean red ginseng for acute respiratory illness: A randomized and double-blind clinical trial. J. Korean Med. Sci. 2012, 27, 1472–1478. [Google Scholar] [CrossRef]
- Jin, Y.; Kim, Y.-J.; Jeon, J.-N.; Wang, C.; Min, J.-W.; Noh, H.-Y.; Yang, D.-C. Effect of white, red and black ginseng on physicochemical properties and ginsenosides. Plant. Food Hum. Nutr. 2015, 70, 141–145. [Google Scholar] [CrossRef]
- Xu, X.-F.; Gao, Y.; Xu, S.-Y.; Liu, H.; Xue, X.; Zhang, Y.; Zhang, H.; Liu, M.-N.; Xiong, H.; Lin, R.-C. Remarkable impact of steam temperature on ginsenosides transformation from fresh ginseng to red ginseng. J. Ginseng Res. 2018, 42, 277–287. [Google Scholar] [CrossRef]
- Yapo, B.M.; Koffi, K.L. Utilisation of model pectins reveals the effect of demethylated block size frequency on calcium gel formation. Carbohydr. Polym. 2013, 92, 1–10. [Google Scholar] [CrossRef]
- Leed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar]
- Yoo, D.-G.; Kim, M.-C.; Park, M.-K.; Song, J.-M.; Quan, F.-S.; Park, K.-M.; Cho, Y.-K.; Kang, S.-M. Protective effect of Korean red ginseng extract on the infections by H1N1 and H3N2 influenza viruses in mice. J. Med. Food 2012, 15, 855–862. [Google Scholar] [CrossRef]
- Baskin, C.R.; Bielefeldt-Ohmann, H.; Tumpey, T.M.; Sabourin, P.J.; Long, J.P.; García-Sastre, A.; Tolnay, A.-E.; Albrecht, R.; Pyles, J.A.; Olson, P.H. Early and sustained innate immune response defines pathology and death in nonhuman primates infected by highly pathogenic influenza virus. Proc. Natl. Acad. Sci. USA 2009, 106, 3455–3460. [Google Scholar] [CrossRef] [Green Version]
- Kash, J.C.; Tumpey, T.M.; Proll, S.C.; Carter, V.; Perwitasari, O.; Thomas, M.J.; Basler, C.F.; Palese, P.; Taubenberger, J.K.; García-Sastre, A. Genomic analysis of increased host immune and cell death responses induced by 1918 influenza virus. Nature 2006, 443, 578–581. [Google Scholar] [CrossRef]
- Huang, F.-F.; Barnes, P.F.; Feng, Y.; Donis, R.; Chroneos, Z.C.; Idell, S.; Allen, T.; Perez, D.R.; Whitsett, J.A.; Dunussi-Joannopoulos, K. GM-CSF in the lung protects against lethal influenza infection. Am. Thorac. Soc. 2011, 184, 259–268. [Google Scholar] [CrossRef]
- Rouse, B.T.; Sehrawat, S. Immunity and immunopathology to viruses: What decides the outcome? Nat. Rev. Immunol. 2010, 10, 514–526. [Google Scholar] [CrossRef]
- Chen, X.; Liu, S.; Goraya, M.U.; Maarouf, M.; Huang, S.; Chen, J.-L. Host immune response to influenza A virus infection. Front. Immunol. 2018, 9, 320–333. [Google Scholar] [CrossRef]
- Rojas, J.M.; Avia, M.; Martín, V.; Sevilla, N. IL-10: A multifunctional cytokine in viral infections. J. Immunol. Res. 2017, 2017, 6104054–6104068. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Z.; Liu, J.; Wang, Y.; Zhou, Q.; Wang, S.; Wang, X. Ginsenoside Rg3 improves cyclophosphamide-induced immunocompetence in Balb/c mice. Int. Immunopharmacol. 2019, 72, 98–111. [Google Scholar] [CrossRef]
- Yang, J.; Li, S.; Wang, L.; Du, F.; Zhou, X.; Song, Q.; Zhao, J.; Fang, R. Ginsenoside Rg3 attenuates lipopolysaccharide-induced acute lung injury via MerTK-dependent activation of the PI3K/AKT/mTOR pathway. Front. Pharmacol. 2018, 9, 850–863. [Google Scholar] [CrossRef]
- Moscona, A. Global transmission of oseltamivir-resistant influenza. N. Engl. J. Med. 2009, 360, 953–956. [Google Scholar] [CrossRef]
- Carey, B.; Trapnell, B.C. The molecular basis of pulmonary alveolar proteinosis. Clin. Immunol. 2010, 135, 223–235. [Google Scholar] [CrossRef] [Green Version]
- Shibata, Y.; Berclaz, P.-Y.; Chroneos, Z.C.; Yoshida, M.; Whitsett, J.A.; Trapnell, B.C. GM-CSF regulates alveolar macrophage differentiation and innate immunity in the lung through PU. 1. Immunity 2001, 15, 557–567. [Google Scholar] [CrossRef]
- Halstead, E.S.; Umstead, T.M.; Davies, M.L.; Kawasawa, Y.I.; Silveyra, P.; Howyrlak, J.; Yang, L.; Guo, W.; Hu, S.; Hewage, E.K. GM-CSF overexpression after influenza a virus infection prevents mortality and moderates M1-like airway monocyte/macrophage polarization. Respir. Res. 2018, 19, 3–17. [Google Scholar] [CrossRef]
- Sever-Chroneos, Z.; Murthy, A.; Davis, J.; Florence, J.M.; Kurdowska, A.; Krupa, A.; Tichelaar, J.W.; White, M.R.; Hartshorn, K.L.; Kobzik, L. GM-CSF modulates pulmonary resistance to influenza A infection. Antivir. Res. 2011, 92, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Subramaniam, R.; Hillberry, Z.; Chen, H.; Feng, Y.; Fletcher, K.; Neuenschwander, P.; Shams, H. Delivery of GM-CSF to protect against influenza pneumonia. PLoS ONE 2015, 10, e0124593. [Google Scholar] [CrossRef]
- Kim, M.K.; Lee, J.W.; Lee, K.Y.; Yang, D.-C. Microbial conversion of major ginsenoside Rb 1 to pharmaceutically active minor ginsenoside Rd. J. Microbiol. 2005, 43, 456–462. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acid Polysaccharides | Ginsenosides | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Rb1 | Rb2 | Rc | Rd | Re | Rg1 | Rg3(S) | Rk1 | Rg5 | Rh1 | ||
| m/g Extract | |||||||||||
| RG 1 | 0.37 | 4.91 | 2.21 | 3.23 | 1.75 | 3.74 | 1.29 | 0.42 | 0.12 | 0.46 | 0.43 |
| BG 2 | 2.63 | 0.83 | ND 3 | ND | ND | 1.34 | ND | 4.12 | 4.75 | 4.54 | 0.94 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.-H.; Kim, S.-W.; Park, S.-J.; Kim, S.; Yu, K.-M.; Kim, S.G.; Lee, S.H.; Seo, Y.-K.; Cho, N.-H.; Kang, K.; et al. Greater Efficacy of Black Ginseng (CJ EnerG) over Red Ginseng against Lethal Influenza A Virus Infection. Nutrients 2019, 11, 1879. https://doi.org/10.3390/nu11081879
Kim E-H, Kim S-W, Park S-J, Kim S, Yu K-M, Kim SG, Lee SH, Seo Y-K, Cho N-H, Kang K, et al. Greater Efficacy of Black Ginseng (CJ EnerG) over Red Ginseng against Lethal Influenza A Virus Infection. Nutrients. 2019; 11(8):1879. https://doi.org/10.3390/nu11081879
Chicago/Turabian StyleKim, Eun-Ha, Son-Woo Kim, Su-Jin Park, Semi Kim, Kwang-Min Yu, Seong Gyu Kim, Seung Hun Lee, Yong-Ki Seo, Nam-Hoon Cho, Kimoon Kang, and et al. 2019. "Greater Efficacy of Black Ginseng (CJ EnerG) over Red Ginseng against Lethal Influenza A Virus Infection" Nutrients 11, no. 8: 1879. https://doi.org/10.3390/nu11081879
APA StyleKim, E. -H., Kim, S. -W., Park, S. -J., Kim, S., Yu, K. -M., Kim, S. G., Lee, S. H., Seo, Y. -K., Cho, N. -H., Kang, K., Soung, D. Y., & Choi, Y. -K. (2019). Greater Efficacy of Black Ginseng (CJ EnerG) over Red Ginseng against Lethal Influenza A Virus Infection. Nutrients, 11(8), 1879. https://doi.org/10.3390/nu11081879