Plasma Oxidative Status in Preterm Infants Receiving LCPUFA Supplementation: A Pilot Study

 ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Supplementation
2.3. Clinical and Anthropometrical Parameters
2.4. Blood Sample Extraction and Plasma Collection
2.5. Plasma Parameters of Oxidative Status
2.6. Plasma LCPUFAs
2.7. Statistical Analysis
3. Results
3.1. Neonatal Cohort and Baseline Characteristics
3.2. Diet Supplementation and Plasma LCPUFA Profiles
3.3. Diet Supplementation and Plasma Oxidative Status
3.4. Effect of Diet Supplementation on Neonatal Growth
4. Discussion
4.1. Effect of Supplementation on Plasma LCPUFAs
4.2. Effect of Supplementation on Oxidative Status
4.3. Effect of LCPUFA on Infant Growth
4.4. Influence of Sex
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, J.W.; Davis, J.M. Future applications of antioxidants in premature infants. Curr. Opin. Pediatr. 2011, 23, 161–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrone, S.; Tataranno, M.L.; Negro, S.; Cornacchione, S.; Longini, M.; Proietti, F.; Soubasi, V.; Benders, M.J.; VanBel, F.; Buonocore, G. May oxidative stress biomarkers in cord blood predict the occurrence of necrotizing enterocolitis in preterm infants? J. Matern. Fetal. Neonatal. Med. 2012, 25 (Suppl. 1), 128–131. [Google Scholar] [CrossRef]
- Shim, S.Y.; Kim, H.S. Oxidative stress and the antioxidant enzyme system in the developing brain. Korean J. Pediatr. 2013, 56, 107–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Cuevas, I.; Parra-Llorca, A.; Sánchez-Illana, A.; Nuñez-Ramiro, A.; Kuligowski, J.; Cháfer-Pericás, C.; Cernada, M.; Escobar, J.; Vento, M. Oxygen and oxidative stress in the perinatal period. Redox Biol. 2017, 12, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Perrone, S.; Bracciali, C.; Di Virgilio, N.; Buonocore, G. Oxygen Use in Neonatal Care: A Two-edged Sword. Front. Pediatr. 2017, 4, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuipers, R.S.; Luxwolda, M.F.; Offringa, P.J.; Boersma, E.R.; Dijck-Brouwer, D.A.; Muskiet, F.A. Fetal intrauterine whole body linoleic, arachidonic and docosahexaenoic acid contents and accretion rates. Prostaglandins Leukot. Essent. Fat. Acids 2012, 86, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.T.; Martin, C.R. Fatty acid requirements for the preterm infant. Semin. Fetal Neonatal Med. 2017, 22, 8–14. [Google Scholar] [CrossRef]
- Martin, C.R.; Dasilva, D.A.; Cluette-Brown, J.E.; Dimonda, C.; Hamill, A.; Bhutta, A.Q.; Coronel, E.; Wilschanski, M.; Stephens, A.J.; Driscoll, D.F.; et al. Decreased postnatal docosahexaenoic and arachidonic acid blood levels in premature infants are associated with neonatal morbidities. J. Pediatr. 2011, 159, 743–749. [Google Scholar] [CrossRef] [Green Version]
- Carlson, S.E.; Cooke, R.J.; Werkman, S.H.; Tolley, E.A. First year growth of preterm infants fed standard compared to marine oil n-3 supplemented formula. Lipids 1992, 27, 901–907. [Google Scholar] [CrossRef]
- Robinson, D.T.; Palac, H.L.; Baillif, V.; Van Goethem, E.; Dubourdeau, M.; Van Horn, L.; Martin, C.R. Long chain fatty acids and related pro-inflammatory, specialized pro-resolving lipid mediators and their intermediates in preterm human milk during the first month of lactation. Prostaglandins Leukot. Essent. Fat. Acids 2017, 121, 1–6. [Google Scholar] [CrossRef]
- Biagetti, C.; Correani, A.; D’Ascenzo, R.; Bellagamba, M.P.; Marchionni, P.; Antognoli, L.; Pompilio, A.; Cogo, P.; Carnielli, V.P. Does intravenous fish oil affect the growth of extremely low birth weight preterm infants on parenteral nutrition? Clin. Nutr. 2019, 38, 2319–2324. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B.; Bergmann, K.; Brenna, J.T.; Calder, P.C.; Campoy, C.; Clandinin, M.T.; Colombo, J.; Daly, M.; Decsi, T.; Demmelmair, H.; et al. Should formula for infants provide arachidonic acid along with DHA? A position paper of the European Academy of Paediatrics and the Child Health Foundation. Am. J. Clin. Nutr. 2019. [Google Scholar] [CrossRef] [PubMed]
- Agostoni, C.; Buonocore, G.; Carnielli, V.P.; De Curtis, M.; Darmaun, D.; Decsi, T.; Domellöf, M.; Embleton, N.D.; Fusch, C.; Genzel-Boroviczeny, O.; et al. Enteral nutrient supply for preterm infants: Commentary from the European Society of Paediatric Gastroenterology, Hepatology and Nutrition Committee on Nutrition. J. Pediatr. Gastroenterol. Nutr. 2010, 50, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A. Effect of fish and fish oil-derived omega-3 fatty acids on lipid oxidation. Redox Rep. 2004, 9, 193–197. [Google Scholar] [CrossRef]
- Richard, D.; Kefi, K.; Barbe, U.; Bausero, P.; Visioli, F. Polyunsaturated fatty acids as antioxidants. Pharmacol. Res. 2008, 57, 451–455. [Google Scholar] [CrossRef]
- Malamas, A.; Chranioti, A.; Tsakalidis, C.; Dimitrakos, S.A.; Mataftsi, A. The omega-3 and retinopathy of prematurity relationship. Int. J. Ophthalmol. 2017, 10, 300–305. [Google Scholar] [CrossRef]
- Kajarabille, N.; Hurtado, J.A.; Peña-Quintana, L.; Peña, M.; Ruiz, J.; Diaz-Castro, J.; Rodríguez-Santana, Y.; Martin-Alvarez, E.; López-Frias, M.; Soldado, O.; et al. Omega-3 LCPUFA supplement: A nutritional strategy to prevent maternal and neonatal oxidative stress. Matern. Child Nutr. 2017, 13, e12300. [Google Scholar] [CrossRef]
- Eritsland, J. Safety considerations of polyunsaturated fatty acids. Am. J. Clin. Nutr. 2000, 71 (Suppl. 1), 197S–201S. [Google Scholar] [CrossRef]
- Shoji, H.; Franke, C.; Campoy, C.; Rivero, M.; Demmelmair, H.; Koletzko, B. Effect of docosahexaenoic acid and eicosapentaenoic acid supplementation on oxidative stress levels during pregnancy. Free. Radic. Res. 2006, 40, 379–384. [Google Scholar] [CrossRef]
- Saghaei, M. Random allocation software for parallel group randomized trials. BMC Med. Res. Methodol. 2004, 4, 26. [Google Scholar] [CrossRef] [Green Version]
- Lapillonne, A.; Jensen, C.L. Reevaluation of the DHA requirement for the premature infant. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Muktan, D.; Singh, R.R.; Bhatta, N.K.; Shah, D. Neonatal mortality risk assessment using SNAPPE- II score in a neonatal intensive care unit. BMC Pediatr. 2019, 19, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenton, T.R.; Kim, J.H. A systematic review and meta-analysis to revise the Fenton growth chart for preterm infants. BMC Pediatr. 2013, 13, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, A.L.; Engstrom, J.L.; Meier, P.P.; Jegier, B.J.; Kimura, R.E. Calculating postnatal growth velocity in very low birth weight (VLBW) premature infants. J. Perinatol. 2009, 29, 618–622. [Google Scholar] [CrossRef] [Green Version]
- Fenton, T.R.; Anderson, D.; Groh-Wargo, S.; Hoyos, A.; Ehrenkranz, R.A.; Senterre, T. An Attempt to Standardize the Calculation of Growth Velocity of Preterm Infants-Evaluation of Practical Bedside Methods. J. Pediatr. 2018, 196, 77–83. [Google Scholar] [CrossRef]
- Condezo-Hoyos, L.; Rubio, M.; Arribas, S.M.; España-Caparrós, G.; Rodríguez-Rodríguez, P.; Mujica-Pacheco, E.; González, M.C. A plasma oxidative stress global index in early stages of chronic venous insufficiency. J. Vasc. Surg. 2013, 57, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Berkban, T.; Boonprom, P.; Bunbupha, S.; Welbat, J.U.; Kukongviriyapan, U.; Kukongviriyapan, V.; Pakdeechote, P.; Prachaney, P. Ellagic Acid Prevents L-NAME-Induced Hypertension via Restoration of eNOS and p47phox Expression in Rats. Nutrients 2015, 7, 5265–5280. [Google Scholar] [CrossRef]
- Jia, Z.; Saha, S.; Zhu, H.; Li, Y.; Misra, H. Spectrofluorometric measurement of reduced glutathione levels in human neuronal cells. In Methods Redox Signaling; Mary Ann Liebert: New York, NY, USA, 2010; pp. 235–237. [Google Scholar]
- Ramiro-Cortijo, D.; Herrera, T.; Rodríguez-Rodríguez, P.; López De Pablo, Á.L.; De La Calle, M.; López-Giménez, M.R.; Mora-Urda, A.I.; Gutiérrez-Arzapalo, P.Y.; Gómez-Rioja, R.; Aguilera, Y.; et al. Maternal plasma antioxidant status in the first trimester of pregnancy and development of obstetric complications. Placenta 2016, 47, 37–45. [Google Scholar] [CrossRef]
- Freedman, S.D.; Blanco, P.G.; Zaman, M.M.; Shea, J.C.; Ollero, M.; Hopper, I.K.; Weed, D.A.; Gelrud, A.; Regan, M.M.; Laposata, M.; et al. Association of cystic fibrosis with abnormalities in fatty acid metabolism. N. Engl. J. Med. 2004, 350, 560–569. [Google Scholar] [CrossRef]
- Brenna, J.T.; Varamini, B.; Jensen, R.G.; Diersen-Schade, D.A.; Boettcher, J.A.; Arterburn, L.M. Docosahexaenoic and arachidonic acid concentrations in human breast milk worldwide. Am. J. Clin. Nutr. 2007, 85, 1457–1464. [Google Scholar] [CrossRef] [Green Version]
- Alshweki, A.; Muñuzuri, A.P.; Baña, A.M.; De Castro, M.J.; Andrade, F.; Aldamiz-Echevarría, L.; Saenz de Pipaón, M.; Fraga, J.M.; Couce, M.L. Effects of different arachidonic acid supplementation on psychomotor development in very preterm infants; a randomized controlled trial. Nutr. J. 2015, 14, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattori, T.; Obinata, H.; Ogawa, A.; Kishi, M.; Tatei, K.; Ishikawa, O.; Izumi, T. G2A plays proinflammatory roles in human keratinocytes under oxidative stress as a receptor for 9-hydroxyoctadecadienoic acid. J. Investig. Dermatol. 2008, 128, 1123–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabbs, M.; Leng, S.; Devassy, J.G.; Monirujjaman, M.; Aukema, H.M. Advances in Our Understanding of Oxylipins Derived from Dietary PUFAs. Adv. Nutr. 2015, 6, 513–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Plopper, C.G.; Lakritz, J.; Storms, D.H.; Hammock, B.D. Leukotoxindiol: A putative toxic mediator involved in acute respiratory distress syndrome. Am. J. Respir. Cell Mol. Biol. 2001, 25, 434–438. [Google Scholar] [CrossRef]
- McMurchie, E.J.; Rinaldi, J.A.; Burnard, S.L.; Patten, G.S.; Neumann, M.; McIntosh, G.H.; Abbey, M.; Gibson, R.A. Incorporation and effects of dietary eicosapentaenoate (20:5(n-3)) on plasma and erythrocyte lipids of the marmoset following dietary supplementation with differing levels of linoleic acid. Biochim. Biophys. Acta 1990, 1045, 164–173. [Google Scholar] [CrossRef]
- Dias, C.B.; Wood, L.G.; Garg, M.L. Effects of dietary saturated and n-6 polyunsaturated fatty acids on the incorporation of long-chain n-3 polyunsaturated fatty acids into blood lipids. Eur. J. Clin. Nutr. 2016, 70, 812–818. [Google Scholar] [CrossRef]
- Gibson, R.A. Musings about the role dietary fats after 40 years of fatty acid research. Prostaglandins Leukot. Essent. Fat. Acids 2018, 131, 1–5. [Google Scholar] [CrossRef]
- Uauy, R.; Mena, P. Long-chain polyunsaturated fatty acids supplementation in preterm infants. Curr. Opin. Pediatr. 2015, 27, 165–171. [Google Scholar] [CrossRef]
- Ruiz-Hurtado, G.; Condezo-Hoyos, L.; Pulido-Olmo, H.; Aranguez, I.; Gónzalez, M.C.; Arribas, S.M.; Cerezo, C.; Segura, J.; Praga, M.; Fernández-Alfonso, M.S.; et al. Development of albuminuria and enhancement of oxidative stress during chronic renin-angiotensin system suppression. J. Hypertens. 2014, 32, 2082–2091. [Google Scholar] [CrossRef]
- Siahanidou, T.; Lazaropoulou, C.; Michalakakou, K.; Papassotiriou, I.; Bacoula, C.; Mandyla, H. Oxidative stress in preterm infants fed a formula containing long-chain polyunsaturated fatty acids (LCPUFA). Am. J. Perinatol. 2007, 24, 475–479. [Google Scholar] [CrossRef]
- Deshpande, G.; Simmer, K.; Deshmukh, M.; Mori, T.A.; Croft, K.D.; Kristensen, J. Fish Oil (SMOFlipid) and olive oil lipid (Clinoleic) in very preterm neonates. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 177–182. [Google Scholar] [CrossRef]
- Halliwell, B. The antioxidant paradox: Less paradoxical now? Br. J. Clin. Pharmacol. 2013, 75, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Pingitore, A.; Lima, G.P.; Mastorci, F.; Quinones, A.; Iervasi, G.; Vassalle, C. Exercise and oxidative stress: Potential effects of antioxidant dietary strategies in sports. Nutrition 2015, 31, 916–922. [Google Scholar] [CrossRef]
- Eriksson, J.G.; Kajantie, E.; Osmond, C.; Thornburg, K.; Barker, D.J. Boys live dangerously in the womb. Am. J. Hum. Biol. 2010, 22, 330–335. [Google Scholar] [CrossRef] [Green Version]
- Peelen, M.J.; Kazemier, B.M.; Ravelli, A.C.; De Groot, C.J.; Van Der Post, J.A.; Mol, B.W.; Hajenius, P.J.; Kok, M. Impact of fetal gender on the risk of preterm birth, a national cohort study. Acta Obstet. Gynecol. Scand. 2016, 95, 1034–1041. [Google Scholar] [CrossRef] [Green Version]
- Ramiro-Cortijo, D.; Zozaya-Nieto, C.; López de Pablo, A.L.; López-Giménez, M.R.; Saenz de Pipaon, M.; Arribas, S.M. Higher risk of late-onset sepsis in very low birth weight male preterm infants. Rev. Med. Univ. 2018, 20, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Zozaya, C.; Avila-Alvarez, A.; Arruza, L.; García-Muñoz, R.F.; Fernandez-Perez, C.; Castro, A.; Cuesta, M.T.; Vacas, B.; Couce, M.L.; Vento, M.; et al. The Effect of Morbidity and Sex on Postnatal Growth of Very Preterm Infants: A Multicenter Cohort Study. Neonatology 2019, 115, 348–354. [Google Scholar] [CrossRef]
- Lavoie, J.C.; Tremblay, A. Sex-Specificity of Oxidative Stress in Newborns Leading to a Personalized Antioxidant Nutritive Strategy. Antioxidants 2018, 7, 49. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Figure 12. | FormulaidTM | MCT NM |
|---|---|---|
| Lauric Acid (C12:0) | 1.3 | - |
| Myristic Acid (C14:0) | 3.1 | 0.1 |
| Palmitic Acid (C16:0) | 14.2 | 16.3 |
| Palmitoleic Acid (C16:1 n-7) | 0.5 | 0.2 |
| Stearic Acid (C18:0) | 4.5 | 2.7 |
| Oleic Acid (C18:1 n-9) | 20.8 | 20.4 |
| Linoleic Acid (C18:2 n-6) | 27.5 | 53.5 |
| Linolenic Acid (C18:3 n-6) | 1.0 | - |
| Alpha Linoleic Acid (C18:3 n-3) | 3.0 | 4.6 |
| Arachidic Acid (C20:0) | 0.4 | 0.3 |
| Gondoic Acid (C20:1 n-9) | 0.2 | 0.3 |
| Eicosadienoic Acid (C20:2 n-6) | 0.2 | - |
| Eicosatrienoic Acid (C20:3 n-6) | 0.9 | - |
| Arachidonic Acid (C20:4 n-6) | 13.5 | - |
| Eicosapentaenoic Acid (C20:5 n-3) | 0.1 | - |
| Behenic Acid (C22:0) | 0.5 | - |
| Docosahexaenoic Acid (C22:6 n-3) | 6.3 | - |
| Lignoceric Acid (C24:0) | 0.4 | - |
| MCT-S (n = 12) | ARA:DHA-S (n =1 0) | p-Value | |
|---|---|---|---|
| Maternal age (years) | 34.5 (9.0) | 31.0 (3.0) | 0.447 |
| Gestational age (weeks) | 28.9 (2.6) | 27.9 (1.4) | 0.382 |
| Apgar at 1 min | 7.0 (1.3) | 7.0 (1.0) | 0.602 |
| Apgar at 5 min | 8.0 (1.0) | 8.0 (2.0) | 0.549 |
| Score Neonatal Acute Physiology-II | 24.0 (15.3) | 23.0 (5.0) | 0.862 |
| Sex (female) | 50.0% (6) | 85.7% (7) | 0.112 |
| Birth weight (g) | 856.0 (314.5) | 1075.0 (381.0) | 0.241 |
| Birth weight z-score | −0.12 (1.5) | 0.4 (0.8) | 0.102 |
| Intrauterine growth restriction | 16.7% (2) | 0% (0) | 0.198 |
| Birth length (cm) | 34.0 (4.4) | 35.5 (4.3) | 0.844 |
| Birth length z-score | −0.63 (2.2) | 0.45 (1.0) * | 0.020 |
| Birth head circumference (cm) | 25.0 (4.3) | 25.5 (3.2) | 0.262 |
| Birth head circumference z-score | −0.31 (1.2) | 0.98 (1.9) | 0.080 |
| MCT-S (n = 10) | ARA:DHA-S (n = 12) | p-Value | |
|---|---|---|---|
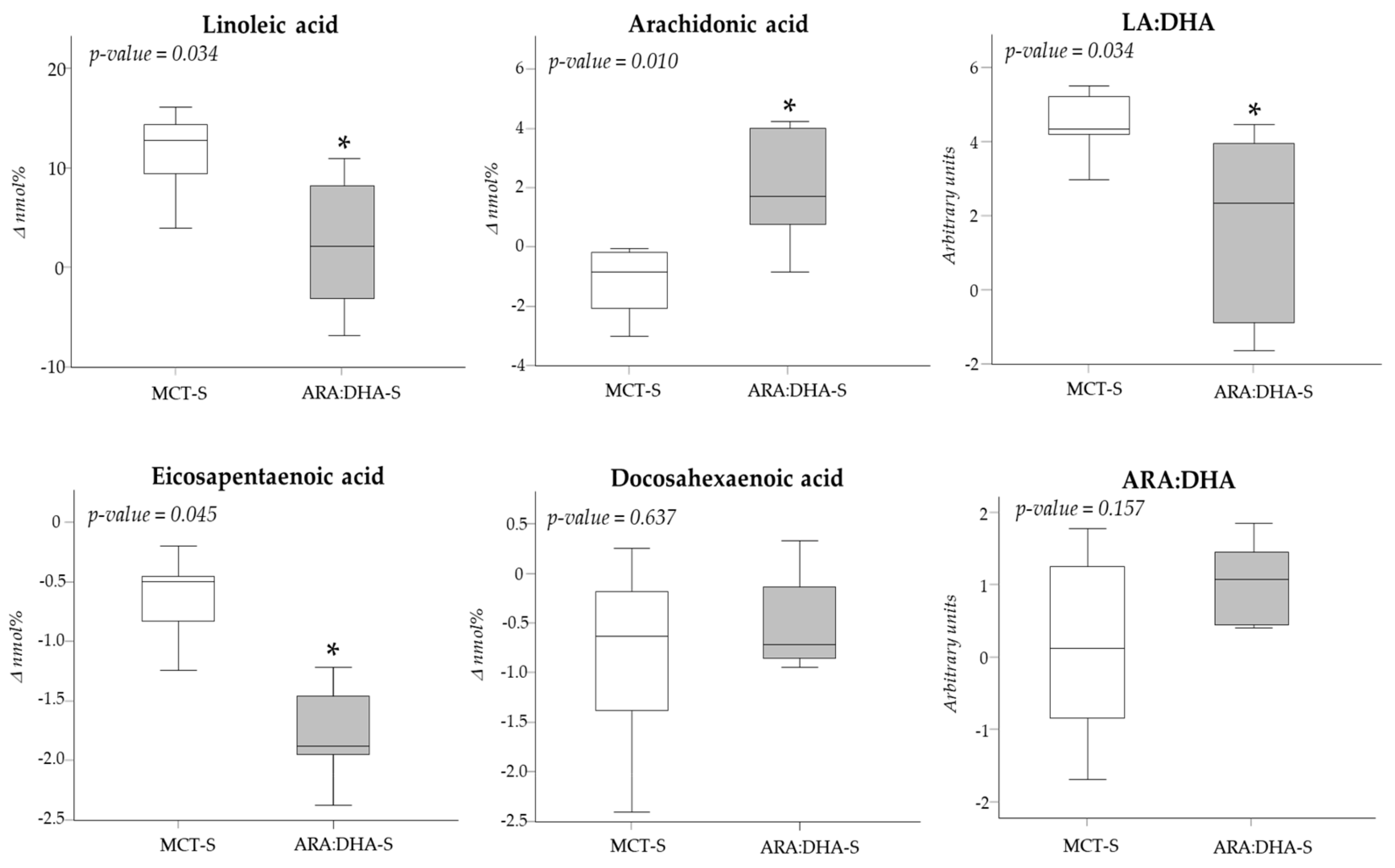
| Linoleic acid (nmol %) | |||
| At birth | 11.10 (5.0) | 15.52 (5.36) | 0.063 |
| At 28 DPL | 19.38 (0.57) | 15.74 (3.78) * | 0.005 |
| At 36 WPA | 22.80 (1.41) | 18.54 (1.68) * | 0.018 |
| Dihomo-γ-linolenic acid (nmol %) | |||
| At birth | 1.60 (0.41) | 1.40 (0.69) | 0.518 |
| At 28 DPL | 2.34 (0.48) | 1.96 (0.37) * | 0.040 |
| At 36 WPA | 2.32 (0.58) | 1.68 (0.38) * | 0.018 |
| Arachidonic acid (nmol %) | |||
| At birth | 8.31 (1.54) | 7.45 (3.45) | 0.470 |
| At 28 DPL | 7.91 (1.48) | 8.29 (2.47) | 0.329 |
| At 36 WPA | 7.50 (1.27) | 9.27 (2.34) | 0.050 |
| α-Linolenic acid (nmol %) | |||
| At birth | 0.23 (0.19) | 0.78 (0.49) | 0.121 |
| At 28 DPL | 0.16 (0.09) | 0.50 (0.35) * | 0.006 |
| At 36 WPA | 0.22 (0.10) | 0.52 (0.33) * | 0.018 |
| Eicosapentaenoic acid (nmol %) | |||
| At birth | 0.78 (0.65) | 1.63 (0.57) * | 0.037 |
| At 28 DPL | 0.55 (0.80) | 0.62 (0.90) | 0.491 |
| At 36 WPA | 0.28 (0.31) | 0.44 (0.24) | 0.289 |
| Docosahexaenoic acid (nmol %) | |||
| At birth | 3.07 (1.40) | 3.08 (0.91) | 0.909 |
| At 28 DPL | 2.79 (0.62) | 3.08 (0.58) | 0.770 |
| At 36 WPA | 2.61 (1.17) | 2.83 (0.44) | 0.346 |
| Saturated fatty acids (nmol %) | |||
| At birth | 32.41 (3.29) | 31.97 (1.95) | 0.970 |
| At 28 DPL | 32.89 (2.47) | 35.40 (0.66) | 0.064 |
| At 36 WPA | 33.71 (1.91) | 35.70 (2.07) | 0.077 |
| Monounsaturated fatty acids (nmol %) | |||
| At birth | 38.72 (3.15) | 35.85 (3.33) * | 0.007 |
| At 28 DPL | 30.55 (2.93) | 30.75 (3.70) | 0.380 |
| At 36 WPA | 29.21 (2.46) | 30.26 (3.40) | 0.480 |
| n-6:n-3 LCPUFAs (arbitrary units) | |||
| At birth | 3.88 (1.46) | 3.91 (1.10) | 0.970 |
| At 28 DPL | 6.75 (2.45) | 5.16 (1.38) | 0.097 |
| At 36 WPA | 8.17 (1.78) | 7.48 (1.62) | 0.389 |
| Linoleic acid:docosahexaenoic acid (arbitrary units) | |||
| At birth | 4.01 (0.69) | 5.04 (1.17) * | 0.020 |
| At 28 DPL | 6.57 (1.06) | 4.67 (0.84) * | 0.006 |
| At 36 WPA | 8.21 (2.65) | 6.26 (2.35) * | 0.045 |
| Arachidonic acid:docosahexaenoic acid (arbitrary units) | |||
| At birth | 2.65 (1.56) | 2.52 (1.35) | 0.790 |
| At 28 DPL | 2.74 (1.15) | 2.59 (0.84) | 0.558 |
| At 36 WPA | 2.95 (0.77) | 3.41 (0.61) | 0.346 |
| MCT-S (n = 10) | ARA:DHA-S (n = 12) | p-Value | |
|---|---|---|---|
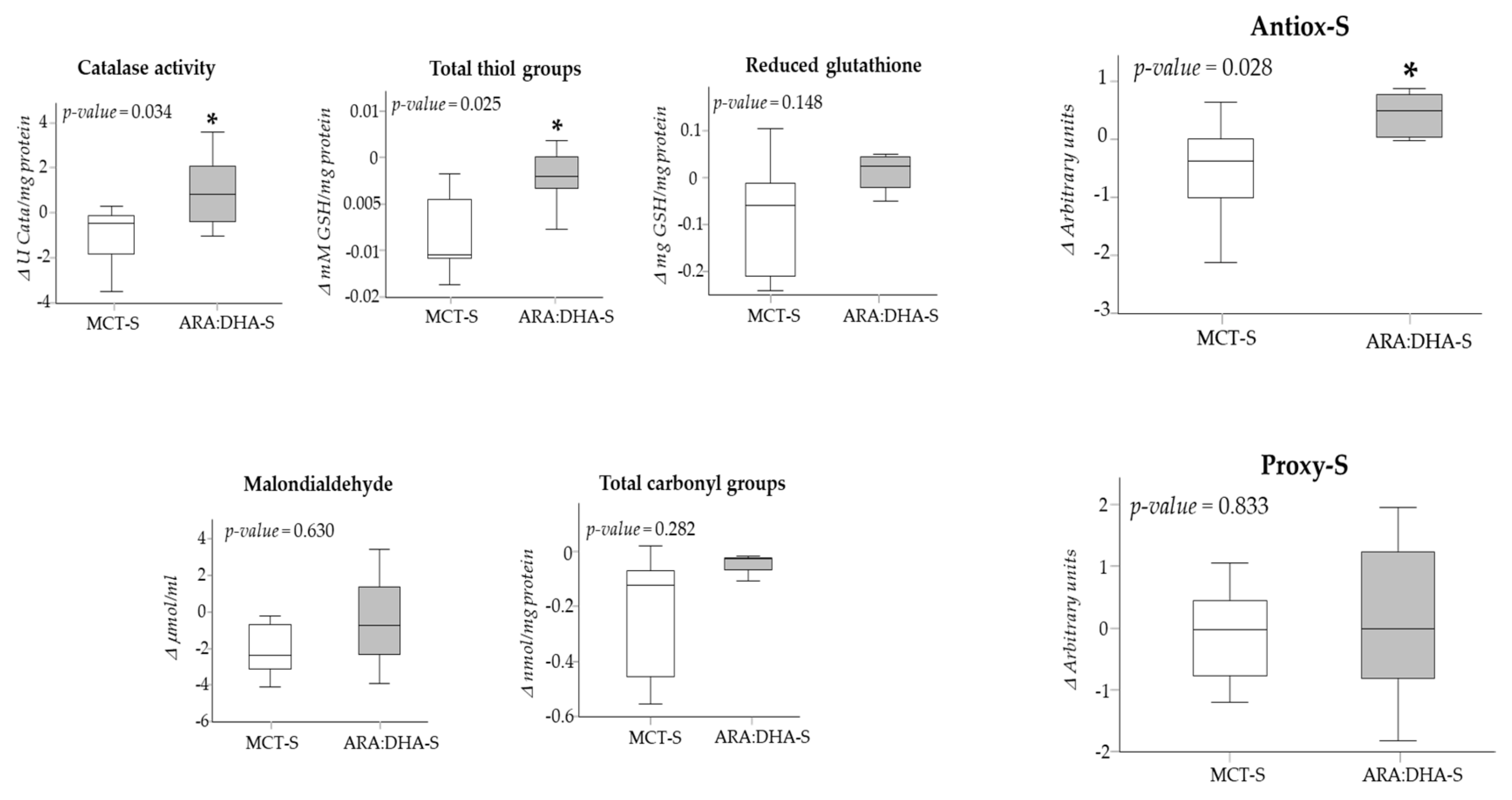
| Catalase activity (U catalase/mg protein) | |||
| At birth | 3.37 (3.16) | 3.86 (1.45) | 0.644 |
| At 28 DPL | 3.04 (1.63) | 4.61 (0.89) | 0.085 |
| At 36 WPA | 2.86 (0.94) | 4.74 (0.40) * | 0.025 |
| Total thiol groups (mM GSH/mg protein) | |||
| At birth | 0.013 (0.011) | 0.011 (0.002) | 0.337 |
| At 28 DPL | 0.007 (0.002) | 0.010 (0.005) | 0.134 |
| At 36 WPA | 0.007 (0.002) | 0.009 (0.003) | 0.113 |
| Reduced glutathione (mg GSH/mg protein) | |||
| At birth | 0.15 (0.22) | 0.20 (0.08) | 0.915 |
| At 28 DPL | 0.09 (0.26) | 0.25 (0.12) | 0.135 |
| At 36 WPA | 0.07 (0.12) | 0.26 (0.19) * | 0.021 |
| Antiox-S (arbitrary units) | |||
| At birth | −0.10 (1.64) | 0.03 (0.52) | 0.702 |
| At 28 DPL | −0.63 (1.11) | 0.36 (0.35) | 0.069 |
| At 36 WPA | −0.67 (0.96) | 0.55 (0.35) * | 0.045 |
| Malondialdehyde (μmol/ml) | |||
| At birth | 4.36 (1.44) | 6.46 (2.27) | 0.118 |
| At 28 DPL | 2.79 (0.68) | 4.63 (1.29) * | 0.005 |
| At 36 WPA | 2.41 (0.90) | 4.81 (3.70) * | 0.008 |
| Total carbonyl groups (nmol/mg protein) | |||
| At birth | 0.29 (0.36) | 0.25 (0.20) | 0.670 |
| At 28 DPL | 0.25 (0.29) | 0.42 (0.40) | 0.342 |
| At 36 WPA | 0.12 (0.14) | 0.22 (0.27) | 0.556 |
| Proxy-S (arbitrary units) | |||
| At birth | −0.01 (1.24) | 0.38 (0.72) | 0.382 |
| At 28 DPL | −0.29 (0.43) | 0.39 (0.93) | 0.052 |
| At 36 WPA | −0.27 (0.64) | 0.27 (1.15) * | 0.010 |
| Antiox-S:Proxy-S (arbitrary units) | |||
| At birth | 1.19 (2.18) | 0.07 (1.22) | 0.148 |
| At 28 DPL | 0.93 (4.10) | 0.43 (0.62) | 0.995 |
| At 36 WPA | 0.80 (1.38) | 0.79 (8.37) | 0.456 |
| Male (n = 8) | Female (n = 13) | p-Value | |
|---|---|---|---|
| Maternal age (years) | 33.0 (6.0) | 31.0 (10.0) | 0.840 |
| Gestational age (weeks) | 27.0 (4.0) | 28.0 (2.0) | 0.916 |
| Apgar at 1 min | 7.0 (2.5) | 7.0 (2.0) | 0.972 |
| Apgar at 5 min | 8.5 (2.0) | 8.0 (2.0) | 0.778 |
| Score of Neonatal Acute Physiology-II | 23.5 (10.5) | 23.0 (6.0) | 0.743 |
| Weight (g) | 1192.5 (472.5) | 856 (200) | 0.030 |
| Weight z-score | 0.44 (0.5) | −0,17 (0.6) | 0.161 |
| Intrauterine growth restriction | 12.5% (1) | 7.7% (1) | 0.716 |
| Length (cm) | 38 (4.0) | 34 (1.5) | 0.030 |
| Length z-score | 0.40 (0.5) | −0.50 (1.3) | 0.196 |
| Head circumference (cm) | 26.9 (3.5) | 24.4 (3.5) | 0.053 |
| Head circumference z-score | 1.16 (1.4) | −0.58 (0.5) | 0.020 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramiro-Cortijo, D.; López de Pablo, Á.L.; López-Giménez, M.R.; Martin, C.R.; Brown, J.; Saenz de Pipaón, M.; Arribas, S.M. Plasma Oxidative Status in Preterm Infants Receiving LCPUFA Supplementation: A Pilot Study. Nutrients 2020, 12, 122. https://doi.org/10.3390/nu12010122
Ramiro-Cortijo D, López de Pablo ÁL, López-Giménez MR, Martin CR, Brown J, Saenz de Pipaón M, Arribas SM. Plasma Oxidative Status in Preterm Infants Receiving LCPUFA Supplementation: A Pilot Study. Nutrients. 2020; 12(1):122. https://doi.org/10.3390/nu12010122
Chicago/Turabian StyleRamiro-Cortijo, David, Ángel Luis López de Pablo, Mᵃ Rosario López-Giménez, Camilia R. Martin, Joanne Brown, Miguel Saenz de Pipaón, and Silvia M. Arribas. 2020. "Plasma Oxidative Status in Preterm Infants Receiving LCPUFA Supplementation: A Pilot Study" Nutrients 12, no. 1: 122. https://doi.org/10.3390/nu12010122
APA StyleRamiro-Cortijo, D., López de Pablo, Á. L., López-Giménez, M. R., Martin, C. R., Brown, J., Saenz de Pipaón, M., & Arribas, S. M. (2020). Plasma Oxidative Status in Preterm Infants Receiving LCPUFA Supplementation: A Pilot Study. Nutrients, 12(1), 122. https://doi.org/10.3390/nu12010122