Glycated Beef Protein Hydrolysates as Sources of Bitter Taste Modifiers



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Maillard Reaction
2.3. Degree of Glycation (DG)
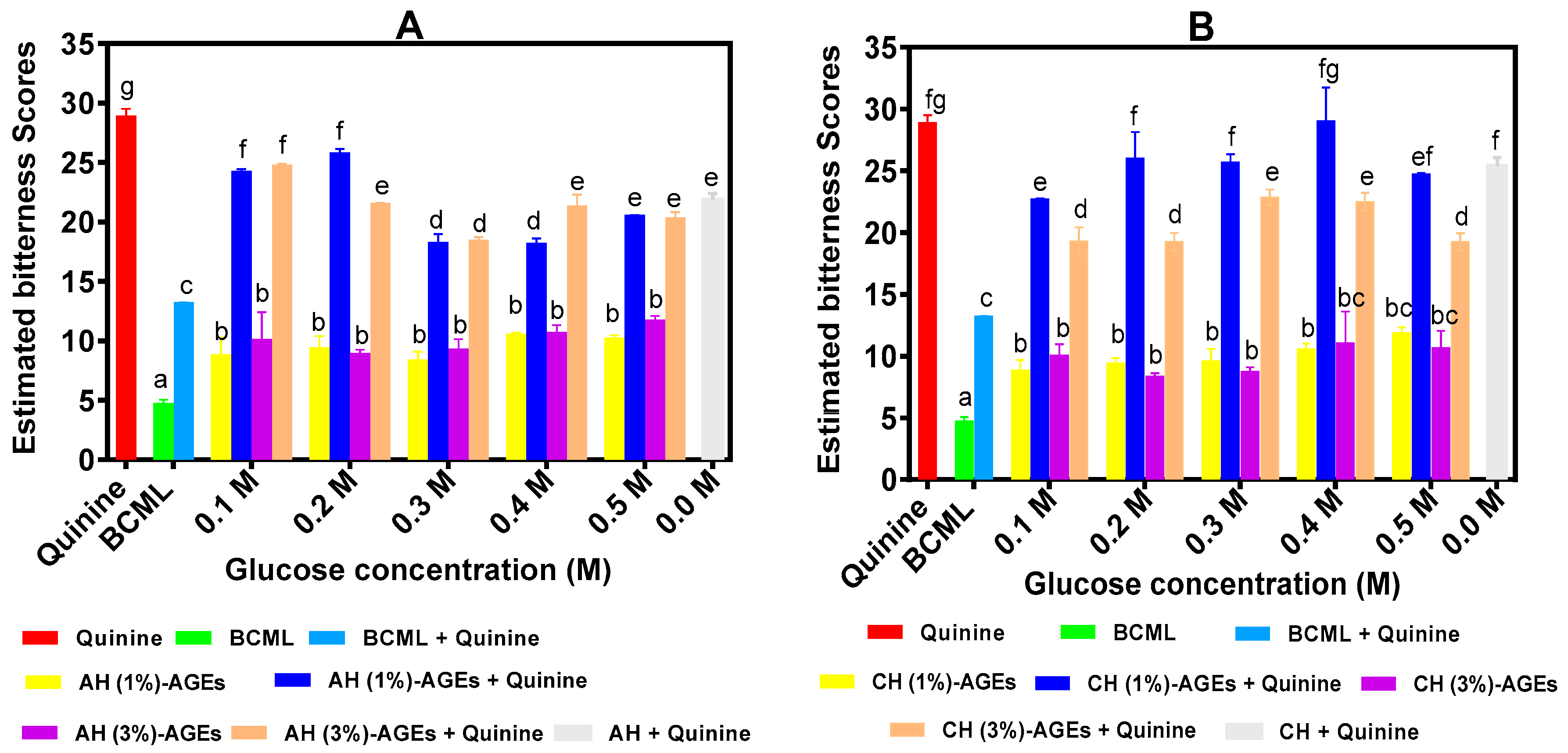
2.4. Estimation of Bitter Scores by Electronic Tongue
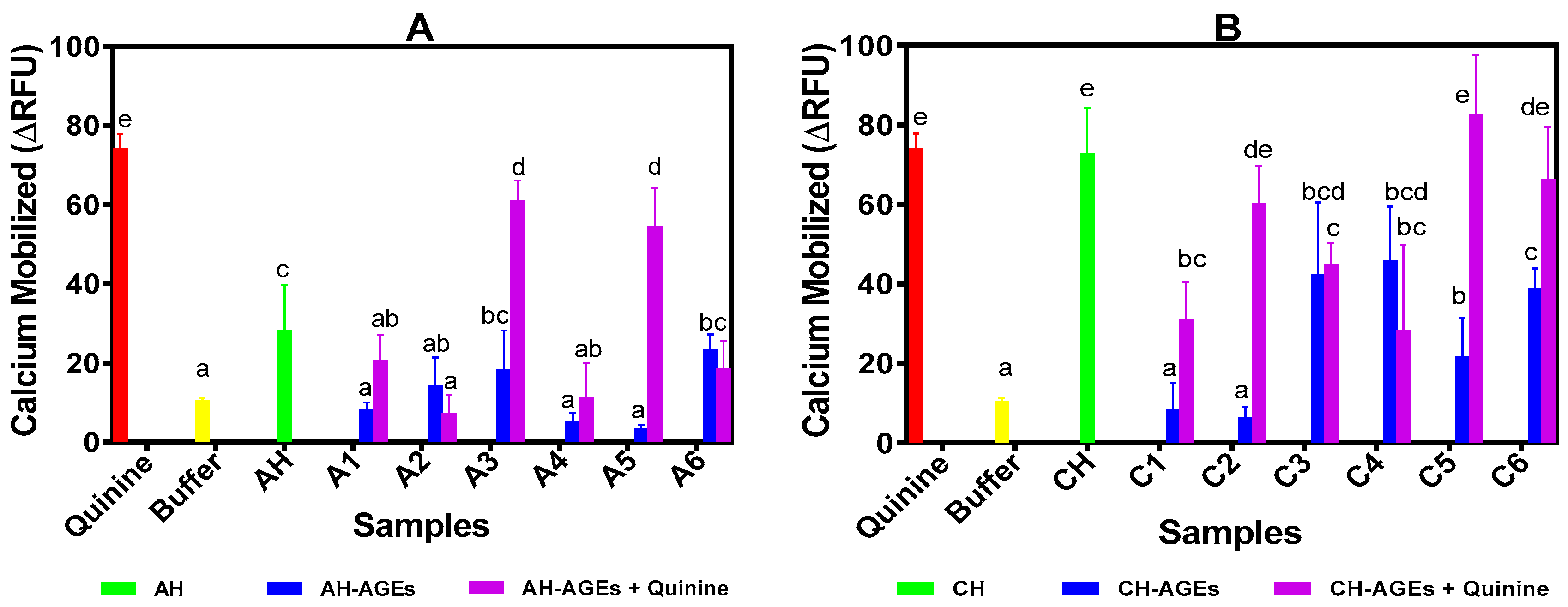
2.5. Determination of Calcium Mobilization
2.6. Statistical Analysis
3. Results
3.1. Degree of Glycation
3.2. Prediction of Bitter Score from Electronic Tongue
3.3. Determination of Inhibitory Ability Against T2R4 Activated by Quinine
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Greene, T.A.; Alarcon, S.; Thomas, A.; Berdougo, E.; Doranz, B.J.; Breslin, P.A.S.; Rucker, J.B. Probenecid inhibits the human bitter taste receptor TAS2R16 and suppresses bitter perception of salicin. PLoS ONE 2011, 6, e20123. [Google Scholar] [CrossRef] [PubMed]
- Pydi, S.P.; Sobotkiewicz, T.; Billakanti, R.; Bhullar, R.P.; Loewen, M.C.; Chelikani, P. Amino acid derivatives as bitter taste receptor (T2R) blockers. J. Biol. Chem. 2014, 289, 25054–25066. [Google Scholar] [CrossRef] [PubMed]
- Roland, W.S.U.; Gouka, R.J.; Gruppen, H.; Driesse, M.; Van Buren, L.; Smit, G.; Vincken, J.P. 6-Methoxyflavanones as bitter taste receptor blockers for hTAS2R39. PLoS ONE 2014, 9, e94451. [Google Scholar] [CrossRef] [PubMed]
- Jaggupilli, A.; Howard, R.; Upadhyaya, J.D.; Bhullar, R.P.; Chelikani, P. Bitter taste receptors: Novel insights into the biochemistry and pharmacology. Int. J. Biochem. Cell Biol. 2016, 77, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Ley, J.P. Masking bitter taste by molecules. Chemosens. Percept. 2008, 1, 58–77. [Google Scholar] [CrossRef]
- Slack, J.P.; Brockhoff, A.; Batram, C.; Menzel, S.; Sonnabend, C.; Born, S.; Meyerhof, W. Modulation of bitter taste perception by a small molecule hTAS2R antagonist. Curr. Biol. 2010, 20, 1104–1109. [Google Scholar] [CrossRef] [PubMed]
- Jaggupilli, A.; Howard, R.; Aluko, R.E.; Chelikani, P. Advanced glycation end-products can activate or block bitter taste receptors. Nutrients 2019, 11, 1317. [Google Scholar] [CrossRef]
- Wu, C.-H.; Huang, S.-M.; Lin, J.A.; Yen, G.-C. Inhibition of advanced glycation endproduct formation by foodstuffs. Food Funct. 2011, 2, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.U.E.; Pyzik, R.; Yong, A.; Striker, G.E. Advanced glycation end products in foods and a practical guide to their reduction in the diet. J. Am. Diet. Assoc. 2013, 110, 911–916. [Google Scholar]
- Uribarri, J.; Dolores, M.; Pía, M.; Maza, D.; Filip, R.; Gugliucci, A.; Wrobel, K. Dietary advanced glycation end products and their role in health and disease. Adv. Nutr. 2015, 6, 461–473. [Google Scholar] [CrossRef]
- Liu, P.; Huang, M.; Song, S.; Hayat, K.; Zhang, X.; Xia, S.; Jia, C. Sensory characteristics and antioxidant activities of Maillard reaction products from soy protein hydrolysates with different molecular weight distribution. Food Bioprocess Tech. 2012, 5, 1775–1789. [Google Scholar] [CrossRef]
- Oh, N.S.; Lee, H.A.; Lee, J.Y.; Joung, J.Y.; Lee, K.B.; Kim, Y.; Kim, S.H. The dual effects of Maillard reaction and enzymatic hydrolysis on the antioxidant activity of milk proteins. J. Dairy Sci. 2013, 96, 4899–4911. [Google Scholar] [CrossRef] [PubMed]
- Namiki, T.; Nakamura, T. Enhancement of Sugar Sweetness by Furanones and/or Cyclotene. Japanese Patent JP 04008264, 1992. [Google Scholar]
- Ottinger, H.; Soldo, T.; Hofmann, T. Discovery and structure determination of a novel Maillard-derived sweetness enhancer by application of the comparative taste dilution analysis (cTDA). J. Agric. Food Chem. 2003, 51, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Wei, B.; Chen, B.; McClements, D.J.; Decker, E.A. Chemical and antioxidant properties of casein peptide and its glucose Maillard reaction products in fish oil-in-water emulsions. J. Agric. Food Chem. 2011, 59, 13311–13317. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.G. Nutritional composition of red meat. Nutr. Diet. 2007, 64 (Suppl. 4), S113–S119. [Google Scholar] [CrossRef] [Green Version]
- Guerard, F.; Dufosse, L.; De La Broise, D.; Binet, A. Enzymatic hydrolysis of proteins from yellowfin tuna (Thunnus albacares) wastes using Alcalase. J. Mol. Catal. B 2001, 11, 1051–1059. [Google Scholar] [CrossRef]
- Feng, L.; Qiao, Y.; Zou, Y.; Huang, M.; Kang, Z.; Zhou, G. Effect of Flavourzyme on proteolysis, antioxidant capacity and sensory attributes of Chinese sausage. Meat Sci. 2014, 98, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Meinlschmidt, P.; Sussmann, D.; Schweiggert-Weisz, U.; Eisner, P. Enzymatic treatment of soy protein isolates: Effects on the potential allergenicity, technofunctionality, and sensory properties. Food Sci. Nutr. 2016, 4, 11–23. [Google Scholar] [CrossRef]
- O’Meara, G.M.; Munro, P.A. Hydrolysis of the sarcoplasmic, myofibrillar and connective tissue proteins of lean beef by alcalase and its relationship to whole meat hydrolysis. Meat Sci. 1985, 12, 91–103. [Google Scholar] [CrossRef]
- Zhang, C.; Alashi, A.M.; Singh, N.; Liu, K.; Chelikani, P.; Aluko, R.E. Beef protein-derived peptides as bitter taste receptor T2R4 blockers. J. Agric. Food Chem. 2018, 66, 4902–4912. [Google Scholar] [CrossRef]
- Charoenphun, N.; Cheirsilp, B.; Sirinupong, N.; Youravong, W. Calcium-binding peptides derived from tilapia (Oreochromis niloticus) protein hydrolysate. Eur. Food Res. Technol. 2013, 236, 57–63. [Google Scholar] [CrossRef]
- Nielsen, P.M.; Petersen, D.; Dambmann, C. Improved method for determining food protein degree of hydrolysis. J. Food Sci. 2001, 66, 642–646. [Google Scholar] [CrossRef]
- Alpha MOS. Astree Electrochemical Sensor Technology—Technical Note: T-SAS-04; Alpha MOS: Toulouse, France, 2004. [Google Scholar]
- Li, Y.; Zhong, F.; Ji, W.; Yokoyama, W.; Shoemaker, C.F.; Zhu, S.; Xia, W. Functional properties of Maillard reaction products of rice protein hydrolysates with mono-, oligo- and polysaccharides. Food Hydrocoll. 2013, 30, 53–60. [Google Scholar] [CrossRef]
- Song, N.; Tan, C.; Huang, M.; Liu, P.; Eric, K.; Zhang, X.; Jia, C. Transglutaminase cross-linking effect on sensory characteristics and antioxidant activities of Maillard reaction products from soybean protein hydrolysates. Food Chem. 2013, 136, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Amezquita, L.E.; Martinez-Alvarenga, M.S.; Olivas, G.I.; Zamudio-Flores, P.B.; Acosta-Muñiz, C.H.; Sepulveda, D.R. Effect of Maillard reaction conditions on the degree of glycation and functional properties of whey protein isolate-Maltodextrin conjugates. Food Hydrocoll. 2014, 38, 110–118. [Google Scholar]
- Ogasawara, M.; Katsumata, T.; Egi, M. Taste properties of Maillard-reaction products prepared from 1000 to 5000 Da peptide. Food Chem. 2006, 99, 600–604. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, biochemistry, and safety of acrylamide. A review. J. Agric. Food Chem. 2003, 51, 4504–4526. [Google Scholar] [CrossRef]
- Vattem, D.A.; Shetty, K. Acrylamide in food: A model for mechanism of formation and its reduction. Innov. Food Sci. Emerg. Technol. 2003, 4, 331–338. [Google Scholar] [CrossRef]
- Li, W.; Zhao, H.; He, Z.; Zeng, M.; Qin, F.; Chen, J. Modification of soy protein hydrolysates by Maillard reaction: Effects of carbohydrate chain length on structural and interfacial properties. Colloids Surf. B 2016, 138, 70–77. [Google Scholar] [CrossRef]
- Newman, J.; Harbourne, N.; O’Riordan, D.; Jacquier, J.C.; O’Sullivan, M. Comparison of a trained sensory panel and an electronic tongue in the assessment of bitter dairy protein hydrolysates. J. Food Eng. 2014, 128, 127–131. [Google Scholar] [CrossRef]
- Akitomi, H.; Tahara, Y.; Yasuura, M.; Kobayashi, Y.; Ikezaki, H.; Toko, K. Quantification of tastes of amino acids using taste sensors. Sens. Actuators B 2013, 179, 276–281. [Google Scholar] [CrossRef]
- Choi, D.H.; Kim, N.A.; Nam, T.S.; Lee, S.; Jeong, S.H. Evaluation of taste-masking effects of pharmaceutical sweeteners with an electronic tongue system. Drug Dev. Ind. Pharm. 2014, 40, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Miyanaga, Y.; Tanigake, A.; Nakamura, T.; Kobayashi, Y.; Ikezaki, H.; Taniguchi, A.; Uchida, T. Prediction of the bitterness of single, binary- and multiple-component amino acid solutions using a taste sensor. Int. J. Pharm. 2002, 248, 207–218. [Google Scholar] [CrossRef]
- Rachid, O.; Simons, F.E.R.; Rawas-Qalaji, M.; Simons, K.J. An electronic tongue: Evaluation of the masking efficacy of sweetening and/or flavoring agents on the bitter taste of epinephrine. AAPS PharmSciTech. 2010, 11, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Onitake, H.; Haraguchi, T.; Tahara, Y.; Yatabe, R.; Yoshida, M.; Toko, K. Quantitative prediction of bitterness masking effect of high-potency sweeteners using taste sensor. Sens. Actuators B 2016, 235, 11–17. [Google Scholar] [CrossRef]
- Behrens, M.; Meyerhof, W. Bitter taste receptors and human bitter taste perception. Cell. Mol. Life Sci. 2006, 63, 1501–1509. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Compounds | Used to Build Bitterness Standard Model | Used to Validate Bitterness Standard Model | Concentration (mM) | Published Values |
|---|---|---|---|---|
| Caffeine | √ | 0.24 2.36 | 2.5 8.5 | |
| Quinine | √ | 0.03 0.12 | 9 15.5 | |
| Prednisolone | √ | 0.44 0.88 | 13.5 17 | |
| Paracetamol | √ | 3.31 19.85 | 4 11 | |
| Loperamide | √ | 0.002 0.01 | 7.5 14 | |
| Famotidine | √ | 0.06 0.15 | 4.2 9 |
| Hydrolysate Concentration (%) | Glucose Concentration (M) | DG (%) * | |
|---|---|---|---|
| Alcalase AGEs | Chymotrypsin AGEs | ||
| 1 | 0.1 | 46.67 a | 29.53 a |
| 1 | 0.2 | 47.53 b | 29.11 a |
| 1 | 0.3 | 54.15 e | 30.13 b |
| 1 | 0.4 | 54.57 f | 36.67 d |
| 1 | 0.5 | 56.45 g | 37.14 e |
| 3 | 0.1 | 48.00 c | 29.49 a |
| 3 | 0.2 | 48.29 c | 28.86 a |
| 3 | 0.3 | 48.68 d | 36.63 cd |
| 3 | 0.4 | 54.88 f | 36.40 c |
| 3 | 0.5 | 54.96 fg | 38.60 f |
| Intensity | Range |
|---|---|
| Taste not detected | 1–4.5 |
| Slight taste | 4.5–8.5 |
| Acceptable | 8.5–12.5 |
| Acceptable limit | 12.5–16.5 |
| Not acceptable | 16.5–20.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Alashi, A.M.; Singh, N.; Chelikani, P.; Aluko, R.E. Glycated Beef Protein Hydrolysates as Sources of Bitter Taste Modifiers. Nutrients 2019, 11, 2166. https://doi.org/10.3390/nu11092166
Zhang C, Alashi AM, Singh N, Chelikani P, Aluko RE. Glycated Beef Protein Hydrolysates as Sources of Bitter Taste Modifiers. Nutrients. 2019; 11(9):2166. https://doi.org/10.3390/nu11092166
Chicago/Turabian StyleZhang, Chunlei, Adeola M. Alashi, Nisha Singh, Prashen Chelikani, and Rotimi E. Aluko. 2019. "Glycated Beef Protein Hydrolysates as Sources of Bitter Taste Modifiers" Nutrients 11, no. 9: 2166. https://doi.org/10.3390/nu11092166
APA StyleZhang, C., Alashi, A. M., Singh, N., Chelikani, P., & Aluko, R. E. (2019). Glycated Beef Protein Hydrolysates as Sources of Bitter Taste Modifiers. Nutrients, 11(9), 2166. https://doi.org/10.3390/nu11092166