Macroalgae—A Sustainable Source of Chemical Compounds with Biological Activities



 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Algae Chemical Composition
2.1. Protein and Amino Acid Composition
2.2. Lipid and Fatty Acid Composition
2.3. Carbohydrates
2.4. Pigments
2.4.1. Carotenoids
2.4.2. Chlorophylls
2.5. Phenolic Compounds
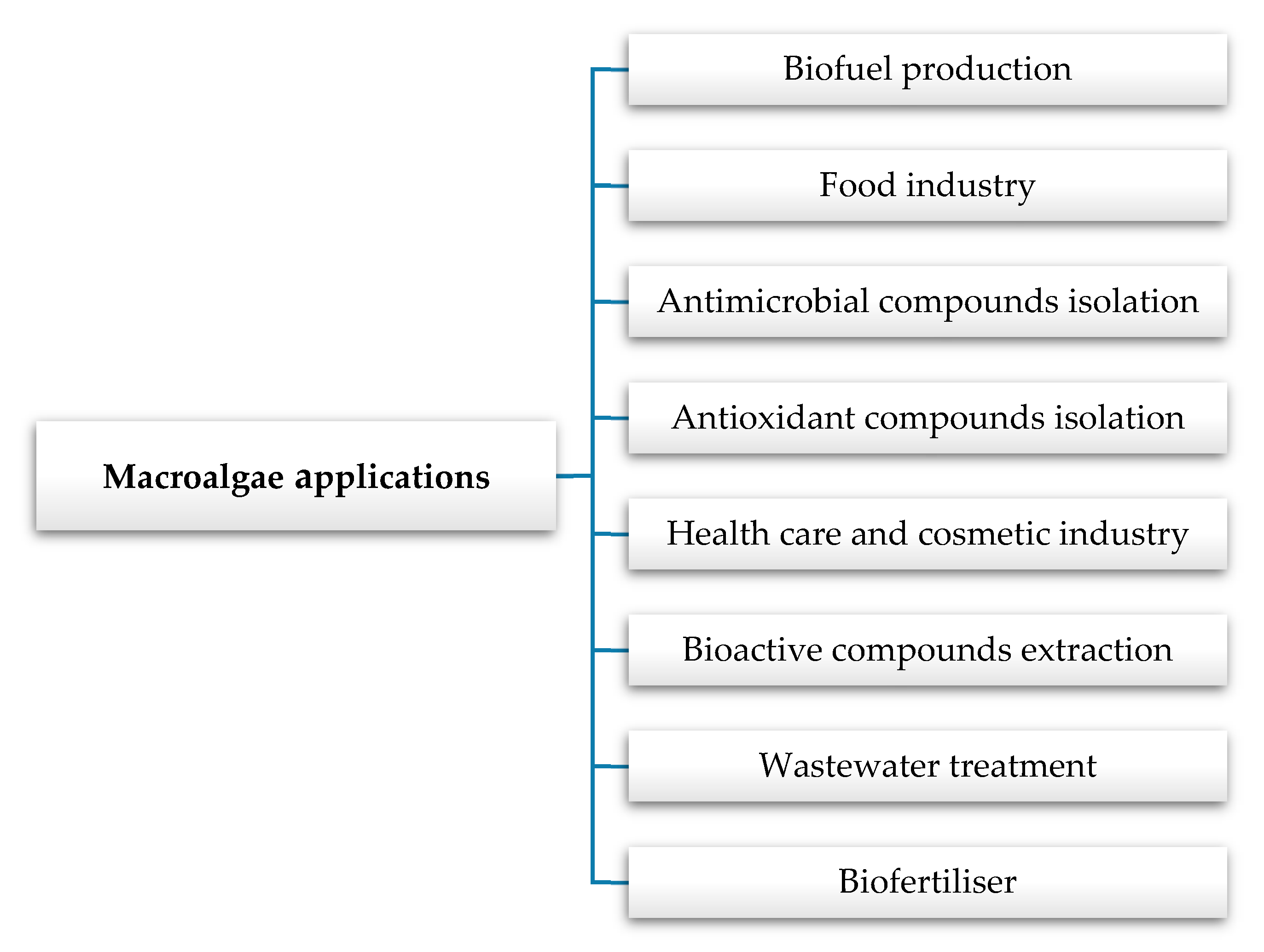
3. Algae Applications in the Food Industry
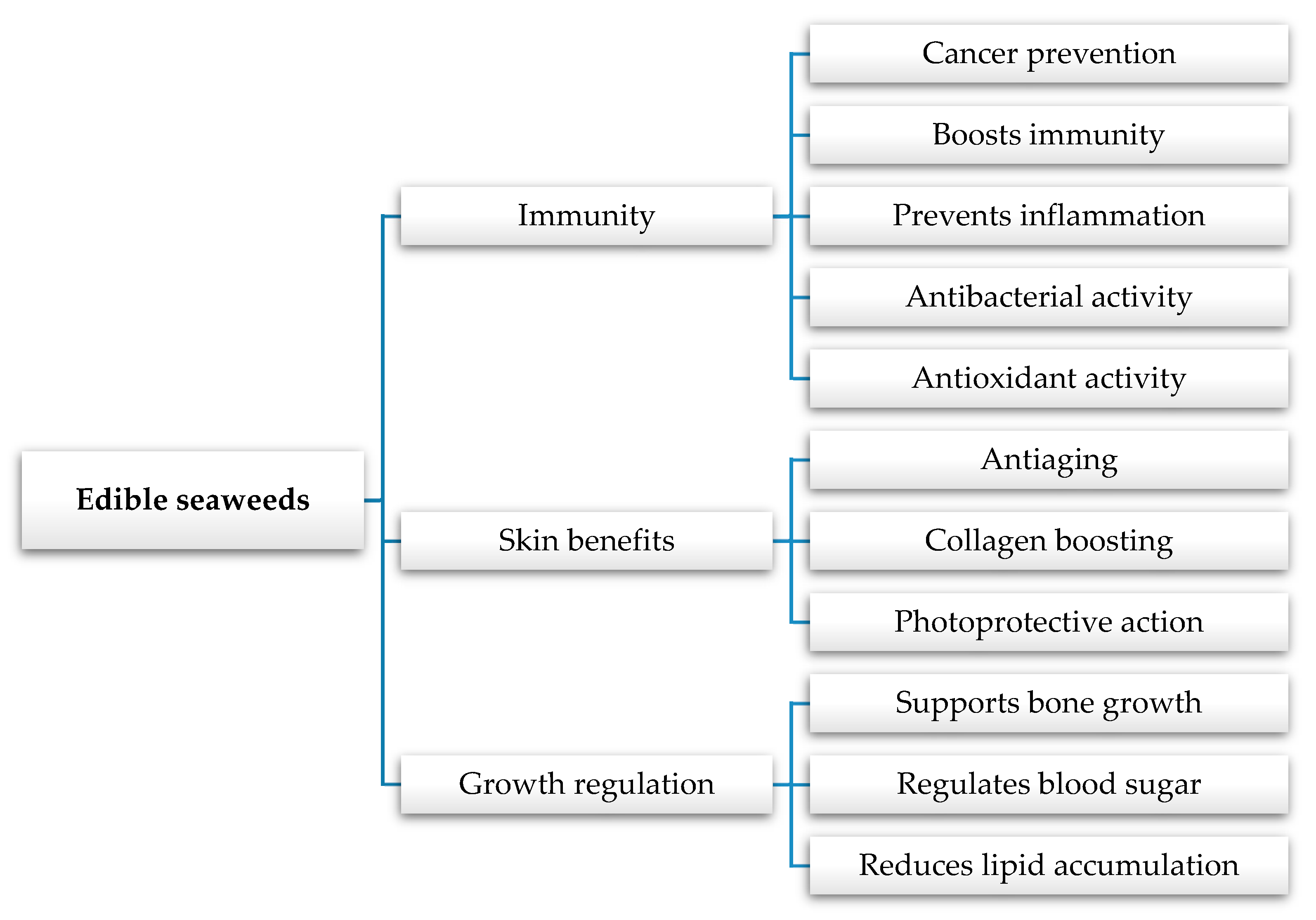
4. Health Effects
4.1. Blood Pressure, Sugar, and Fat Reduction
4.2. Anticoagulant and Antithrombotic Properties
4.3. Antiaging, Antidepression, and Antifatigue Properties
4.4. Antimicrobial and Antioxidant Potential
4.5. Antiallergic Effect
4.6. Anticancer Properties
4.7. Anti-Inflammatory Property
4.8. Antifungal Effects
4.9. Antiviral Effects
5. Conclusions and Future Trends
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brodie, J.; Lewis, J. Introduction in unravelling the algae: The past, present, and future of algal systematics. In Unravelling the Algae: The Past, Present, and Future of Algal Systematics, 1st ed.; Brodie, J., Lewis, J., Eds.; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Skrovankova, S. Seaweed vitamins as nutraceuticals, Toldra, F. Ed. Adv. Food Nutr. Res. 2011, 64, 357–369. [Google Scholar] [PubMed]
- Mahadevan, K. Seaweeds: A sustainable food source. In Seaweed Sustainability, 1st ed.; Tiwari, B.K., Troy, D.J., Eds.; Academic Press: London, UK, 2015; pp. 347–364. [Google Scholar]
- Bueno, A.M.; Marcílio Candido, T.; Morocho-Jácome, A.L.; Rolim Baby, A.; Robles Velasco, M.V.; M de Carvalho, J.C. Cosmetic attributes of algae–A review. Algal Res. 2017, 25, 483–487. [Google Scholar]
- Garcia-Vaquero, M.; Ummat, V.; Tiwari, B.; Rajauria, G. Exploring ultrasound, microwave and ultrasound–microwave assisted extraction technologies to increase the extraction of bioactive compounds and antioxidants from brown macroalgae. Mar. Drugs 2020, 18, 172. [Google Scholar] [CrossRef] [Green Version]
- Hayes, M. Seaweeds: A nutraceutical and health food. In Seaweed Sustainability, 1st ed.; Tiwari, B.K., Troy, D.J., Eds.; Academic Press: London, UK, 2015; pp. 365–387. [Google Scholar]
- Indira, K.; Balakrishnan, S.; Srinivasan, M.; Bragadeeswaran, S.; Balasubramanian, T. Evaluation of in vitro antimicrobial property of seaweed (Halimeda tuna) from Tuticorin coast, Tamil Nadu, Southeast coast of India. Afr. J. Biotechnol. 2013, 12, 284–289. [Google Scholar]
- Ganesan, A.; Tiwari, U.; Rajauria, G. Seaweed nutraceuticals and their role in disease prevention. Food Sci. Hum. Wellness. 2019, 8, 252–263. [Google Scholar] [CrossRef]
- Biris-Dorhoi, E.S. Cercetari Privind Utilizarea Algelor Marine in Bioremedierea Apelor Uzate Industrial–o Tehnica Eco-Friendly in Rezolvarea Poluarii Mediului; Mega: Cluj-Napoca, Romania, 2018; pp. 66–117. [Google Scholar]
- Negreanu-Pîrjol, B.; Negreanu-Pîrjol, T.; Paraschiv, G.-M.; Bratu, M.M.; Sirbu, R.; Roncea, F.; Meghea, A. Physical-chemical characterization of some green and red macrophyte algae from the Romanian Black Sea littoral. Sci. Study Res. Chem. Chem. Eng. 2011, 12, 173–184. [Google Scholar]
- Boukhari, E.L.; Barakate, M.; Bouhia, Y.; Lyamlouli, K. Trends in seaweed extract based biostimulants: Manufacturing process and beneficial effect on soil-plant systems. Plants 2020, 9, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbot, Y.N.; Hashem, A.; Benz, R. A Review on the valorization of macroalgal wastes for biomethane production. Mar. Drugs 2016, 14, 120. [Google Scholar] [CrossRef] [Green Version]
- Mæhre, K.H.; Malde, M.K.; Eilertsen, K.-E.; Elvevoll, E.O. Characterization of protein, lipid and mineral contents in common Norwegian seaweeds and evaluation of their potential as food and feed. J. Sci. Food Agric. 2014, 94, 3281–3290. [Google Scholar] [CrossRef]
- Kazir, M.; Abuhassira, Y.; Robin, A.; Nahor, O.; Luo, J.; Israel, A.; Golberg, A.; Livney, Y.D. Extraction of proteins from two marine macroalgae, Ulva sp. and Gracilaria sp., for food application, and evaluating digestibility, amino acid composition and antioxidant properties of the protein concentrates. Food Hydrocol. 2019, 87, 197–203. [Google Scholar] [CrossRef]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Hamid, N.; Ma, Q.; Boulom, S.; Liu, T.; Zheng, Z.; Balbas, J.; Robertson, J. Seaweed minor constituents. In Seaweed Sustainability, 1st ed.; Tiwari, B.K., Troy, D.J., Eds.; Academic Press: London, UK, 2015; pp. 193–242. [Google Scholar]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Rizzo, G.; Laganà, A.S.; Chiara Rapisarda, A.M.; Grazia La Ferrera, G.M.; Buscema, M.; Rossetti, P.; Nigro, A.; Muscia, V.; Valenti, G.; Sapia, F.; et al. Vitamin B12 among vegetarians: Status, assessment and supplementation. Nutrients 2016, 8, 767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, D.; Freitas, A.C.; Pereira, L.; Rocha-Santos TA, P.; Vasconcelos, M.W.; Roriz, M.; Rodriguez-Alcala, L.M.; Gomes AM, P.; Duarte, A.C. Chemical composition of red, brown and green macroalgae from Buarcos bay in Central West Coast of Portugal. Food Chem. 2015, 183, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Fleurence, J.; Morançais, M.; Dumay, J. Seaweed proteins. In Proteins in Food Processing, 2nd ed.; Yada, R.Y., Ed.; Woodhead Publishing: Cambridge, UK, 2018; pp. 245–262. [Google Scholar]
- Fleurence, J.; Gall, E.A. Antiallergic Properties. In Seaweed in Health and Disease Prevention, 1st ed.; Fleurence, J., Levine, I., Eds.; Academic Press: London, UK, 2016; pp. 389–420. [Google Scholar]
- Dawczynski, C.; Schubert, R.; Jahreis, G. Amino acids, fatty acids, and dietary fibre in edible seaweed products. Food Chem. 2007, 103, 891–899. [Google Scholar] [CrossRef]
- Biancarosa, I.; Belghit, I.; Bruckner, C.G.; Liland, N.S.; Waagbo, R.; Amlund, H.; Heesch, S.; Lock, E.J. Chemical characterization of 21 species of marine macroalgae common in Norwegian waters: Benefits of and limitations to their potential use in food and feed. J. Sci. Food Agric. 2018, 98, 2035–2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, S.-M.; Kim, H.-J.; Woo, M.-N.; Lee, M.-K.; Chul Shin, Y.; Bok Park, Y.; Choi, M.-S. Fucoxanthin-rich seaweed extract suppresses body weight gain and improves lipid metabolism in high-fat-fed C57BL/6J mice. Biotechnol. J. 2010, 5, 961–969. [Google Scholar] [CrossRef]
- Peng, Y.; Hu, J.; Yang, B.; Lin, X.-P.; Zhou, X.-F.; Yang, X.-W.; Liu, Y. Chemical composition of seaweeds. In Seaweed Sustainability; Tiwari, B.K., Troy, D.J., Eds.; Academic Press: London, UK, 2015; pp. 79–124. [Google Scholar]
- Jayasinghe, G.D.T.M.; Jinadasa, B.K.K.K.; Chinthaka, S.D.M. Study on lipid content and fatty acid profile of four marine macro algae (Seaweeds) collected from South East Coast of Sri Lanka. Asian J. Chem. Pharm. Sci. 2018, 1, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Kellogg, J.; Esposito, D.; Grace, M.H.; Komarnytsky, S.; Lila, M.A. Alaskan seaweeds lower inflammation in RAW 264.7 macrophages and decrease lipid accumulation in 3T3-L1 adipocytes. J. Func. Foods 2015, 15, 396–407. [Google Scholar] [CrossRef]
- Ma, A.-C.; Chen, Z.; Wang, T.; Song, N.; Yan, Q.; Fang, Y.-C.; Guan, H.-S.; Liu, H.-B. Isolation of the molecular species of monogalactosyldiacylglycerols from brown edible seaweed Sargassum horneri and their inhibitory effects on triglyceride accumulation in 3T3-L1 adipocytes. J. Agric. Food Chem. 2014, 62, 11157–11162. [Google Scholar] [CrossRef]
- Tsai, C.-J.; Sun Pan, B.A. Identification of sulfoglycolipid bioactivities and characteristic fatty acids of marine macroalgae. J. Agric. Food Chem. 2012, 60, 8404–8410. [Google Scholar] [CrossRef] [PubMed]
- Ramola, B.; Kumar, V.; Nanda, M.; Mishra, Y.; Tyagi, T.; Gupta, A.; Sharma, N. Evaluation, comparison of different solvent extraction, cell disruption methods and hydrothermal liquefaction of Oedogonium macroalgae for biofuel production. Biotech. Rep. 2019, 22, e00304. [Google Scholar] [CrossRef] [PubMed]
- Margareta, W.; Nagarajan, D.; Changa, J.S.; Lee, D.J. Dark fermentative hydrogen production using macroalgae (Ulva sp.) as the renewable feedstock. Appl. Energy 2020, 262, 114547. [Google Scholar] [CrossRef]
- Hofheins, J. An overview of macronutrients. In Essentials of Sport Nutrition and Supplements; Antonio, J., Kalman, D., Stout, J.R., Greenwood, M., Willoughby, D.S., Haff, G.G., Eds.; Humana Press: Totowa, NJ, USA, 2008; pp. 237–249. [Google Scholar]
- Praveen, M.A.; Karthika Parvathy, K.R.; Balasubramanian, P.; Jayabalan, R. An overview of extraction and purification techniques of seaweed dietary fibers for immunomodulation on gut microbiota. Trends Food Sci. Technol. 2019, 92, 46–64. [Google Scholar] [CrossRef]
- Patel, S.; Goyal, A. Functional oligosaccharides: Production, properties and applications. World J. Microbiol. Biotechnol. 2011, 27, 1119–1128. [Google Scholar] [CrossRef]
- Fleita, D.; El-Sayed, M.; Rifaat, D. Evaluation of the antioxidant activity of enzymatically-hydrolyzed sulfated polysaccharides extracted from red algae Pterocladia capillacea. LWT–Food Sci. Technol. 2015, 63, 1236–1244. [Google Scholar] [CrossRef]
- Xu, J.; Xu, L.-L.; Zhou, Q.-W.; Hao, S.-X.; Zhou, T.; Xie, H.-J. Isolation, purification, and antioxidant activities of degraded polysaccharides from Enteromorpha prolifera. Int. J. Biol. Macromol. 2015, 81, 1026–1030. [Google Scholar] [CrossRef]
- Koutsaviti, A.; Ioannou, E.; Roussis, V. Bioactive seaweed substances. In Bioactive Seaweeds for Food Applications, 1st ed.; Quin, Y., Ed.; Academic Press: London, UK, 2018; pp. 25–52. [Google Scholar]
- Wang, L.; Park, Y.; Jeon, Y.; Ryu, B. Bioactivities of the edible brown seaweed, Undaria pinnatifida: A review. Aquaculture 2018, 495, 873–880. [Google Scholar] [CrossRef]
- Lafarga, T.; Acién-Fernándeza, F.G.; Garcia-Vaquero, M. Bioactive peptides and carbohydrates from seaweed for food applications: Natural occurrence, isolation, purification, and identification. Algal. Res. 2020, 48, 101909. [Google Scholar] [CrossRef]
- Leandro, A.; Pacheco, D.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A. Seaweed’s bioactive candidate compounds to food industry and global food security. Life (Basel) 2020, 10, 140. [Google Scholar] [CrossRef]
- Ullah, S.; Khalil, A.A.; Shaukat, F.; Song, Y. Sources, extraction and biomedical properties of polysaccharides. Foods 2019, 8, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatesan, J.; Sukumaran, A.; Kim, S.-K. Introduction to seaweed polysaccharides. In Seaweed Polysaccharides, 1st ed.; Venkatesan, J., Anil, S., Kim, S.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–9. [Google Scholar]
- Bouanati, T.; Colson, E.; Moins, S.; Cabrera, J.C.; Eeckhaut, I.; Raquez, J.M.; Gerbaux, P. Microwave-assisted depolymerization of carrageenans from Kappaphycus alvarezii and Eucheuma spinosum: Controlled and green production of oligosaccharides from the algae biomass. Algal. Res. 2020, 51, 102054. [Google Scholar] [CrossRef]
- Vera, J.; Castro, J.; Gonzalez, A.; Moenne, A. Seaweed polysaccharides and derived oligosaccharides stimulate defense responses and protection against pathogens in plants. Mar. Drugs 2011, 9, 2514–2525. [Google Scholar] [CrossRef] [PubMed]
- Ueno, M.M.N.; Siddiqui, R.; Watashi, K.; Wakita, T.; Kato, N.; Ikeda, M.; Okimura, T.; Isaka, S.; Oda, T.; Ariumi, Y. Acidic polysaccharides isolated from marine algae inhibit the early step of viral infection. Int. J. Biol. Macromol. 2019, 124, 282–290. [Google Scholar] [CrossRef]
- Dumay, J.; Morançais, M. Proteins and Pigments. In Seaweed in Health and Disease Prevention, 1st ed.; Fleurence, J., Levine, I., Eds.; Academic Press: London, UK, 2016; pp. 275–318. [Google Scholar]
- Koyande, A.K.; Chewa, K.W.; Rambabub, K.; Taoc, Y.; Chud, D.T.; Show, P.L. Microalgae: A potential alternative to health supplementation for humans. Food Sci. Hum. Well. 2019, 8, 16–24. [Google Scholar] [CrossRef]
- Aryee AN, A.; Agyei, D.; Akanbi, T.O. Recovery and Utilization of Seaweed Pigments in Food Processing. Curr. Opin. Food Sci. 2018, 19, 113–119. [Google Scholar] [CrossRef]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [Green Version]
- Pangestuti, R.; Kim, S.K. Carotenoids, bioactive metabolites derived from seaweeds. In Springer Handbook of Marine Biotechnology; Kim, S.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 815–821. [Google Scholar]
- Prasanna, K.I.; King, P.; Prasad, V.S.R.K. Adsorption of zinc from aqueous solution using marine green algae—Ulva fasciata sp. Chem. Eng. J. 2007, 129, 161–166. [Google Scholar] [CrossRef]
- Pádua, D.; Rocha, E.; Gargiulo, D.; Ramos, A.A. Bioactive compounds from brown seaweeds: Phloroglucinol, fucoxanthin and fucoidan as promising therapeutic agents against breast cancer. Phytochem. Lett. 2015, 14, 91–98. [Google Scholar] [CrossRef]
- Erdogan, A.; Demirel, Z.; Dalay, M.; Eroglu, A.E. Fucoxanthin content of Cylindrotheca closterium and its oxidative stress mediated enhancement. Turk. J. Fish. Aquat. Sci. 2016, 16, 491–498. [Google Scholar] [CrossRef]
- Kim, K.N.; Heo, S.J.; Yoon, W.J.; Kang, S.M.; Ahn, G.; Yi, T.H.; Jeon, Y.J. Fucoxanthin inhibits the inflammatory response by suppressing the activation of NF-κB and MAPKs in lipopolysaccharide-induced RAW 264.7 macrophages. Eur. J. Pharmacol. 2010, 649, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Chung, T.W.; Choi, H.J.; Lee, J.Y.; Jeong, H.S.; Kim, C.H.; Joo, M.; Choi, J.Y.; Han, C.W.; Kim, S.Y.; Choi, J.S.; et al. Marine algal fucoxanthin inhibits the metastatic potential of cancer cells. Biochem. Biophys. Res. Commun. 2013, 439, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.J. Fucoxanthin and its metabolite fucoxanthinol in cancer prevention and treatment. Mar. Drugs 2015, 13, 4784–4798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, E.; Wang, L.J.; Jiang, B.; Riaz, S.; Butt, G.Y.; Shi, D.Y. A review of the components of brown seaweed as potential candidates in cancer therapy. RSC Adv. 2016, 6, 12592–12610. [Google Scholar] [CrossRef]
- Aki, T.; Yamamoto, M.; Takahashi, K.; Tomita, T.; Rieko, T.; Kazuhiro, I.; Seji, K.; Masashi, H.; Kazuo, M.; Kazuhisa, O. Regulation of polyunsaturated fatty acid biosynthesis by seaweed fucoxanthin and its metabolite in cultured hepatocytes. Lipids 2014, 49, 133–141. [Google Scholar] [CrossRef]
- Pereira, D.M.; Valentão, P.; Andrade, P.B. Marine natural pigments: Chemistry, distribution and analysis. Dye Pigment. 2014, 111, 124–134. [Google Scholar] [CrossRef]
- Murray, M.; Dordevic, A.L.; Ryan, L.; Bonham, M.P. An emerging trend in functional foods for the prevention of cardiovascular disease and diabetes: Marine algal polyphenols. Crit. Rev. Food Sci. Nutr. 2018, 58, 1342–1358. [Google Scholar] [CrossRef]
- Gupta, S.; Cox, S.; Rajauria, G.; Abu-Ghannam, N.; Jaiswal, A.K. Growth inhibition of common food spoilage and pathogenic microorganisms in the presence of brown seaweed extracts. Food Bioproc. Tech. 2010, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Senthilkumar, P.; Sudha, S. Antioxidant and antibacterial properties of methanolic extract of green seaweed Chaetomorpha linum from Gulf of Mannar: Southeast Coast of India. Jundishapur J. Microbiol. 2012, 5, 411–415. [Google Scholar] [CrossRef] [Green Version]
- Cotas, J.; Leandro, A.; Monteiro, P.; Pacheco, D.; Figueirinha, A.; Gonçalves AM, M.; da Silva, G.J.; Pereira, L. Seaweed phenolics: From extraction to applications. Mar. Drugs 2020, 18, 384. [Google Scholar] [CrossRef]
- Santos, S.; Félix, R.; Pais, A.; Rocha, S.M.; Silvestre, A. The quest for phenolic compounds from macroalgae: A review of extraction and identification methodologies. Biomolecules 2019, 9, 847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ścieszka, S.; Klewicka, E. Algae in food: A general review, Critical Reviews. Food Sci. Nutr. 2019, 59, 3538–3547. [Google Scholar]
- Cofrades, S.; Benedí, J.; Garcimartin, A.; Sánchez-Muniz, F.J.; Jimenez-Colmenero, F. A comprehensive approach to formulation of seaweed-enriched meat products: From technological development to assessment of healthy properties. Food Res. Internat. 2017, 99, 1084–1094. [Google Scholar] [CrossRef] [PubMed]
- Garicano Vilar, E.; Ouyang, H.; O’Sullivan, M.G.; Kerry, J.P.; Hamill, R.M.; O’Grady, M.N.; Mohammed, H.O.; Kilcawley, K.N. Effect of salt reduction and inclusion of 1% edible seaweeds on the chemical, sensory and volatile component profile of reformulated frankfurter. Meat Sci. 2020, 161, 108001. [Google Scholar] [CrossRef] [PubMed]
- Roohinejad, S.; Kouba, M.; Barba, F.J.; Saljoughian, S.; Amid, M.; Greiner, R. Application of seaweeds to develop new food products with enhanced shelf-life, quality and health-related beneficial properties. Food Res. Internat. 2017, 99, 1066–1083. [Google Scholar] [CrossRef]
- Cofrades, S.; Serdaroglu, M.; Jiménez-Colmenero, F. Design of healthier foods and beverages containing whole algae. In Functional Ingredients from Algae for Foods and Nutraceuticals, 1st ed.; Dominguez, H., Ed.; Woodhead Publishing: Cambridge, UK, 2013; pp. 609–633. [Google Scholar]
- Lalic, L.M.; Berkovic, K. The influence of algae addition on physicochemical properties of cottage cheese. Milchwissenschaft 2005, 60, 151–154. [Google Scholar]
- Hopkins, D.L.; Clayton, E.H.; Lamb, T.A.; van de Ven, R.J.; Refshauge, G.; Kerr, M.J.; Bailes, K.; Lewandowski, P.; Ponnampalam, E.N. The impact of supplementing lambs with algae on growth, meat traits and oxidative status. Meat Sci. 2014, 98, 135–141. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Burnett, F.V.; Norng, S.; Hopkins, D.L.; Plozza, T.; Jacobs, J.L. Muscle antioxidant (vitamin E) and major fatty acid groups, lipid oxidation and retail colour of meat from lambs fed a roughage-based diet with flaxseed or algae. Meat Sci. 2015, 111, 154–160. [Google Scholar] [CrossRef]
- Bortoluzzi, C.; Schmidt, J.M.; Bordignon, H.L.F.; Fülber, L.M.; Layter, J.R.; Fernandes, J.I.M. Efficacy of yeast derived glucomannan or algae-based antioxidant or both as feed additives to ameliorate mycotoxicosis in heat stressed and unstressed broiler chickens. Livest. Sci. 2016, 193, 20–25. [Google Scholar] [CrossRef]
- Xiao, R.; Power, R.F.; Mallonee, D.; Crowdus, C.; Brennan, K.M.; Ao, T.; Pierce, J.L.; Dawson, K.A. A comparative transcriptomic study of vitamin E and an algae-based antioxidant as antioxidative agents: Investigation of replacing vitamin E with the algae-based antioxidant in broiler diets. Poult. Sci. 2011, 90, 136–146. [Google Scholar] [CrossRef]
- Rico, D.; Martín Diana, A.B.; Milton-Laskibar, I.; Fernández-Quintela, A.; Silván, J.M.; Rai, D.K.; Choudhary, A.; Peñas, E.; de Luis, D.A.; Martínez-Villalueng, C. Characterization and in vitro evaluation of seaweed species as potential functional ingredients to ameliorate metabolic syndrome. J. Funct. Foods 2018, 46, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Charoensiddhi, S.; Abraham, R.E.; Su, P.; Zhang, W. Seaweed and seaweed-derived metabolites as prebiotics. Adv. Food Nutr. Res. 2020, 91, 97–156. [Google Scholar] [PubMed]
- Wang, Y.; Han, F.; Hu, B.; Li, J.B.; Yu, W.G. In vivo prebiotic properties of alginate oligosaccharides prepared through enzymatic hydrolysis of alginate. Nut. Res. 2006, 26, 597–603. [Google Scholar] [CrossRef]
- Kuda, T.; Yano, T.; Matsuda, N.; Nishizawa, M. Inhibitory effects of laminaran and low molecular alginate against the putrefactive compounds produced by intestinal microflora in vitro and in rats. Food Chem. 2005, 91, 745–749. [Google Scholar] [CrossRef]
- Gudiel-Urbano, M.; Goni, I. Effect of edible seaweeds (Undaria pinnatifida and Porphyra ternera) on the metabolic activities of intestinal microflora in rats. Nut. Res. 2002, 22, 323–331. [Google Scholar] [CrossRef]
- Baweja, P.; Kumar, S.; Sahoo, D.; Levine, I.A. Biology of Seaweeds. In Seaweed in Health and Disease Prevention, 1st ed.; Fleurence, J., Levine, I., Eds.; Academic Press: London, UK, 2016; pp. 41–106. [Google Scholar]
- Zaharudin, N.; Salmeán, A.A.; Dragsted, L.O. Inhibitory effects of edible seaweeds, polyphenolics and alginates on the activities of porcine pancreatic α-amylase. Food Chem. 2018, 245, 1196–1203. [Google Scholar] [CrossRef]
- Reboleira, J.; Freitas, R.; Pinteus, S.; Silva, J.; Alves, C.; Pedrosa, R.; Bernardino, S. Brown Seaweeds. In Nonvitamin and Nonmineral Nutritional Supplements; Nabavi, S.M., Silva, A.S., Eds.; Academic Press: London, UK, 2019; pp. 171–176. [Google Scholar]
- Rabanal, M.; Ponce NM, A.; Navarro, D.A.; Gómez, R.M.; Stortz, C.A. The system of fucoidans from the brown seaweed Dictyota dichotoma: Chemical analysis and antiviral activity. Carbohydr. Polym. 2014, 101, 804–811. [Google Scholar] [CrossRef]
- Shibneva, S.Y.; Skriptsova, A.V. Morphological variability of Undaria pinnatifida (Harvey) Suringar, 1873 (Phaeophyceae, Laminariales) in Peter the Great Bay, Sea of Japan. Russ. J. Mar. Biol. 2012, 38, 381–391. [Google Scholar] [CrossRef]
- Guidone, M.; Thornber, C.; Wysor, B.; O’Kelly, C.J. Molecular and morphological diversity of Narragansett Bay (RI, USA) Ulva (Ulvales, Chlorophyta) populations. J. Phycol. 2013, 49, 979–995. [Google Scholar] [CrossRef]
- Zhong, R.; Wan, X.; Wang, D.; Zhao, C.; Liu, D.; Gao, L.; Wang, M.; Wu, C.; Nabavid, S.M.; Daglia, M.; et al. Polysaccharides from marine Enteromorpha: Structure and function. Trends Food Sci. Technol. 2020, 99, 11–20. [Google Scholar] [CrossRef]
- Wijesinghe, W.A.J.P.; Jeon, Y.J. Exploiting biological activities of brown seaweed Ecklonia cava for potential industrial applications: A review. Int. J. Food Sci. Nutr. 2012, 63, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.A.; Lee, K.H.; Chae, S.; Zhang, R.; Jung, M.S.; Lee, Y.; Kim, S.Y.; Kim, H.S.; Joo, H.G.; Park, J.W.; et al. Eckol isolated from Ecklonia cava attenuates oxidative stress induced cell damage in lung fibroblast cells. FEBS Lett. 2005, 579, 6295–6304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, M.J.; Yoon, K.D.; Min, S.Y.; Lee, J.S.; Kim, J.H.; Kim, T.G.; Kim, S.H.; Kim, N.G.; Huh, H.; Kim, J. Inhibition of HIV-1 reverse transcriptase and protease by phlorotannins from the brown alga Ecklonia cava. Biol. Pharm. Bull. 2004, 27, 544–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Déléris, P.; Nazih, H.; Bard, J.-M. Seaweeds in Human Health. In Seaweed in Health and Disease Prevention, 1st ed.; Fleurence, J., Levine, I., Eds.; Academic Press: London, UK, 2016; pp. 319–367. [Google Scholar]
- Guh, D.P.; Zhang, W.; Bansback, N.; Amarsi, Z.; Birmingham, C.L.; Anis, A.H. The incidence of co-morbidities related to obesity and overweight: A systematic review and meta-analysis. BMC Public Health 2009, 9, 88. [Google Scholar] [CrossRef] [Green Version]
- Unnikrishnan, M.K.; Veerapur, V.; Nayak, Y.; Paul, P.; Mathew, G. Antidiabetic, antihyperlipidemic and antioxidant effects of the flavonoids. In Polyphenols in Human Death and Disease, 1st ed.; Watson, R., Preedy, V., Zibadi, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 143–161. [Google Scholar]
- Souza, R.B.; Frota, A.F.; Sousa, R.S.; Cezario, N.A.; Santos, T.B.; Souza LM, F.; da Cunha, R.M.S. Neuroprotective effects of sulphated agaran from marine alga Gracilaria cornea in rat 6-hydroxydopamine Parkinson’s disease model: Behavioural, neurochemical and transcriptional alterations. Basic Clin. Pharmacol. Toxicol. 2017, 120, 159–170. [Google Scholar] [CrossRef]
- Djousse´, L.; Arnett, D.K.; Pankow, J.S.; Hopkins, P.N.; Province, M.A.; Ellison, R.C. Dietary linolenic acid is associated with a lower prevalence of hypertension in the NHLBI family heart study. Hypertension 2005, 45, 368–373. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, S.M.; Pereira, O.R.; Seca, A.M.L.; Pinto, D.C.G.A.; Silva, A.M.S. Seaweeds as preventive agents for cardiovascular diseases: From nutrients to functional foods. Mar. Drugs 2015, 13, 6838–6865. [Google Scholar] [CrossRef] [Green Version]
- Yimin, Q. Health benefits of bioactive seaweed substances. In Bioactive Seaweeds for Food Applications, 1st ed.; Quin, Y., Ed.; Academic Press: London, UK, 2018; pp. 180–199. [Google Scholar]
- Hall, A.C.; Fairclough, A.C.; Mahadevan, K.; Paxman, J.R. Ascophyllum nodosum enriched bread reduces subsequent energy intake with no effect on post-prandial glucose and cholesterol in healthy, overweight males. A pilot study. Appetite 2012, 58, 379–438. [Google Scholar] [CrossRef] [Green Version]
- Odunsi, S.T.; Vázquez-Roque, M.I.; Camilleri, M.; Papathanasopoulos, A.; Clark, M.M.; Wodrich, L.; Lempke, M.; McKinzie, S.; Ryks, M.; Burton, D.; et al. Effect of alginate on satiation, appetite, gastric function, and selected gut satiety hormones in overweight and obesity. Obesity (Silver Spring) 2010, 18, 1579–1584. [Google Scholar] [CrossRef] [Green Version]
- Sharifuddin, Y.; Chin, Y.-X.; Lim, P.-E.; Phang, S.-M. Potential bioactive compounds from seaweed for diabetes management. Mar. Drugs 2015, 13, 5447–5491. [Google Scholar] [CrossRef] [Green Version]
- Umekawa, H.; Takuya, T.; Midori Hirayama, Y.W.; Hattori, M.; Araki, T. Effect of Porphyra extract on blood pressure in spontaneously hypertensive rats. Nippon Shokuhin Kagaku Kogaku Kaishi 2008, 55, 502–505. [Google Scholar] [CrossRef]
- Qu, W.; Ma, H.; Pan, Z.; Luo, L.; Wang, Z.; He, R. Preparation and antihypertensive activity of peptides from Porphyra yezoensis. Food Chem. 2010, 123, 14–20. [Google Scholar] [CrossRef]
- Jeong, Y.T.; Kim, Y.D.; Jung, Y.M.; Park, D.C.; Lee, D.S.; Ku, S.K.; Li, X.; Lu, Y.; Chao, G.H.; Kim, K.J.; et al. Low molecular weight fucoidan improves endoplasmic reticulum stress-reduced insulin sensitivity through AMP-activated protein kinase activation in L6 myotubes and restores lipid homeostasis in a mouse model of type 2 diabetes. Mol. Pharmacol. 2013, 84, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.V.; Tsou, Y.C.; Chen, Y.T.; Lu, W.J.; Hwang, P.A. Effects of low-molecular-weight fucoidan and high stability fucoxanthin on glucose homeostasis, lipid metabolism, and liver function in a mouse model of type II diabetes. Mar. Drugs 2017, 15, 113. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kim, J.; Park, Y. Trans-10, cis-12 conjugated linoleic acid enhances endurance capacity by increasing fatty acid oxidation and reducing glycogen utilization in mice. Lipids 2012, 47, 855–863. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Gerbst, A.G.; Ushakova, N.A.; Tsvetkova, E.A.; Dmitrenok, A.S.; Usov, A.I.; Nifantiev, N.E. Anticoagulant and antithrombotic activities of modified xylofucan sulfate from the brown alga Punctaria plantaginea. Carbohydr. Polym. 2016, 136, 826–833. [Google Scholar] [CrossRef]
- Fujimura, T.; Tsukahara, K.; Moriwaki, S.; Kitahara, T.; Sano, T.; Takema, Y. Treatment of human skin with an extract of Fucus vesiculosus changes its thickness and mechanical properties. J. Cosmet. Sci. 2002, 53, 1–9. [Google Scholar]
- Tian, T.; Chang, H.; He, K.; Ni, Y.; Li, C.; Hou, M.; Chen, L.; Xu, Z.; Chen, B.; Ji, M. Fucoidan from seaweed Fucus vesiculosus inhibits 2, 4-dinitrochlorobenzene-induced atopic dermatitis. Int. Immunopharmacol. 2019, 75, 105823. [Google Scholar] [CrossRef]
- Wang HM, D.; Chen, C.C.; Huynh, P.; Chang, J.S. Exploring the potential of using algae in cosmetics. Biores. Tech. 2015, 184, 355–362. [Google Scholar] [CrossRef]
- Kidd, P. Astaxanthin, cell membrane nutrient with diverse clinical benefits and anti-aging potential. Altern. Med. Rev. 2011, 16, 355–364. [Google Scholar]
- Barbosa, M.; Valentão, P.; Andrade, P.B. Bioactive compounds from macroalgae in the new millennium: Implications for neurodegenerative diseases. Mar. Drugs 2014, 12, 4934–4972. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Tanaka, K.; Okubo, H.; Sasaki, S.; Arakawa, M. Seaweed consumption and prevalence of depressive symptoms during pregnancy in Japan: Baseline data from the Kyushu Okinawa Maternal and Child Health Study. BMC Pregnancy Childbirth 2014, 14, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, J.T.; Wang, M.Y.; Zheng, L.B. Antifatigue effect of Gracilaria eucheumoides in mice. Exp. Ther. Med. 2013, 6, 1512–1516. [Google Scholar] [CrossRef] [PubMed]
- Parsaeimehr, A.; Lutzu, G.A. Algae as a Novel Source of Antimicrobial Compounds: Current and Future Perspectives. In Antibiotic Resistance, 1st ed.; Kon, K., Rai, M., Eds.; Academic Press: London, UK, 2016; pp. 377–413. [Google Scholar]
- Shanmughapriya, S.; Manilal, A.; Sujith, S.; Selvin, J.; Kiran, G.S.; Natarajaseenivasan, K. Antimicrobial activity of seaweeds extracts against multiresistant pathogens. Ann. Microbiol. 2008, 58, 535–541. [Google Scholar] [CrossRef]
- Prarthana, J.; Maruthi, K.R. Fresh water algae as a potential source of bioactive compounds for aquaculture and significance of solvent system in extraction of antimicrobials. Asian J. Sci. Res. 2019, 12, 18–28. [Google Scholar] [CrossRef]
- Tüney, I.; Çadirci, B.H.; Ünal, D.; Sukatar, A. Antimicrobial activities of the extracts of marine algae from the Coast of Urla (Izmir, Turkey). Turk. J. Biol. 2006, 30, 171–175. [Google Scholar]
- Bhuyar, P.; Rahim, M.H.; Sundararaju, S.; Maniam, G.P.; Govindan, N. Antioxidant and antibacterial activity of red seaweed; Kappaphycus alvarezii against pathogenic bacteria. Glob. J. Environ. Sci. Manag. 2020, 6, 47–58. [Google Scholar]
- Lim, C.W.; Leem J., S.; Cho, Y.J. Structures and some properties of the antimicrobial compounds in the red alga, Symphyocladia latiuscula. Kor. J. Fish Aquatic. Sci. 2000, 33, 280–287. [Google Scholar]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. An assessment of the antioxidant and antimicrobial activity of six species of edible Irish seaweeds. Int. Food Res. J. 2010, 17, 205–220. [Google Scholar]
- Sims, J.J.; Donnell, M.S.; Leary, J.V.; Lacy, G.H. Antimicrobial agents from marine algae. Antimicrob. Agents Chemother. 1975, 7, 320–321. [Google Scholar] [CrossRef] [Green Version]
- Gamal, A.A.E. Biological importance of marine algae. Saudi Pharm. J. 2010, 18, 1–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Saif, S.S.; Abdel-Raouf, N.; El-Wazanani, H.A.; Aref, I.A. Antibacterial substances from marine algae isolated from Jeddah coast of Red sea, Saudi Arabia. Saudi J. Biol. Sci. 2014, 21, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahidi, M.E.; Amraoui, B.E.; Amraoui, M.E.; Bamhaoud, T. Screening of antimicrobial activity of macroalgae extracts from the Moroccan Atlantic coast. Ann. Pharm. Fr. 2015, 73, 190–196. [Google Scholar] [CrossRef]
- Pina-Pérez, M.C.; Rivas, A.; Martínez, A.; Rodrigo, D. Antimicrobial potential of macro and microalgae against pathogenic and spoilage microorganisms in food. Food Chem. 2017, 235, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Sanmukh, S.G.; Benedict, B.; Udhaya, R.; Krishna, K.; Sandhya, S.; Waman, N.P. Bioactive compounds derived from microalgae showing antimicrobial activities. Aquaculture 2014, 5, 100224. [Google Scholar] [CrossRef] [Green Version]
- Danyal, A.; Mubeen, U.; Malik, K.A. Investigating two native algal species to determine antibiotic susceptibility against some pathogens. Curr. Res. J. Biol. Sci. 2013, 5, 70–74. [Google Scholar] [CrossRef]
- Lee, J.H. Antibacterial effect of marine algae against oral-borne patogens. Res. J. Med. Plants 2014, 8, 196–203. [Google Scholar]
- Jang, K.; Lee, J. Investigation of anti-bacterial activity against food-borne pathogens among korean domestic algae. Adv. J. Food Sci. Technol. 2015, 7, 490–495. [Google Scholar] [CrossRef]
- Nshimiyumukiza, O.; Kang, S.K.; Kim, H.J.; Mog, Y.; Lee, E.H.; Han, H.N.; Kim, Y.; Kim, D.H.; Kim, J.H.; Eom, S.H. Synergistic Antibacterial activity of Ecklonia cava (Phaeophyceae: Laminariales) against Listeria monocytogenes (Bacillales: Listeriaceae). Fish Aquac. Sci. 2015, 18, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Kim BW, R.; Lim, S.I.; Ahn, D.H. Antibacterial mechanism of Myagropsis myagroides extract on Listeria monocytogenes. Food Control 2014, 42, 23–28. [Google Scholar] [CrossRef]
- Rajauria, G. Optimization and validation of reverse phase HPLC method forqualitative and quantitative assessment of polyphenols in seaweed. J. Pharmaceut. Biomed. Anal. 2018, 148, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.G.; Kang, O.H.; Brice, O.O.; Lee, Y.S.; Chae, H.S.; Oh, Y.C.; Sohn, D.H.; Park, H.; Choi, H.G.; Kim, S.G.; et al. Antibacterial activity of Ecklonia cava against methicillin-resistant Staphylococcus aureus and Salmonella spp. Foodborne Pathog. Dis. 2010, 7, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.H.; Kim, Y.M.; Kim, S.-K. Antimicrobial effect of phlorotannins from marine brown algae. Food Chem. Toxicol. 2012, 50, 3251–3255. [Google Scholar] [CrossRef] [PubMed]
- Negreanu -Pirjol, T.; Negreanu -Pirjol, B.; Sirbu, R.; Paraschiva, G.M.; Meghea, A. Comparative studies regarding the antioxidative activity of some therapeutic marine algae species along the Romanian Black Sea Coast. J. Environ. Prot. Ecol. 2012, 13, 1744–1750. [Google Scholar]
- Ngo, D.H.; Kim, S.K. Marine bioactive peptides as potential antioxidantsm. Curr. Protein Pept. Sci. 2013, 14, 189–198. [Google Scholar] [CrossRef]
- Pimentel, F.B.; Alves, R.C.; Harnedy, P.A.; FitzGerald, R.J.; Oliveira, M.B.P.P. Macroalgal-derived protein hydrolysates and bioactive peptides: Enzymatic release and potential health enhancing properties. Trends Food Sci. Technol. 2019, 93, 106–124. [Google Scholar] [CrossRef]
- Ishihara, K.; Oyamada, C.; Matsushima, R.; Murata, M.; Muraoka, T. Inhibitory effect of porphyran, prepared from dried “Nori”, on contact hypersensitivity in mice. Biosci. Biotechnol. Biochem. 2005, 69, 1824–1830. [Google Scholar] [CrossRef]
- Sugiura, Y.; Matusda, K.; Yamada, Y. Anti-allergic phlorotannins from the edible alga, Eisenia arborea. Food Sci. Technol. Res. 2007, 13, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Vo, T.; Ngo, D.; Kim, S. Marine algae as a potential pharmaceutical source for anti-allergic therapeutics. Proc. Biochem. 2012, 47, 386–394. [Google Scholar] [CrossRef]
- Van Ginneken, V.J.; Helsper, J.P.; de Visser, W.; van Keulen, H.; Brandenburg, W.A. Polyunsaturated fatty acids in various macroalgal species from North Atlantic and tropical seas. Lipids Health Dis. 2011, 10, 104. [Google Scholar] [CrossRef] [Green Version]
- Sakthivel, R.; Pandima Devi, K. Antioxidant, anti-inflammatory and anticancer potential of natural bioactive compounds from seaweeds. In Studies in Natural Products Chemistry, 1st ed.; Rahman, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 63, pp. 113–154. [Google Scholar]
- Gamal-Eldeen, A.M.; Ahmed, E.F.; Abo-Zeid, M.A. In vitro cancer chemopreventive properties of polysaccharide extract from the brown alga, Sargassum latifolium. Food Chem. Toxicol. 2009, 47, 378–1384. [Google Scholar] [CrossRef] [PubMed]
- Ryu, M.J.; Kim, A.D.; Kang, K.A.; Chung, H.S.; Kim, H.S.; Suh, I.S.; Chang, W.Y.; Hyun, J.W. The green algae Ulva fasciata Delile extract induces apoptotic cell death in human colon cancer cells. In Vitro Cell Dev. Biol. Anim. 2013, 49, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Wang, Z.J.; Xie, D.; Sun, X.; Yang, W.; Zhao, X.; Xu, N. Characterization and potential antitumor activity of polysaccharide from Gracilariopsis lemaneiformis. Mar. Drugs 2017, 15, 100. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chiu, L.; Ooi, V.; Ang, P. A potent antitumor polysaccharide from the edible brown seaweed Hydroclathrus clathratus. Bot. Mar. 2010, 53, 265–274. [Google Scholar] [CrossRef]
- Chale-Dzul, J.; Freile-Pelegrín, Y.; Robledo, D.; Moo-Puc, R. Protective effect of fucoidans from tropical seaweeds against oxidative stress in HepG2 cells. J. Appl. Phycol. 2017, 29, 2229–2238. [Google Scholar] [CrossRef]
- Palanisamy, S.; Vinosha, M.; Marudhupandi, T.; Rajasekar, P.; Prabhu, N.M. Isolation of fucoidan from Sargassum polycystum brown algae: Structural characterization, in vitro antioxidant and anticancer activity. Int. J. Biol. Macromol. 2017, 102, 405–412. [Google Scholar] [CrossRef]
- Menshova, R.V.; Shevchenko, N.M.; Imbs, T.I.; Zvyagintseva, T.N.; Malyarenko, O.S.; Zaporoshets, T.S.; Ermakova, S.P. Fucoidans from brown alga Fucus evanescens: Structure and biological activity. Front. Mar. Sci. 2016, 3, 129. [Google Scholar] [CrossRef] [Green Version]
- Namvar, F.; Mohamed, S.; Fard, S.G.; Behravan, J.; Mustapha, N.M.; Alitheen NB, M.; Othman, F. Polyphenol-rich seaweed (Eucheuma cottonii) extract suppresses breast tumour via hormone modulation and apoptosis induction. Food Chem. 2012, 130, 376–382. [Google Scholar] [CrossRef]
- Reddy, P.; Urban, S. Meroditerpenoids from the southern australian marine brown alga Sargassum fallax. Phytochem. 2009, 70, 250–255. [Google Scholar] [CrossRef]
- Sornsiri, J.; Srisook, K.; Pornngam, P.; Sootanan, P. Prediction of biochemical mechanism of anti-inflammation explained from two marine-derived bioactive compounds. Agric. Nat. Res. 2018, 52, 588–595. [Google Scholar] [CrossRef]
- Kazlowska, K.; Hsu, T.; Hou, C.C.; Yang, W.C.; Tsai, G.J. Antiinflammatory properties of phenolic compounds and crude extract from Porphyra dentate. J. Ethnopharmacol. 2010, 128, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Bitencourt MA, O.; Dantas, G.R.; Lira, D.P.; Filho JM, B.; Miranda GE, C.; Santos BV, O.; Souto, J.T. Aqueous and methanolic extracts of Caulerpa mexicana suppress cell migration and ear edema induced by inflammatory agents. Mar. Drugs 2011, 9, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.J.; Yoon, W.Y.; Kim, K.N.; Ahn, G.N.; Kang, S.M.; Kang, D.H.; Affan, A.; Oh, C.; KyoJung, W.; JinJeon, Y. Evaluation of anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food Chem. Toxicol. 2010, 48, 2045–2051. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.D.; Hien, H.M.; Anh, H.T. Studies on the analgesic and anti-inflammatory activities of Sargassum swartzii (Turner), C. Agardh (Phaeophyta) and Ulva reticulata Forsskal (Chlorophyta) in experiment animal models. Afr. J. Biotechnol. 2011, 10, 2308–2314. [Google Scholar]
- Croci, D.O.; Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; Piccoli, A.; Totani, L.; Ustyuzhanina, N.E.; Bilan, M.I.; Usov, A.I.; Grachev, A.A. Fucans, but not fucomannoglucuronas, determine the biological activities of sulfated polysaccharides from Laminaria saccharina brown seaweed. PLoS ONE 2011, 6, e17283. [Google Scholar] [CrossRef] [Green Version]
- Robertson, R.C.; Guihéneuf, F.; Bahar, B.; Schmid, M.; Stengel, D.B.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. The anti-inflammatory effect of algae-derived lipid extracts on lipopolysaccharide (LPS)-stimulated human THP-1 macrophages. Mar. Drugs 2015, 13, 5402–5424. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues JA, G.; Oliveira Vanderlei ED, S.; Silva, L.M.; de Araujo, I.W.; de Queiroz, I.N.; de Paula, G.A.; Abreu, T.M.; Ribeiro, N.A.; Bezerra, M.M.; Chaves, H.V. Antinociceptive and anti-inflammatory activities of a sulfated polysaccharide isolated from the green seaweed Caulerpa cupressoides. Pharmacol. Rep. 2012, 64, 282–292. [Google Scholar] [CrossRef]
- Albuquerque, I.R.L.; Cordeiro, S.L.; Gomes, D.L.; Dreyfuss, J.L.; Filgueira, L.G.A.; Leite, E.L.; Nader, H.B.; Rocha, H.A.O. Evaluation of anti-nociceptive and anti-inflammatory activities of a heterofucan from Dyctiota menstrualis. Mar. Drugs 2013, 11, 2722–2740. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.I.; Jung, S.H.; Lee, K.T.; Choi, J.H. Dieckol isolated from edible brown algae Ecklonia cava induces apoptosis of ovarian cancer cells and inhibits tumor xenografr growth. J. Cancer Res. Clin. Oncol. 2015, 68, 141–255. [Google Scholar]
- Coura, C.O.; Souza, R.B.; Rodrigues, J.A.; Vanderlei Ede, S.; de Arau´ jo, I.W.; Ribeiro, N.A.; Frota, A.F.; Ribeiro, K.A.; Chaves, H.V. Mechanisms involved in the anti-inflammatory action of a poly sulfated fraction from Gracilaria cornea in rats. PLoS ONE 2015, 10, e0119319. [Google Scholar] [CrossRef]
- Isaka, S.; Cho, K.; Nakazono, S.; Abu, R.; Ueno, M.; Kim, D.; Oda, T. Antioxidant and anti-inflammatory activities of porphyran isolated from discolored nori (Porphyra yezoensis). Int. J. Biol. Macromol. 2015, 74, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Ha, Y.; Lee, W.H.; Jeong, J.; Park, M.; Ko, J.Y.; Kwon, O.W.; Lee, J.; Kim, Y.J. Pyropia yezoensis extract suppresses IFN-Gamma- and TNF-Alpha-induced proinflammatory chemokine production in HaCaT cells via the down-regulation of NF-κB. Nutrients 2020, 12, 1238. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, R.C.L.; da Silva, M.S.J.; de Alencar, D.B.; de Pires, F.A.; de Alencar, N.M.; Pereira, M.G.; Cavada, B.S.; Sampaio, A.H.; Farias, W.R.L.; Assreuy, A.M.S. In vivo anti-inflammatory effect of a sulfated polysaccharide isolated from the marine brown algae Lobophora variegate. Pharm. Biol. 2011, 49, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agyemang, K.; Han, L.; Liu, E.; Zhang, Y.; Wang, T.; Gao, X. Recent advances in Astragalus membranaceus anti-diabetic research: Pharmacological effects of its phytochemical constituents. Evid. Based Complement. Altern. Med. 2013, 654643, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Volpi, N. Anti-inflammatory activity of chondroitin sulphate: New functions from an old natural macromolecule. Inflammopharmacology 2011, 19, 299–306. [Google Scholar] [CrossRef]
- Wijesekara, I.; Pangestuti, R.; Kim, S.K. Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohydr. Polym. 2011, 84, 14–21. [Google Scholar] [CrossRef]
- Lee, M.H.; Lee, K.B.; Oh, S.M. Antifungal activities of dieckol isolated from the marine brown alga Ecklonia cava against Trichophyton rubrum. J Korean Soc. Appl. Biol. Chem. 2010, 53, 504–507. [Google Scholar] [CrossRef]
- Ertürk, O.; Taş, B. Antibacterial and antifungal effects of some marine algae. Kafkas Univ. Vet. Fak. Derg. 2011, 17, 121–124. [Google Scholar]
- Manivannan, K.; Karthikai, D.G.; Anantharaman, P.; Balasubramanian, T. Antimicrobial potential of selected brown seaweeds from Vedalai coastal waters, Gulf of Mannar. Asian Pac. J. Trop. Biomed. 2011, 1, 114–120. [Google Scholar] [CrossRef] [Green Version]
- Mickymaray, S.; Alturaiki, W. Antifungal efficacy of marine macroalgae against fungal isolates from bronchial asthmatic cases. Molecules 2018, 23, 3032. [Google Scholar] [CrossRef] [Green Version]
- Hamed, S.M.; Abd El-Rhman, A.A.; Abdel-Raouf, N.; Ibraheem, I.B.M. Role of marine macroalgae in plant protection & improvement for sustainable agriculture technology review. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 104–110. [Google Scholar]
- Shobier, A.H.; Abdel Ghani, S.A.; Barakat, K.M. GC/MS spectroscopic approach and antifungal potential of bioactive extracts produced by marine macroalgae. Egypt J. Aqua. Res. 2016, 42, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Oh, K.; Lee, J.H.; Chung, S.C.; Shin, J.; Shin, H.J.; Kimd, H.K.; Lee, H.S. Antimicrobial activities of the bromophenols from the red alga Odonthalia corymbifera and some synthetic derivatives. Bioorg. Med. Chem. Lett. 2008, 18, 104–108. [Google Scholar] [CrossRef]
- Wang, W.; Wang, S.X.; Guan, H.-S. The antiviral activities and mechanisms of marine polysaccharides: An overview. Mar. Drugs 2012, 10, 2795–2816. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, A.; Zorofchian, M.S.; Zandi, A.Z. Antiviral potential of algae polysaccharides isolated from marine sources: A review. Biomed. Res. Int. 2015, 825203. [Google Scholar] [CrossRef] [Green Version]
- Elizondo-Gonzalez, R.; Cruz-Suarez, E.; Ricque-Marie, D.; Mendoza-Gamboa, E.; Rodriguez-Padilla, C.; Trejo-Avila, L.M. In vitro characterization of the antiviral activity of fucoidan from Cladosiphon okamuranus against Newcastle Disease Virus. Virol. J. 2012, 9, 307. [Google Scholar] [CrossRef] [Green Version]
- Pujol, C.A.; Ray, S.; Ray, B.; Damonte, E.B. Antiviral activity against dengue virus of diverse classes of algal sulfated polysaccharides. Int. J. Biol. Macromol. 2012, 51, 412–416. [Google Scholar] [CrossRef]
- Krylova, N.V.; Ermakova, S.P.; Lavrov, V.F.; Leneva, I.A.; Kompanets, G.G.; Iunikhina, O.V.; Nosik, M.N.; Ebralidze, L.K.; Falynskova, I.N.; Silchenko, A.S.; et al. The comparative analysis of antiviral activity of native and modified fucoidans from brown algae Fucus evanescens in vitro and in vivo. Mar. Drugs 2020, 18, 224. [Google Scholar] [CrossRef] [Green Version]
- Eom, S.H.; Moon, S.Y.; Lee, D.S.; Kim, H.J.; Park, K.; Lee, E.W.; Kim, Y.M. In vitro antiviral activity of dieckol and phlorofucofuroeckol-A isolated from edible brown alga Eisenia bicyclis against murine norovirus. Algae 2015, 30, 241–246. [Google Scholar] [CrossRef] [Green Version]
- Serkedjieva, J. Antiviral activity of the red marine alga Ceramium rubrum. Phytother. Res. 2004, 18, 480–483. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Laminaria digitata | Dark brown, to 2 m in length; presents a claw-like holdfast, a smooth, flexible stipe, and also a laminate blade to 1.5 m long split into finger-like segments [80] |
| The species is rich in alginates, mannitol, and amino acids. [20,81] | |
| Very rich in iodine; therefore, this seaweed promotes slimming and has antiseptic properties [82] | |
| Dictyota dichotoma | Thallus is flat and leaf-like, up to 30 mm long and 5–30 mm broad. The fronds are thin and translucent; colors differ from olive to golden-brown [80] |
| Produces large amounts of heterogeneous polysaccharides when submitted to the extraction procedures used to obtain fucoidans [83] | |
| Undaria pinnatifida | Thallus fixed by a ramified holdfast [84] |
| Rich source of eicosapentaenoic acid, an omega-3 fatty acid, and presents high levels of sodium, calcium, iodine, thiamine, and niacin [16,38] | |
| Enteromorpha linza | Can be distinguished by its smooth thallus, most with a ruffled margin [85] |
| Rich in essential amino acids, fatty acids, vitamins, dietary fiber, and resistant protein [2,3,86] | |
| Ecklonia cava | A perennial brown alga and exists mainly in subtidal areas off the coast of Japan and Korea [87] |
| Eckol isolated from E. cava attenuates oxidative stress-induced cell damage in lung fibroblast cells [88] | |
| Exhibits radical scavenging activity, but also antiplasmin inhibiting activity, antimutagenic activity, bactericidal activity, human immunodeficiency virus type 1 (HIV-1) reverse transcriptase, and protease inhibition [89] |
| Algae Species | Active Extract/Compound | Biological Activity | Reference |
|---|---|---|---|
| Phorphyra dentate | Methanolic extracts | Anti-inflammatory effect in lipopolysaccharide (LPS) induced mouse RAW 264.7 macrophages cell line. | [152] |
| Caulerpa mexicana | Methanolic extracts | Decrease the xylene-induced ear edema and reduce cell migration to different sites. | [153] |
| Myagropsis myagroides | Fucoxanthin | LPS-stimulated RAW 264.7 macrophages. | [154] |
| Ulva reticulate | Methanolic extracts | Carrageenan-induced hind paw edema in rats and peritonitis in acute and chronic inflammatory models. | [155] |
| Laminaria saccharina | Sulfated polysaccharides | Inhibits leukocyte recruitment in rat and the neutrophil adhesion to platelets. | [156] |
| Porphyra dioica Palmaria palmate Chondrus crispus | β-carotene fucoxanthin PUFA | Able to inhibit LPS-induced inflammatory pathways in human macrophages. | [157] |
| Caulerpa cupressoides | Sulfated polysaccharides | Decrease neutrophils migration. Strongly reduced the carrageenan-induced rat paw edema. | [158] |
| Dictyota menstrualis | Heterofucan | Binds to the surface of leucocytes and inhibits migration of leucocytes to the site of injury. Inhibit the chemical-induced leukocyte migration into the peritoneal cavity. | [159] |
| Dictyopteris prolifera Grateloupia lanceolata Grateloupia filicina | Ethanolic extract | Concentration-dependent reduction of LPS-induced prostaglandin E2 production. Suppresses the expression of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) at the protein level in RAW 264.7 cells. Reduced the release of tumor necrosis factor alpha (TNF-α) and interleukin 6 (IL-6) into the medium. | [160] |
| Gracilaria cornea | Sulfated polysaccharide fraction | Significantly inhibits rat paw edema induced by different inflammatory agents (carrageenan and dextran, histamine and L-arginin). Downregulates interleukin-1β (IL-1β), TNF-α, and COX-2 mRNA and protein levels. | [161] |
| Porphyra yezoensis | Dc-porphyran | Inhibits nitric oxide (NO) production in LPS-stimulated RAW 264.7 cells. | [162] |
| Pyropia yezoensis | Astaxanthin Xanthophyl | Anti-inflammatory action. | [163] |
| Lobophora variegata | Fucans | Inhibits the paw edema, plasma exudation, nitrite content, and leukocyte migration. | [164] |
| Red algae | Carrageenan, Fucoidan, Chondroitin | Lowered the expression of inducible nitric oxide synthase (iNOS). Inhibited the expressions of TNF-α, IL-1β, and interferon-c (IFN-c). Repressed pro-inflammatory cytokines and suppressed the activity of COX-2. | [41,165,166,167] |
| Fucus vesiculosus | Fucoidan | Anti-atopic dermatitis | [103,107] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biris-Dorhoi, E.-S.; Michiu, D.; Pop, C.R.; Rotar, A.M.; Tofana, M.; Pop, O.L.; Socaci, S.A.; Farcas, A.C. Macroalgae—A Sustainable Source of Chemical Compounds with Biological Activities. Nutrients 2020, 12, 3085. https://doi.org/10.3390/nu12103085
Biris-Dorhoi E-S, Michiu D, Pop CR, Rotar AM, Tofana M, Pop OL, Socaci SA, Farcas AC. Macroalgae—A Sustainable Source of Chemical Compounds with Biological Activities. Nutrients. 2020; 12(10):3085. https://doi.org/10.3390/nu12103085
Chicago/Turabian StyleBiris-Dorhoi, Elena-Suzana, Delia Michiu, Carmen R. Pop, Ancuta M. Rotar, Maria Tofana, Oana L. Pop, Sonia A. Socaci, and Anca C. Farcas. 2020. "Macroalgae—A Sustainable Source of Chemical Compounds with Biological Activities" Nutrients 12, no. 10: 3085. https://doi.org/10.3390/nu12103085
APA StyleBiris-Dorhoi, E. -S., Michiu, D., Pop, C. R., Rotar, A. M., Tofana, M., Pop, O. L., Socaci, S. A., & Farcas, A. C. (2020). Macroalgae—A Sustainable Source of Chemical Compounds with Biological Activities. Nutrients, 12(10), 3085. https://doi.org/10.3390/nu12103085