Phosvitin Derived Phospho-Peptides Show Better Osteogenic Potential than Intact Phosvitin in MC3T3-E1 Osteoblastic Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
2.3. Preparation of Phosvitin and PPP
2.4. Western Blotting
2.5. ELISA
2.6. Immunofluorescence
2.7. BrDU Incorporation Assay
2.8. Statistics
3. Results
3.1. Phosvitin Induces Cellular Markers of Osteoblast Differentiation
3.2. PPP also Induces Cellular Markers of Osteoblast Differentiation
3.3. Both Phosvitin and PPP Attenuate RANKL Release from Osteoblasts
3.4. PPP, But Not Phosvitin Alone, Increases Type I Collagen in Osteoblasts
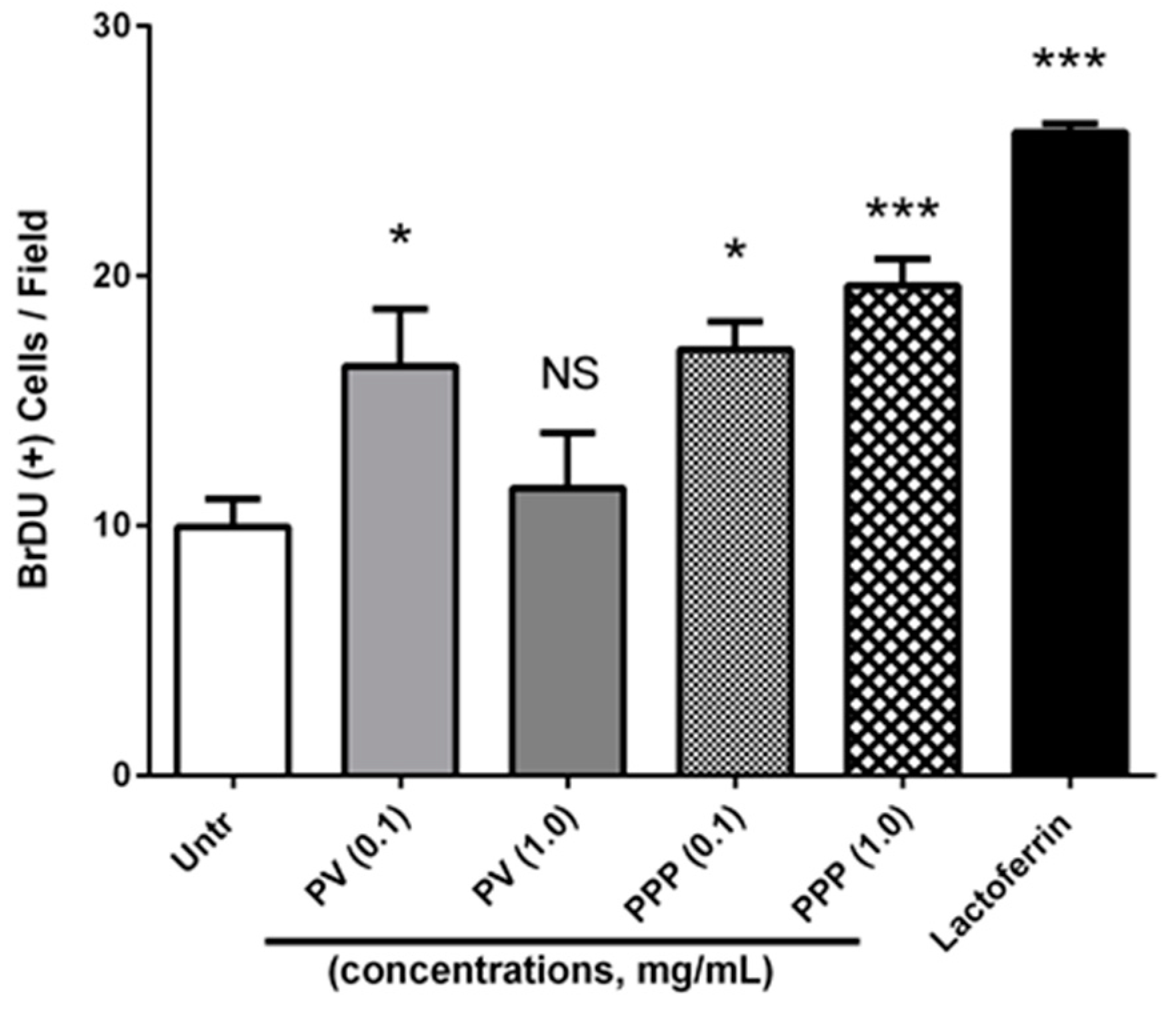
3.5. Both Phosvitin and PPP Promote Osteoblast Proliferation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chakrabarti, S.; Jahandideh, F.; Wu, J. Food-derived bioactive peptides on inflammation and oxidative stress. Biomed. Res. Int. 2014, 2014, 608979. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, F.; Zhong, Y. Bioactive peptides. J. AOAC Int. 2008, 91, 914–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salih, E. Phosphoproteomics by mass spectrometry and classical protein chemistry approaches. Mass. Spectrom. Rev. 2005, 24, 828–846. [Google Scholar] [CrossRef]
- Samaraweera, H.; Zhang, W.G.; Lee, E.J.; Ahn, D.U. Egg yolk phosvitin and functional phosphopeptides—review. J. Food Sci. 2011, 76, R143–R150. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Lonnerdal, B. Bioactive peptides derived from human milk proteins--mechanisms of action. J. Nutr. Biochem. 2014, 25, 503–514. [Google Scholar] [CrossRef]
- Scholz-Ahrens, K.E.; Schrezenmeir, J. Effects of bioactive substances in milk on mineral and trace element metabolism with special reference to casein phosphopeptides. Br. J. Nutr. 2000, 84, S147–S153. [Google Scholar] [CrossRef] [Green Version]
- Bouhallab, S.; Bougle, D. Biopeptides of milk: Caseinophosphopeptides and mineral bioavailability. Reprod Nutr. Dev. 2004, 44, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Tsuchita, H.; Suzuki, T.; Kuwata, T. The effect of casein phosphopeptides on calcium absorption from calcium-fortified milk in growing rats. Br. J. Nutr. 2001, 85, 5–10. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Lee, J.; Jang, J.H.; Sakchaisri, K.; Hwang, J.; Cha-Molstad, H.J.; Kim, K.A.; Ryoo, I.J.; Lee, H.G.; Kim, S.O.; et al. Osteoporosis regulation by salubrinal through eIF2alpha mediated differentiation of osteoclast and osteoblast. Cell. Signal. 2013, 25, 552–560. [Google Scholar] [CrossRef] [Green Version]
- Jobke, B.; Milovanovic, P.; Amling, M.; Busse, B. Bisphosphonate-osteoclasts: Changes in osteoclast morphology and function induced by antiresorptive nitrogen-containing bisphosphonate treatment in osteoporosis patients. Bone 2014, 59, 37–43. [Google Scholar] [CrossRef]
- Wysowski, D.K.; Chang, J.T. Alendronate and risedronate: Reports of severe bone, joint, and muscle pain. Arch. Intern. Med. 2005, 165, 346–347. [Google Scholar] [PubMed]
- Wu, J.; Majumder, K. Will intake of eggs/egg products have benefits for hypertension management? Poult. Sci. 2008, 87, 125. [Google Scholar]
- Abeyrathne, E.; Lee, Y.; Ahn, D. Egg white proteins and their potential use in food processing or as nutraceutical and pharmaceutical agents-A review. Poult. Sci. 2013, 92, 3292–3299. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Chakrabarti, S.; Majumder, K.; Jiang, Y.; Davidge, S.T.; Wu, J. Egg-derived peptide IRW inhibits TNF-alpha-induced inflammatory response and oxidative stress in endothelial cells. J. Agric. Food Chem. 2010, 58, 10840–10846. [Google Scholar] [CrossRef] [PubMed]
- Majumder, K.; Chakrabarti, S.; Morton, J.S.; Panahi, S.; Kaufman, S.; Davidge, S.T.; Wu, J.P. Egg-derived ACE-inhibitory peptides IQW and LKP reduce blood pressure in spontaneously hypertensive rats. J. Funct. Foods 2015, 13, 50–60. [Google Scholar] [CrossRef]
- Miguel, M.; Aleixandre, A. Anti hypertensive peptides derived from egg proteins. J. Nutr. 2006, 136, 1457–1460. [Google Scholar] [CrossRef] [Green Version]
- Mine, Y. Egg proteins and peptides in human health--chemistry, bioactivity and production. Curr. Pharm. Des. 2007, 13, 875–884. [Google Scholar] [CrossRef]
- Grogan, J.; Shirazi, A.; Taborsky, G. Phosphorus nuclear magnetic resonance of diverse phosvitin species. Comp. Biochem. Physiol. B 1990, 96, 655–663. [Google Scholar] [CrossRef]
- Byrne, B.; Schip, A.; Vandeklundert, J.; Arnberg, A.; Gruber, M.; Ab, G. Amino-acid-sequence of phosvitin derived from the nucleotide-sequence of part of the chicken vitellogenin gene. Biochemistry 1984, 23, 4275–4279. [Google Scholar] [CrossRef]
- Ishikawa, S.I.; Tamaki, S.; Arihara, K.; Itoh, M. Egg yolk protein and egg yolk phosvitin inhibit calcium, magnesium, and iron absorptions in rats. J. Food Sci. 2007, 72, S412–S419. [Google Scholar] [CrossRef]
- Liu, J.; Czernick, D.; Lin, S.C.; Alasmari, A.; Serge, D.; Salih, E. Novel bioactivity of phosvitin in connective tissue and bone organogenesis revealed by live calvarial bone organ culture models. Dev. Biol. 2013, 381, 256–275. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Wu, J. Thermal-aided phosvitin extraction from egg yolk. J. Sci. Food Agric. 2015, 95, 2595–2600. [Google Scholar] [CrossRef]
- Ren, J.; Li, Q.; Offengenden, M.; Wu, J. Preparation and characterization of phosphopeptides from egg yolk phosvitin. J. Funct. Foods 2015, 18, 190–197. [Google Scholar] [CrossRef]
- Rampersad, S.N. Multiple applications of Alamar Blue as an indicator of metabolic function and cellular health in cell viability bioassays. Sensors 2012, 12, 12347–12360. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Davidge, S.T. Analysis of G-Protein Coupled Receptor 30 (GPR30) on Endothelial Inflammation. Methods Mol. Biol. 2016, 1366, 503–516. [Google Scholar]
- Majumder, K.; Chakrabarti, S.; Davidge, S.T.; Wu, J. Structure and activity study of egg protein ovotransferrin derived peptides (IRW and IQW) on endothelial inflammatory response and oxidative Stress. J. Agric. Food Chem. 2013, 61, 2120–2129. [Google Scholar] [CrossRef] [PubMed]
- Jahandideh, F.; Chakrabarti, S.; Majumder, K.; Li, Q.; Panahi, S.; Morton, J.S.; Davidge, S.T.; Wu, J. Egg white protein hydrolysate reduces blood pressure, improves vascular relaxation and modifies aortic angiotensin II receptors expression in spontaneously hypertensive rats. J. Funct. Foods 2016, 27, 667–673. [Google Scholar] [CrossRef] [Green Version]
- Flowers, S.; Patel, P.J.; Gleicher, S.; Amer, K.; Himelman, E.; Goel, S.; Moran, E. p107-dependent recruitment of SWI/SNF to the alkaline phosphatase promoter during osteoblast differentiation. Bone 2014, 69, 47–54. [Google Scholar] [CrossRef]
- Nakashima, Y.; Haneji, T. Stimulation of osteoclast formation by RANKL requires interferon regulatory factor-4 and is inhibited by simvastatin in a mouse model of bone loss. PLoS ONE 2013, 8, e72033. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chen, Q.; Xia, X.; Mo, P.; Shen, Y.; Yu, C. Mycoepoxydiene suppresses RANKL-induced osteoclast differentiation and reduces ovariectomy-induced bone loss in mice. Appl. Microbiol. Biotechnol. 2013, 97, 767–774. [Google Scholar] [CrossRef]
- Hou, J.M.; Xue, Y.; Lin, Q.M. Bovine lactoferrin improves bone mass and microstructure in ovariectomized rats via OPG/RANKL/RANK pathway. Acta Pharmacol. Sin. 2012, 33, 1277–1284. [Google Scholar] [CrossRef] [Green Version]
- Florencio-Silva, R.; Sasso, G.R.; Sasso-Cerri, E.; Simoes, M.J.; Cerri, P.S. Biology of bone tissue: Structure, function, and factors that influence bone cells. Biomed. Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malone, S.R. Vitamin D: Nutrition, Side Effects, and Supplements; Hauppauge: New York, NY, USA, 2010. [Google Scholar]
- Wilczynski, C.; Camacho, P. Calcium use in the management of osteoporosis: Continuing questions and controversies. Curr. Osteoporos. Rep. 2014, 12, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Ilich, J.Z.; Kerstetter, J.E. Nutrition in bone health revisited: A story beyond calcium. J. Am. Coll. Nutr. 2000, 19, 715–737. [Google Scholar] [CrossRef] [PubMed]
- Grant, W.B.; Holick, M.F. Benefits and requirements of vitamin D for optimal health: A review. Altern. Med. Rev. 2005, 10, 94–111. [Google Scholar] [PubMed]
- Mardon, J.; Zangarelli, A.; Walrand, S.; Davicco, M.J.; Lebecque, P.; Demigne, C.; Horcajada, M.N.; Boirie, Y.; Coxam, V. Impact of energy and casein or whey protein intake on bone status in a rat model of age-related bone loss. Br. J. Nutr. 2008, 99, 764–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donida, B.M.; Mrak, E.; Gravaghi, C.; Villa, I.; Cosentino, S.; Zacchi, E.; Perego, S.; Rubinacci, A.; Fiorilli, A.; Tettamanti, G.; et al. Casein phosphopeptides promote calcium uptake and modulate the differentiation pathway in human primary osteoblast-like cells. Peptides 2009, 30, 2233–2241. [Google Scholar] [CrossRef]
- Tsuchita, H.; Goto, T.; Shimizu, T.; Yonehara, Y.; Kuwata, T. Dietary casein phosphopeptides prevent bone loss in aged ovariectomized rats. J. Nutr. 1996, 126, 86–93. [Google Scholar] [CrossRef]
- Yuan, Y.V.; Kitts, D.D. Calcium absorption and bone utilization in spontaneously hypertensive rats fed on native and heat-damaged casein and soya-bean protein. Br. J. Nutr. 1994, 71, 583–603. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Li, C.; Geng, F.; Huang, X.; Ma, M. Hen egg yolk phosvitin stimulates osteoblast differentiation in the absence of ascorbic acid. J. Sci. Food Agric. 2017, 97, 4532–4538. [Google Scholar] [CrossRef]
- Jie, Y.; Li, X.; Cai, Z.; Ma, M.; Jin, Y.; Ahn, D.U.; Huang, X. Phosphorylation of phosvitin plays a crucial effects on the protein-induced differentiation and mineralization of osteoblastic MC3T3-E1 cells. Int. J. Biol. Macromol. 2018, 118, 1848–1854. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Li, X.; Hu, W.; Zeng, B.; Liang, R.; Liu, H.; Li, Z.; Zhang, Z. Phosvitin phosphopeptide preparation using immobilised trypsin and enhancing calcium absorption in growing rats. Czech J. Food Sci. 2016, 34, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Yoo, H.; Bamdad, F.; Gujral, N.; Suh, J.W.; Sunwoo, H. High hydrostatic pressure-assisted enzymatic treatment improves antioxidant and anti-inflammatory properties of phosvitin. Curr. Pharm. Biotechnol. 2017, 18, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Arora, H.; Shang, N.; Bhullar, K.S.; Wu, J. Pea protein-derived tripeptide LRW shows osteoblastic activity on MC3T3-E1 cells via the activation of the Akt/Runx2 pathway. Food Funct. 2020, 11, 7197–7207. [Google Scholar] [CrossRef] [PubMed]
- Reddi, S.; Shanmugam, V.P.; Kapila, S.; Kapila, R. Identification of buffalo casein-derived bioactive peptides with osteoblast proliferation activity. Eur. Food Res. Technol. 2016, 242, 2139–2146. [Google Scholar] [CrossRef]
- Pandey, M.; Kapila, S.; Kapila, R.; Trivedi, R.; Karvande, A. Evaluation of the osteoprotective potential of whey derived-antioxidative (YVEEL) and angiotensin-converting enzyme inhibitory (YLLF) bioactive peptides in ovariectomised rats. Food Funct. 2018, 9, 4791–4801. [Google Scholar] [CrossRef]
- Shang, N.; Bhullar, K.S.; Hubbard, B.P.; Wu, J. Tripeptide IRW initiates differentiation in osteoblasts via the RUNX2 pathway. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 1138–1146. [Google Scholar] [CrossRef]
- Akkouch, A.; Zhang, Z.; Rouabhia, M. Engineering bone tissue using human dental pulp stem cells and an osteogenic collagen-hydroxyapatite-poly (L-lactide-co-epsilon-caprolactone) scaffold. J. Biomater. Appl. 2014, 28, 922–936. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chakrabarti, S.; Ren, J.; Wu, J. Phosvitin Derived Phospho-Peptides Show Better Osteogenic Potential than Intact Phosvitin in MC3T3-E1 Osteoblastic Cells. Nutrients 2020, 12, 2998. https://doi.org/10.3390/nu12102998
Chakrabarti S, Ren J, Wu J. Phosvitin Derived Phospho-Peptides Show Better Osteogenic Potential than Intact Phosvitin in MC3T3-E1 Osteoblastic Cells. Nutrients. 2020; 12(10):2998. https://doi.org/10.3390/nu12102998
Chicago/Turabian StyleChakrabarti, Subhadeep, Jiandong Ren, and Jianping Wu. 2020. "Phosvitin Derived Phospho-Peptides Show Better Osteogenic Potential than Intact Phosvitin in MC3T3-E1 Osteoblastic Cells" Nutrients 12, no. 10: 2998. https://doi.org/10.3390/nu12102998
APA StyleChakrabarti, S., Ren, J., & Wu, J. (2020). Phosvitin Derived Phospho-Peptides Show Better Osteogenic Potential than Intact Phosvitin in MC3T3-E1 Osteoblastic Cells. Nutrients, 12(10), 2998. https://doi.org/10.3390/nu12102998